
淹水胁迫及胁迫解除对甜瓜光合色素和光合特性的影响
2013-05-21 02:13:34王俊良龙启炎汪李平骆海波吕慧芳徐翠容
中国蔬菜 2013年8期
王俊良龙启炎汪李平骆海波吕慧芳徐翠容
(1武汉市蔬菜科学研究所,湖北 武汉 430065;2 华中农业大学园艺林学学院,湖北 武汉 430070;3安徽池州学院,安徽 池州 247000)
甜瓜(Cucumis melo L.)性喜温热,对湿涝环境敏感。我国长江中下游地区春季阴雨连绵,夏季经常出现暴雨,甜瓜幼苗生长受到抑制或死亡,导致产量下降,严重影响露地甜瓜的生产,但关于淹水胁迫对甜瓜光合色素和光合特性的影响却鲜有报道。本试验研究了淹水胁迫及胁迫解除后两个耐涝性差异显著的甜瓜品系的光合色素和光合特性的变化,以期为甜瓜的耐涝性研究奠定基础,为抗逆育种和生产提供服务。
1 材料与方法
1.1 试验材料
供试材料为甜瓜品系T106和T241,经耐涝性鉴定,二者对淹水胁迫的耐受性差异显著,其中T106为耐涝品系,T241为不耐涝品系。
1.2 试验设计
试验于2012年6月中旬在华中农业大学塑料大棚中进行,分别挑选两个品系籽粒饱满的甜瓜种子,55 ℃浸种5 h,然后在30 ℃恒温培养箱中催芽,播于72孔育苗盘中,待2片子叶完全展开后,选取长势一致的幼苗移栽至直径10cm的营养钵中,培养土为经过消毒的菜园土。当甜瓜幼苗长至四叶一心时,采用双套盆法(将营养钵放入120cm×60cm×40cm的塑料盆中)进行淹水处理,每个塑料盆中放10个营养钵,在塑料盆中注入经过晾晒的自来水,使水面高于营养钵土面3~4cm,每天加水以保持水位一致,在淹水7d后进行排水处理。采用随机区组设计,设淹水和不淹水(CK)2个处理,每处理3次重复,每重复100株,淹水处理前按常规方法管理。处理期间的平均温度为白天33 ℃,夜间26 ℃,用浙江大学生产的ZDR-21型温湿度记录仪每3 h记录1次。
1.3 测定项目
分别在进行淹水处理的0、1、3、5、7d和排水后的1、3、5、7d进行光合色素和光合特性的测定。
1.3.1 光合参数的测定 采用随机取样的方法,每处理选取60株,选择植株顶部向下第2、3片叶序的功能叶,用TPS-1光合测定仪(英国PPSystems国际有限公司生产)在晴天的9:00~11:00进行测定。采用自然光照射,测定温度为32 ℃左右。
1.3.2 光合色素含量的测定 参照李合生(2000)的分光光度计法进行测定。将采回的叶片样品洗净擦干,去除边缘及中脉,称取约0.2 g,剪碎放入三角瓶内,加入25mL混合液(乙醇∶丙酮∶蒸馏水=4.5 V∶4.5 V∶1.0 V),保鲜膜封口,置于暗处过夜浸提,待材料完全变白后,用721型分光光度计分别测定665、649、470 nm波长下的光密度值,根据经验公式分别计算出叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量。每处理3次重复。
1.4 数据分析
采用Excel 2003软件进行数据处理和作图,采用SAS 8.1软件的ANOVA过程进行方差分析。
2 结果与分析
2.1 淹水胁迫及胁迫解除对甜瓜幼苗叶片光合色素含量的影响
由图1可见,淹水胁迫前2个甜瓜品系幼苗叶片的叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Car)、总叶绿素〔Chl (a+b)〕含量之间差异均不显著;在淹水胁迫条件下,2个甜瓜品系的Chla、Chlb、Car、Chl (a+b)含量的变化趋势相似,均随着淹水胁迫时间的延长,光合色素含量出现不同程度的下降;不耐涝品系T241光合色素含量的下降幅度大于耐涝品系T106。在淹水胁迫第7天时,T106的Chla、Chlb、Car和Chl(a+b)含量均显著高于T241;T106的Chla、Chlb、Car和Chl (a+b)含量分别比淹水前下降了28.54%、9.45%、14.84%、23.77%,而T241则分别比淹水前下降了39.34%、22.41%、32.14%、34.87%。在淹水胁迫解除后,T106的 Chla、Chlb、Car、Chl(a+b)含量的上升幅度均大于T241,光合色素含量恢复较快。在淹水胁迫解除第7天,T106的Chla、Chlb、Car、Chl(a+b)含量均显著高于T241;与淹水胁迫刚解除时相比,T106的Chla、Chlb、Car、Chl(a+b) 含 量 分 别 增 加 了 13.21%、11.94%、12.25%、12.46%,比淹水前下降了18.24%、4.28%、6.47%、12.88%, 而 T241则分别比淹水胁迫刚解除时增加了4.67%、7.27%、9.66%、3.04%,比淹水前下降了36.14%、19.57%、23.38%、31.79%。说明淹水胁迫抑制了甜瓜幼苗叶片光合色素的合成并加速了光合色素的分解,且对T241的光合色素损害程度要大于T106;在淹水胁迫解除后,耐涝品系T106的光合色素的增加量大于不耐涝品系T241。
2.2 淹水胁迫及胁迫解除对甜瓜幼苗叶片光合特性的影响
2.2.1 淹水胁迫及胁迫解除对甜瓜幼苗叶片净光合速率的影响 由图2可见,淹水胁迫前2个甜瓜品系幼苗叶片的净光合速率之间差异不显著;在整个淹水胁迫期间,2个甜瓜品系的净光合速率均呈下降趋势,不耐涝品系T241的净光合速率的下降幅度大于耐涝品系T106。在淹水胁迫第7天,T106的净光合速率显著高于T241;T106的净光合速率比淹水前下降了26.31%,而T241比淹水前下降了39.67%。在淹水胁迫解除后,T106的净光合速率上升幅度大于T241。在淹水胁迫解除第7天,T106的净光合速率显著高于T241;T106的净光合速率比刚解除淹水胁迫时增加了20.19%,比淹水前下降了13.64%;而T241则比刚解除淹水胁迫时增加了9.63%,比淹水前下降了31.44%。2个甜瓜品系的净光合速率在淹水胁迫解除7d后均未恢复到淹水前的水平,说明二者的净光合速率均受到了淹水胁迫的抑制,耐涝品系T106的受抑制程度轻于不耐涝品系T241,在淹水胁迫解除后T106的恢复能力也强于T241。
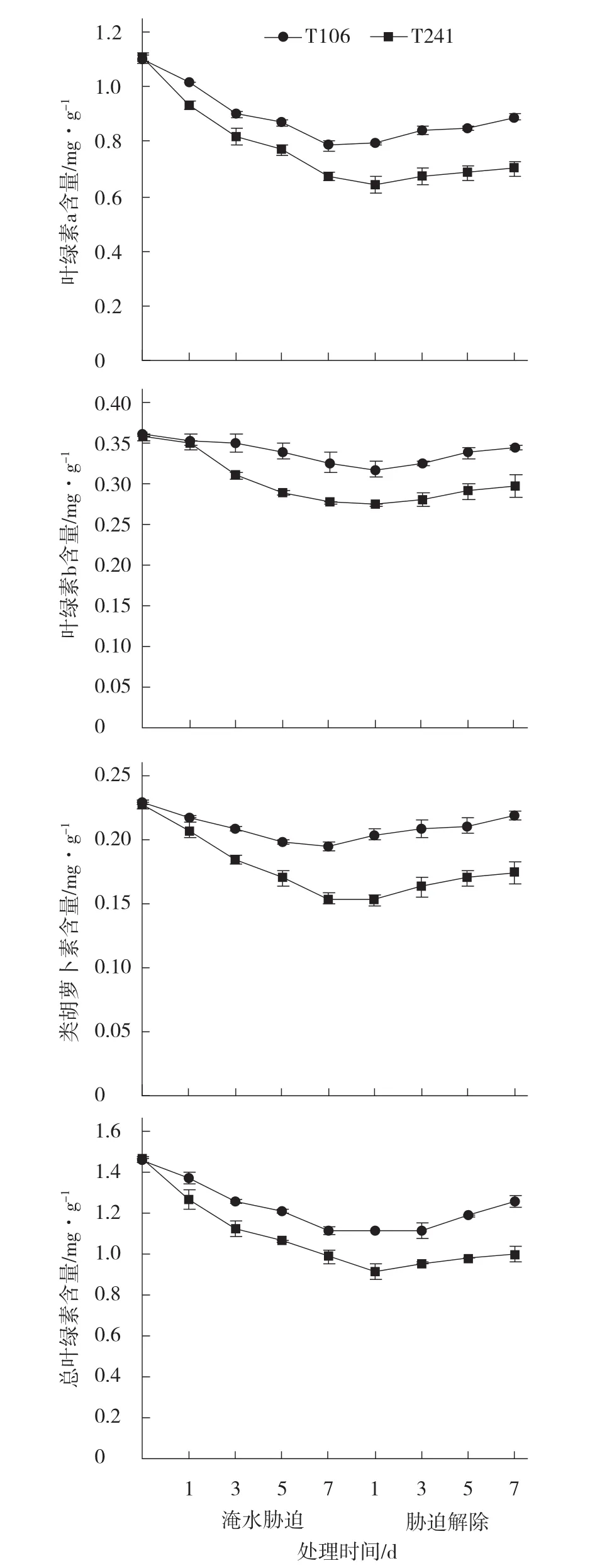
图1 淹水胁迫及胁迫解除对甜瓜幼苗叶片光合色素含量的影响
2.2.2 淹水胁迫及胁迫解除对甜瓜幼苗叶片蒸腾速率的影响 由图3可见,淹水胁迫前2个甜瓜品系幼苗叶片的蒸腾速率之间差异不显著;淹水胁迫下2个甜瓜品系的蒸腾速率均呈下降趋势。在淹水胁迫第7天时,T106的蒸腾速率显著高于T241;T106的蒸腾速率比淹水前下降了28.12%,而T241比淹水前下降了41.78%。在淹水胁迫解除后第1天,T106的蒸腾速率开始增加,而T241则在淹水胁迫解除后第3天开始增加,且T106蒸腾速率的上升幅度大于T241。在淹水胁迫解除第7天,T106的蒸腾速率显著高于T241,T106的蒸腾速率比淹水胁迫刚解除时增加了25.17%,比淹水前下降了10.36%;而T241比淹水胁迫刚解除时增加了9.54%,比淹水前下降了36.64%。说明淹水胁迫明显抑制了甜瓜的蒸腾速率,对不耐涝品系T241的抑制程度大于耐涝品系T106。
2.2.3 淹水胁迫及胁迫解除对甜瓜幼苗叶片气孔导度的影响 由图4可见,淹水胁迫前2个甜瓜品系幼苗叶片的气孔导度之间差异不显著;淹水胁迫下,2个甜瓜品系的气孔导度均呈下降趋势。在淹水胁迫第7天时,耐涝品系T106的气孔导度显著高于不耐涝品系T241;T106的气孔导度比淹水前下降了17.71%,而T241比淹水前下降了32.94%,T106的下降幅度小于T241。在淹水胁迫解除后,2个甜瓜品系的气孔导度都逐渐上升,T106的上升幅度大于T241。在淹水胁迫解除第7天,T106的气孔导度显著高于T241;T106的气孔导度比淹水胁迫刚解除时增加了12.64%,比淹水前下降了7.15%,而T241比淹水胁迫刚解除时增加了8.6%,比淹水前下降了27.12%。说明淹水胁迫导致甜瓜幼苗叶片气孔收缩或部分关闭,淹水胁迫对耐涝品系气孔导度的抑制程度低于不耐涝品系。
2.2.4 淹水胁迫及胁迫解除对甜瓜幼苗叶片胞间CO2浓度的影响 由图5可见,淹水胁迫前2个甜瓜品系幼苗叶片的胞间CO2浓度差异不显著。随着淹水胁迫时间的延长,2个甜瓜品系的胞间CO2浓度均呈逐渐下降趋势,不耐涝品系T241的下降幅度大于耐涝品系T106。在淹水胁迫第7天时,T106的胞间CO2浓度显著高于T241;T106的胞间CO2浓度比淹水前下降了18.74%,而T241比淹水前下降了31.59%。淹水胁迫解除后,T106和T241的胞间CO2浓度都有不同程度的增加,T106的增加幅度大于T241。在淹水胁迫解除第7 天时,T106的胞间CO2浓度显著高于T241;T106的胞间CO2浓度比淹水胁迫刚解除时增加了8.14%,比淹水前下降了12.14%;而T241则比淹水胁迫刚解除时增加了5.18%,比淹水前下降了30.81%。说明淹水胁迫使甜瓜幼苗叶片胞间CO2浓度降低,不耐涝品系的下降幅度大于耐涝品系。
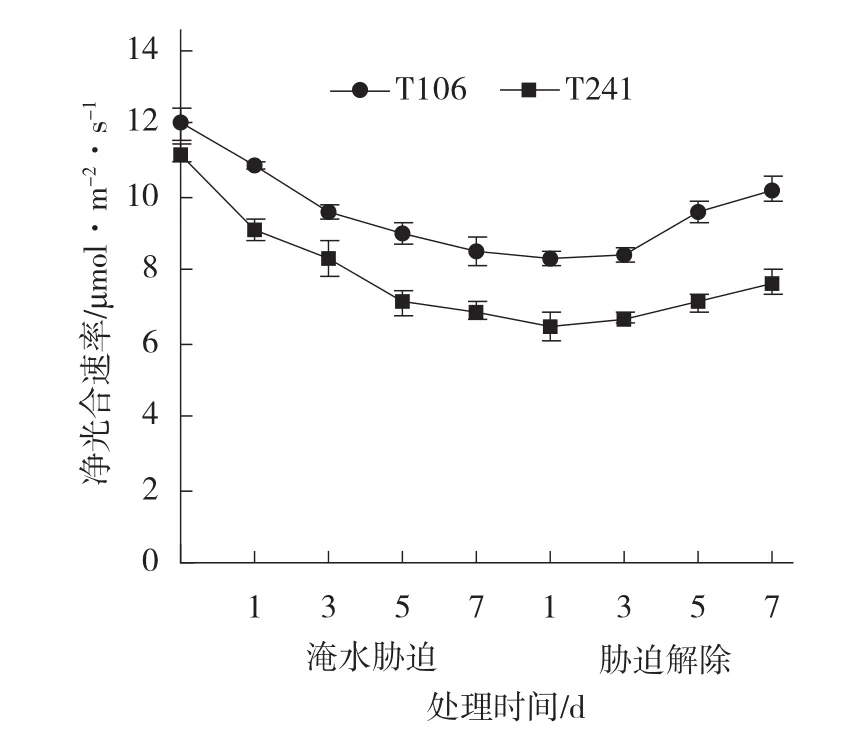
图2 淹水胁迫及胁迫解除对甜瓜幼苗叶片净光合速率的影响

图3 淹水胁迫及胁迫解除对甜瓜幼苗叶片蒸腾速率的影响

图4 淹水胁迫及胁迫解除对甜瓜幼苗叶片气孔导度的影响
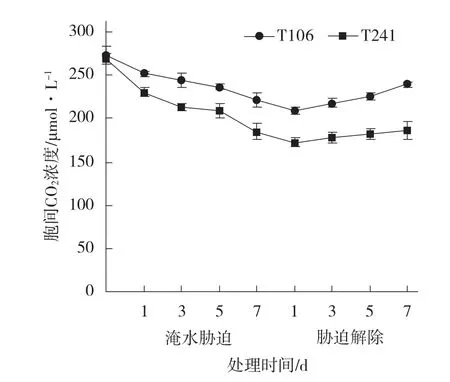
图5 淹水胁迫及胁迫解除对甜瓜幼苗叶片胞间CO2浓度的影响
3 结论与讨论
3.1 淹水胁迫及胁迫解除对甜瓜光合色素含量的影响
叶绿体中有很多光合作用所必须的光合色素。植物对光能的吸收和转化也离不开叶绿素和类胡萝卜素。在正常的外界环境条件下,植物体内的叶绿体色素处于稳定状态,但在各种各样的逆境胁迫下光合色素呈不稳定状态。张克清(2007)的研究表明,在淹水环境条件下植株叶片的叶绿素含量呈逐渐下降趋势。在淹水环境条件下,植株主要进行无氧呼吸,导致根系的能量供应不足,不能有效地吸收矿质元素,使叶片的生长发育受到抑制,叶绿素合成受到抑制,加速了叶绿素的降解过程。宫长荣和汪耀富(1995)对烤烟的耐涝性研究发现,淹水胁迫过程中叶绿素含量的降低与叶片分解代谢加强、合成代谢受阻有关。本试验结果表明:随着淹水胁迫时间的延长,2个甜瓜品系幼苗叶片的叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量均呈下降趋势,不耐涝品系T241的下降幅度大于耐涝品系T106;在淹水胁迫解除后,随着胁迫解除时间的延长,2个甜瓜品系的光合色素含量都得到不同程度的恢复,耐涝品系T106光合色素含量的增加幅度大于不耐涝品系T241。这与王萍等(2007)在牡丹上的研究结果相似。
3.2 淹水胁迫及胁迫解除对甜瓜光合特性的影响
张晓平(2004)的研究表明,在淹水逆境胁迫下植物的光合作用受到显著抑制,同化产物显著减少。在淹水胁迫下植株叶片的气孔会出现不同程度的关闭,导致受淹植物光合作用受到显著抑制,光合作用下降的后期继而引发了叶片的失绿、发黄、逐渐衰老并出现脱落现象。Pezeshki等(1998)的研究发现,落羽杉和洋白蜡之所以耐涝性强是因为在淹水胁迫中其叶片的部分气孔重新开放且能维持较高的气孔导度。王生(1998)在对苗木进行淹水胁迫时发现,淹水处理20d后柳杉的气孔导度呈逐渐增大趋势,而两种杨树则呈逐渐下降趋势。对耐涝性弱的植物来说,在淹水胁迫下植株叶片的气孔导度与其光合速率、蒸腾速率呈显著相关关系。潘向艳(2006)的研究表明,随着淹水胁迫时间的延长,植物的气孔导度和蒸腾速率会显著下降。张晓平(2004)的研究表明,各种对植物生长不利的逆境条件都显著抑制植物的光合作用,主要有两个方面的原因,一是气孔因素,即气孔逐渐闭合,胞间CO2浓度逐渐下降,使叶绿体内CO2的量呈下降趋势;二是非气孔因素,在淹水胁迫下细胞内的CO2溶解度逐渐下降、RuBP羧化酶对CO2的亲和力降低或光合机构关键成分的稳定性降低等。梁哲军等(2009)的研究表明,玉米在淹水胁迫解除后净光合速率、气孔导度、胞间CO2浓度都得到了一定的恢复,耐涝品种的恢复能力要好于不耐涝品种。
本试验中,随着淹水胁迫时间的延长,2个甜瓜品系幼苗叶片的净光合速率、气孔导度、蒸腾速率均呈下降趋势,这与汤玉喜等(2008)的研究结果一致。张晓平(2004)在鹅掌楸上的研究表明,随着淹水时间的延长鹅掌楸的净光合速率、气孔导度、蒸腾速率都呈下降趋势,而胞间CO2浓度则逐渐上升。本试验中,随着淹水胁迫时间的延长,2个甜瓜品系幼苗叶片的胞间CO2浓度逐渐降低,这与前人的结果不一致,原因可能是气孔闭合导致细胞间的CO2浓度逐渐下降。在淹水胁迫期间,2个甜瓜品系幼苗叶片的净光合速率受到了明显的抑制,这与气孔导度下降有关,即气孔的闭合降低了胞间CO2浓度,使叶绿体内CO2的供应受阻;也可能是淹水后期光合色素含量下降所致。这表明在淹水后期(淹水4~7d)甜瓜光合作用受到抑制是来自于气孔限制和非气孔限制的双重作用。随着淹水胁迫解除时间的延长,2个甜瓜品系的净光合速率、气孔导度、蒸腾速率、胞间CO2浓度均得到恢复性增加,T106的增加幅度大于T241,这说明耐涝品系T106在淹水胁迫下光合作用受到的抑制程度低于不耐涝品系T241。这与梁哲军等(2009)在玉米上的研究结果相似。
宫长荣,汪耀富.1995.淹水胁迫对烤烟生理生化特性的影响.中国农业科学,28(s):126-130.
李合生.2000.植物生理生化实验原理和技术.北京:高等教育出版社.
梁哲军,陶洪斌,王璞.2009.淹水解除后玉米幼苗形态及光合生理特征恢复.生态学报,29(7):3978-3983.
潘向艳.2006.杂交鹅掌楸不同无性系对淹水胁迫的反应〔硕士论文〕.南京:南京林业大学.
汤玉喜,周金星,吴敏.2008.淹水胁迫下美洲黑杨新无性系光合特征的变化.林业科技开发,22(6):20-24.
王萍,胡永红,王丽勉.2007.观赏植物耐涝性鉴定指标的种类及其评价方法.北方园艺,(11):78-81.
王生.1998.淹水胁迫对杨树无性系苗期生长及生理的影响.云南林业科技,(2):28-33.
张克清.2007.淹水胁迫对5种豆科植物生长及生理性状的影响〔硕士论文〕.南京:南京农业大学.
张晓平.2004.不同种源鹅掌楸和杂种鹅掌楸对淹水胁迫的响应〔博士论文〕.南京:南京林业大学.
Pezeshki S R,Pardue J H,Delaune R D.1996.Leaf gas exchange and growth of flood-tolerant and flood-sensitive tree species under low soilredox conditions.Tree Physiology,16(4):453-458.
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
湖北农业科学(2021年13期)2021-08-04 03:56:18
新疆农业科技(2021年2期)2021-07-03 06:39:34
节水灌溉(2021年5期)2021-05-26 06:49:38
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
中学生物学(2020年12期)2020-03-30 07:27:46
中国农业科技导报(2020年12期)2020-03-15 10:50:08
生物技术通报(2014年10期)2014-03-21 06:56:38
