
花生果皮全长cDNA文库的构建及初步分析
2013-04-25 03:21姜宝杰蔡铁城曾建斌庄伟建
福建农林大学学报(自然科学版) 2013年1期
陈 华,姜宝杰,张 冲,蔡铁城,曾建斌,邓 烨,庄伟建
(福建省作物分子与细胞生物学重点实验室,福建福州350002)
花生果皮由子房壁发育而来,由外果皮、中果皮、内果皮组成.外果皮含表皮和周皮层;中果皮含薄壁细胞、纤维层和维管束;内果皮由内薄壁细胞和内表皮组成.在发育成熟的过程中,中果皮纤维层日益木质化,果壳逐渐变薄、变硬[1].花生在其生长发育以及储存过程当中极易受到黄曲霉等各种病原菌的侵染,严重影响花生生产、加工和贸易[2].花生果皮是抵御病原菌如黄曲霉菌、禾谷丝核菌侵染及地下害虫如蛴螬破坏的天然屏障,提高花生果皮对各种病虫害的抗性可有效保护花生的食用部位——种仁.花生果壳褐斑病、烂果病、黄曲霉病及地老虎、蛴螬等病虫危害造成花生产量及品质严重下降.然而,目前对于花生抗病虫害分子生物学研究较少且主要集中在花生种皮[3].研究花生果皮功能基因组,对花生果皮抗病虫遗传改良研究提供基因源,尚未有涉足,远远落后于水稻[4-5]等其他作物.
本试验以闽花6号花生不同发育时期的果皮为材料,利用Clontech公司的SMART技术并加以改进构建了花生果皮全长cDNA文库,并随机测序了30个文库单克隆,对测序成功的序列进行生物信息学分析,这将为花生果皮的研究及利用提供材料平台.
1 材料与方法
1.1 材料
1.1.1 果皮材料 花生为福建农林大学油料作物研究所选育的优良花生品种闽花6号,取果针入土后10、20、30、40、50、60 d 6 个时期的果皮为材料,置 -70 ℃冰箱中保存备用.
1.1.2 主要试剂 文库构建试剂盒CreatorTMSMARTTMcDNA Library Construction Kit购自Clontech公司.SuperscriptⅢ Reverse Transcriptase为Invitrogen 公司产品.Ex Taq,dNTP,DL2000、DL15000 分子标准,电转化感受态细胞DH5α均购自大连宝生物公司.胶回收试剂盒购自杭州博日科技有限公司.其他试剂采用国产分析纯.
1.2 方法
1.2.1 花生果皮总RNA的提取 分别取花生果皮各时期的材料等量混合,采用CTAB法提取总RNA:在液氮预冷的研钵中将材料研磨成粉状,转移至65℃含预热CTAB的离心管中,水浴20-30 min,氯仿/异戊醇抽提2次后,上层水相加入LiCl沉淀总RNA.于4℃ 12000 g离心30 min,将沉淀物溶于适量DEPC处理水后用DnaseⅠ处理除去DNA,用ND-2000型分光光度计(美国Thermo公司)检测总RNA的浓度和纯度,琼脂糖凝胶电泳检测其完整性.
1.2.2 cDNA双链的合成与SfiⅠ限制性酶切消化 以3 μg总RNA为模板,按照CreatorTMSMARTTMcDNA Library Construction Kit说明合成cDNA第一链.在PCR管中加入单链模板、10×Ex Taq Buffer、10 mmol·L-1dNTP、5'PCR Primer、Ex Taq,反应在已预热到95℃的PCR仪上进行.反应条件:95℃预变性2 min;95℃变性15 s,68℃复性30 s,72℃延伸5 min,共15个循环;72℃延伸10 min.取2 μL反应产物进行1.0%琼脂糖凝胶电泳鉴定.
50 μL双链cDNA的PCR产物经蛋白酶K灭活DNA聚合酶活性、氯仿/异戊醇抽提、乙醇室温沉淀后重悬于79 μL去离子水中.加入15 μL SfiⅠ酶于50℃酶切2 h,再以1.5%琼脂糖凝胶电泳酶切后的双链,胶回收750 bp以上的片段.回收产物经乙醇、醋酸钠、糖原于-20℃共沉淀后,以7 μL去离子水溶解,保存于-70℃备用.
1.2.3 连接与转化 胶回收产物在T4 DNA连接酶的作用下,与质粒载体 pDNR-LIB(经SfiⅠ酶处理过)在16℃下连接16 h.连接产物经无水乙醇、糖原共沉淀后溶于5 μL双蒸水中.大肠杆菌感受态(45 μL)从-70℃冰箱中取出,冰上解冻后将连接产物加入到感受态中电击转化.迅速用950 μL不含抗生素的LB培养基洗出转化产物,于37℃、225 r·min-1摇菌复苏1 h,得到的菌液即为原始文库.
1.2.4 文库的鉴定 于37℃摇菌1h后,吸取1 μL菌液涂含30 mg·L-1氯霉素的LB培养板,过夜培养,计数单克隆数以计算原始文库量,计算公式为:原始文库滴度=生长单克隆数×103.随机挑取22个单菌落用M13引物(5'-GTAAAACGACGGCCAG-3'、5'-CAGGAAACAGCTATGAC-3')进行菌液PCR鉴定文库的阳性率;随机挑选10个单菌落提取质粒,SfiⅠ酶切后电泳检测插入片段的大小;随机挑取30个单菌落做穿刺培养后送上海国家人类基因组南方研究中心进行测序,将所得序列去除载体和引物序列后分别进行BlastN和BlastX对比和Blast2GO功能注释.
1.2.5 文库的扩增 其余原始文库菌液涂含30 mg·L-1氯霉素的LB培养板,过夜培养,用液体LB培养基洗脱培养板即为扩增文库.洗脱后的菌液用终浓度为25%的甘油分装保存于-80℃的冰箱中.
2 结果与分析
2.1 总RNA的纯度及完整性检测
花生果皮各时期等量混合提取的总RNA经紫外分光光度计检测,D260nm/D280nm为1.98.1.0%琼脂糖凝胶电泳(图1)显示,28S和18S rRNA条带清晰,亮度比接近2∶1,表明获得的RNA纯度较高,没有基因组DNA污染,并且总RNA没有降解,比较完整,可用做逆转录模板合成cDNA第一链.
2.2 ds cDNA的合成及鉴定
LD-PCR法合成的ds cDNA经1.0%琼脂糖凝胶电泳检测,呈现从小到大均匀分布的弥散条带,ds cDNA片段条带大小为200-3000 bp(图2),主要集中在1000 bp以上,基本反映了花生果皮的基因信息.
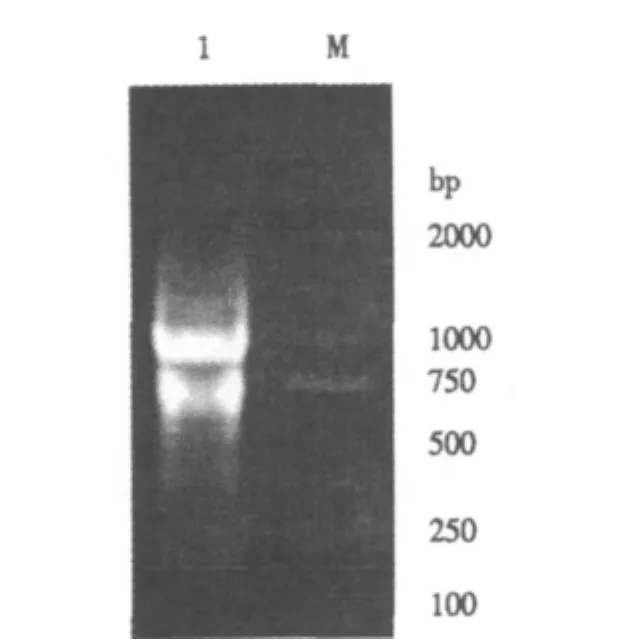
图1 花生果皮总RNA电泳图Fig.1 Gel electrophoresis of total RNA isolated from peanut pericarp
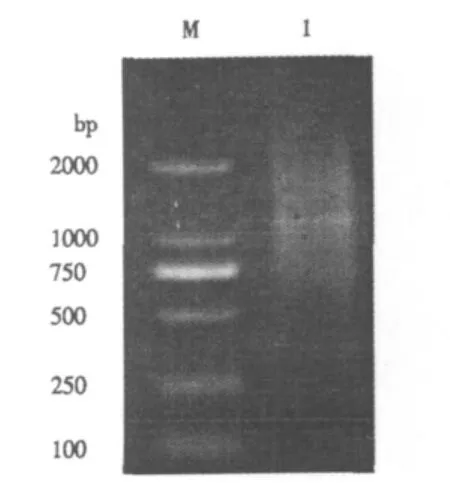
图2 花生果皮ds cDNA电泳图Fig.2 Gel electrophoresis of peanut pericarp ds cDNA
2.3 文库质量的鉴定
双链经酶切、连接载体、电击转化、涂板过夜培养后计数培养板单克隆数,原始文库滴度达1.3×107cfu.随机挑取22个单菌落摇菌,进行菌液PCR扩增,电泳显示仅有1个为空载体(图3),重组率达95.5%.提取质粒酶切后电泳显示,插入片段大小为750-2000 bp,平均长度约1000 bp(图4).重组率及插入片段大小均达到全长cDNA文库的质量要求.
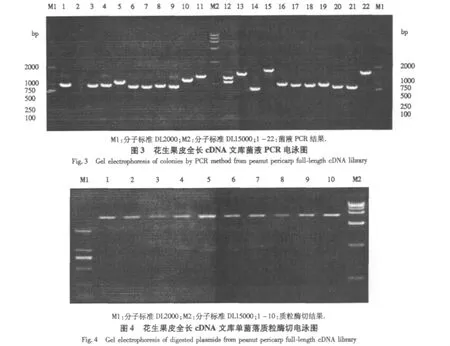
2.4 部分克隆的生物信息
随机挑选文库30个单克隆进行双向测序,利用DNA序列常规分析软件BioXM 2.6分析测序结果,共获得25条有效序列.经Bioedit软件进行序列拼接后,去除引物及载体序列,25条序列3'端均有polyA加尾信号,17条序列5'端具有完整的帽子结构,因此判断所得序列中有17条为全长序列,所占比例为68%.
BlastX分析结果显示,所测序的25条序列与已报道的氨基酸序列均有较高的同源性,有13条序列与其他植物已知功能氨基酸序列具有较高的相似性,包括热休克蛋白序列、钙调蛋白序列、植物血凝素类序列、激酶类序列、预测的抗病相关序列等.未知功能序列有12条(48%).所得到的25条序列没有重复,说明该文库的代表性比较高.BlastX比对结果中有10条序列与大豆序列具有同源性,仅有3个基因与已报道的花生序列具有同源性(表1).这主要是由于大豆与花生同属豆科植物,同源性较高.而花生功能基因组学研究起步晚,目前尚未进行大规模研究,亟待加强,这也进一步说明了构建本文库的重要性.
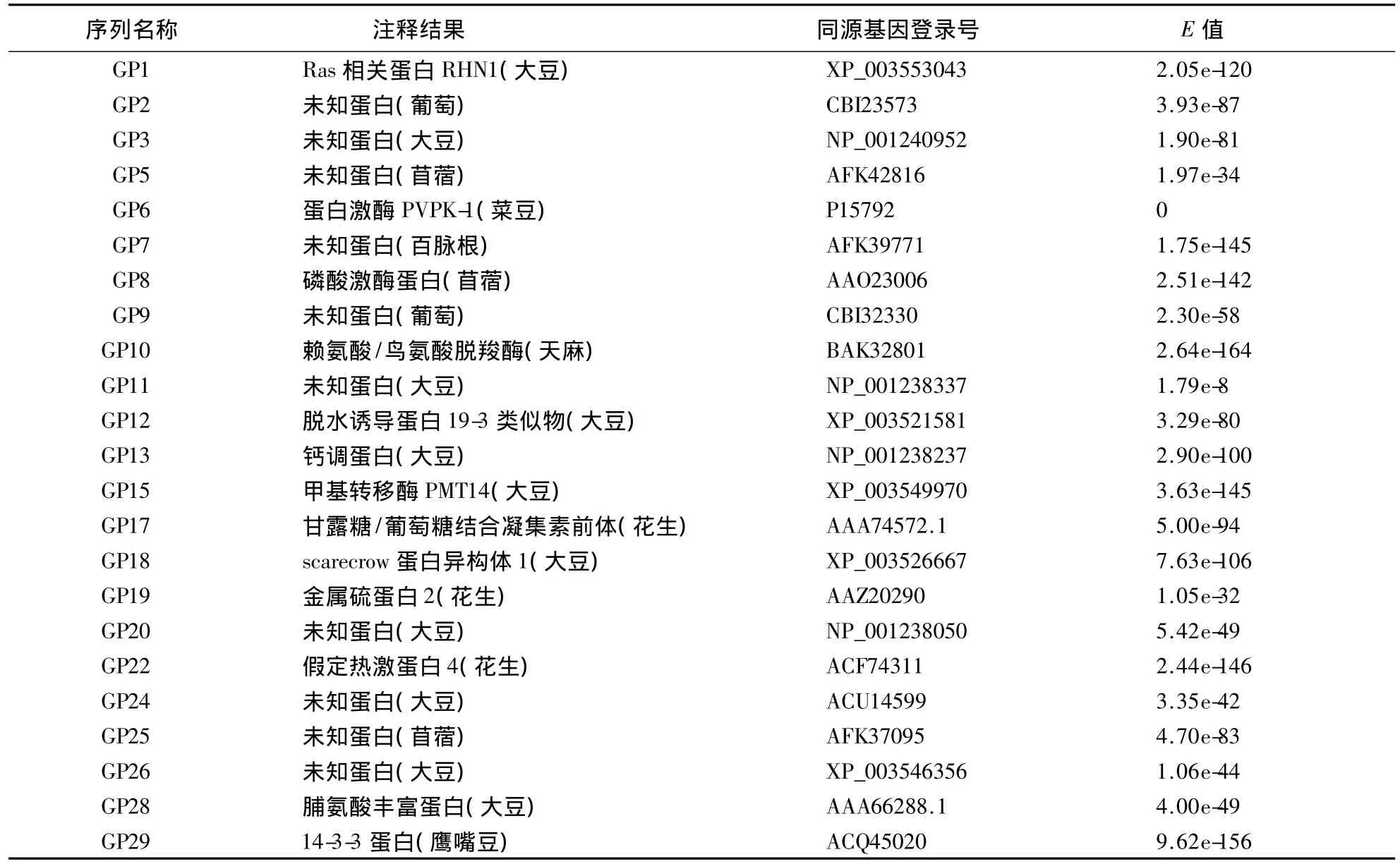
表1 果皮文库BlastX结果Table 1 BlastX result of peanut pericarp full-length cDNA library
将测序结果进行Blast2GO注释和统计分析的结果显示,蛋白结合功能基因最多,此外还含有结构组成、催化活性、分子转导活性、酶调节活性等多种分子功能的基因序列(图5A).参与翻译生物过程的相关基因较丰富,此外还含有生物调控、氧化降解、核糖体生物合成、蛋白代谢、蛋白修饰等生物过程的相关基因(图5C).细胞核基因含量较多,细胞膜、核糖体、细胞质胞液、叶绿体膜、核仁、高尔基体和核质等细胞组分基因均有注释(图5B).可见,本文库所含信息丰富,信息量大,基本包含了花生果皮的各种信息.
3 讨论
黄曲霉毒素一直是世界范围内花生生产、贮藏及加工中面临的严重威胁,被世界卫生组织癌症研究机构划定为一类致癌物质[6],对人畜危害极大.2000年以来,我国多批次出口花生因毒素超标而被退货或销毁,毒素污染已成为花生出口贸易的主要障碍之一[7].闽花6号花生是由本实验室于1997年育成的抗黄曲霉花生新品种,2009年通过国家农作物品种审定委员会审定[8],2006年经福建农林大学植物保护学院用多个高产毒黄曲霉菌株进行接种鉴定试验,结果达到中抗水平[9].本实验室以闽花6号为材料对花生果皮开展了许多相关的研究,应用抑制消减杂交技术构建了花生果皮差异表达文库[10],从中筛选获得一个255 bp的表达序列标签(EST)序列,推测为花生种皮的特异表达基因,可能与细胞内DNA的转录有关[11].近年来,以各种文库为平台研究基因的表达和功能受到人们的重视,许多重要基因如抗性基因[12]、特异表达基因[13-14]、离子通道基因[15]、金属硫蛋白基因[16]均以文库为平台获得.

图5 测序结果Blast2GO注释分析Fig.5 Blast2GO analysis for sequencing results
目前花生对黄曲霉的抗性研究主要集中在花生种皮上.花生对黄曲霉的抗性主要有抗侵染和抗产毒两种类型,其中抗侵染是以花生种皮的完整性及特殊的生化成分抵御黄曲霉菌的侵染和定殖[17].Amaya et al[18]研究指出,抗性品种和易感品种的区别在于其种皮结构和化学组成上的差异,抗性品种具有种脐大、种皮蜡质含量高、栅状层厚、种皮游离氨基酸含量低等特征.种皮蜡质含量和角质层厚度可作为花生抗黄曲霉菌育种的辅助标记性状之一[19].不同抗性水平花生品种种皮纹理超微结构特征不同[20].然而果皮是花生抵抗病原菌如黄曲霉菌侵染的天然屏障,也是第一道防线.因此从花生果皮入手解决花生黄曲霉毒素的污染问题具有更重要的理论及实践意义.目前针对花生果皮对黄曲霉侵染的抗性研究比较少.Delucca et al[21]研究证明,花生壳中类黄酮化合物的作用是通过影响真菌次级代谢途径来抑制黄曲霉毒素产生,黄曲霉在花生壳中的生长不受抑制,而产生的黄曲霉毒素会有所减少.同时花生果皮的完整可防止花生因地老虎等虫害引起的品质和产量下降.除具有抗病虫害功能,花生果皮的多种成分还可用于医药[22-24]、卷烟[25]、食品[23]中.
本试验所构建的花生果皮全长cDNA文库滴度达到1.3×107cfu,重组率为95.5%,基本包含了花生果皮各类基因.插入片段大小多为750-2000 bp,平均大小在1000 bp左右,可用于全长基因的获得.本试验构建了高质量的花生果皮全长cDNA文库,可用于筛选和克隆花生果皮的特异基因,为改善花生果皮对各种病原菌的抗性及花生果壳的综合利用研究提供了平台.
[1]万书波.中国花生栽培学[M].上海:上海科学技术出版社,2003:45.
[2]梁炫强,潘瑞炽,宾金华.花生抗黄曲霉侵染机理的研究进展[J].中国油料作物学报,2000,22(3):77-80.
[3]ACHAR P N,HERMETZ K,RAO S,et al.Microscopic studies on the Aspergillus flavus infected kernels of commercial peanuts in Georgia[J].Ecotoxicol Environ Saf,2009,72(8):2115 -2120.
[4]WANG X G,JI Z J,CAI J,et al.Construction of near isogenic lines for pericarp color and evaluation on their near isogenicity in rice[J].Rice Science,2009,16(4):261 -266.
[5]DONG Y J,XU J L,XIAO K,et al.Genomic regions associated with the degree of red coloration in pericarp of rice(Oryza sativa L.)[J].Journal of Cereal Science,2008,48:556 -560.
[6]ONO E Y,ONO M A,FUNO F Y,et al.Evaluation of fumonisim aflatoxin co-occurrence in Brazilian corn hybrids by ELISA[J].Food Additives and Contaminants,2001,18(8):719 -729.
[7]庞国兴,丁雷霞,姜军,等.青岛地区花生种植地黄曲霉毒素污染调查分析[J].检验检疫科学,2008,18(2):64-65.
[9]庄伟建,石新国,陈华,等.花生新品种闽花6号的选育及优异性状研究[J].福建农业学报,2010,25(3):303-309.
[10]蔡宁波,潘建菁,黄湘文,等.应用抑制消减杂交技术构建花生果皮差异表达文库[J].中国油料作物学报,2007,29(2):46-50.
[11]ZHANG G L,CAI N B,CHEN H,et al.Cloning and characterization of pericarp-and testa-specific,developmentally regulated genes in peanut[J].Genomics and Applied Biology,2009,28(4):640 -644.
[12]张军科,杜敬,李爽,等.白粉菌诱导的华东葡萄环化cDNA文库中抗病基因筛选[J].中国农业科学,2010,43(14):2944-2952.
[13]牛洪斌,覃怀德,王益华,等.水稻谷蛋白的一个新基因克隆及表达分析[J].作物学报,2007,33(3):349-355.
[14]郭俣,刘长青,陆涛峰,等.北京油鸡cDNA文库的构建及purH基因的表达[J].南京农业大学学报:自然科学版,2010,33(4):93-99.
[15]马宁,曾地刚,陈晓汉.凡纳滨对虾基因组fosmid文库的构建及钙离子通道基因的筛选[J].武汉大学学报:理学版,2010,56(3):348 -352.
[16]刘桂丰,褚延广,王玉成,等.星星草cDNA文库构建和金属硫蛋白(MT-1)基因的克隆[J].植物生理学通讯,2005,41(4):424-428.
[17]田立荣,廖伯寿,李玉荣,等.花生抗黄曲霉遗传改良研究进展[J].河北农业科学,2010,14(9):80-83.
[18]AMAYA F J,YOUNG C T,NDRDEN A J,et al.Chemical screening for Aspergillus flavus resistance in peanuts[J].Oleagineux,1980,35:225 -257.
[19]梁炫强,周桂元,潘瑞炽.花生种皮蜡质和角质层与黄曲霉侵染和产毒的关系[J].热带亚热带植物学报,2003,11(1):11-14.
[20]周桂元,梁炫强,李一聪,等.抗黄曲霉花生种皮纹理超微结构的研究[J].中国油料作物学报,1999,14(1):17-19.
[21]DELUCCA A J,PALMGREN M S,DAIGLE D J.Depression of aflatoxin production by flavonoid-type compounds from peanut shells[J].Phytopathology,1987,77(11):1560 -1563.
[22]杨伟强,秦晓春,张吉民,等.花生壳在食品工业中的综合开发与利用[J].花生学报,2003,32(1):33-35.
[23]孙兰萍,林泽梅,许晖,等.花生壳中黄酮物质的提取及对油脂的抗氧化作用[J].资源开发与市场,2008,24(12):1057-1062.
[24]陈孝刚,王志良,王彧,等.花生壳总黄酮提取及其对自由基清除作用的研究[J].辽宁中医药大学学报,2010,12(9):35-38.
[25]朱巍,陈子勇,彤霖,等.花生壳提取物主要成分分析及其在卷烟中的应用[J].香料香精化妆品,2010,6(3):13-17.
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24
中成药(2021年5期)2021-07-21
猪业科学(2021年3期)2021-05-21
现代畜牧科技(2021年2期)2021-03-19
幽默大师(2020年10期)2020-11-10
中华诗词(2019年1期)2019-11-14
猪业科学(2018年4期)2018-05-19
福建中医药(2017年6期)2018-01-09
西南农业学报(2016年4期)2016-05-17
云南畜牧兽医(2014年4期)2014-02-28
