
东北黑木耳野生菌株比较*
2013-03-24 06:37:40张丕奇党阿丽马银鹏韩增华戴肖东
食药用菌 2013年4期
高 娃 张丕奇 党阿丽 马银鹏 韩增华 戴肖东
(黑龙江省科学院微生物研究所,哈尔滨 150010)
东北黑木耳野生菌株比较*
高 娃 张丕奇 党阿丽 马银鹏 韩增华 戴肖东**
(黑龙江省科学院微生物研究所,哈尔滨 150010)
对收集的4个野生黑木耳菌株和6个国家认定的菌株进行形态学和酯酶同工酶的分类鉴定。结果显示:菌丝形态不能有效鉴别供试菌株,野生菌株与其他菌株拮抗显著;酯酶同工酶谱显示,10个菌株得到6个酶谱类型,当遗传相似水平为88%时,测试菌株分为6个类群,4个野生菌株分别聚为不同类群。
黑木耳;野生菌株;酯酶同工酶
黑木耳[Auricularia auricula-Judae(Bull.) Quèl.]属担子菌纲,木耳目,木耳科,木耳属[1],其富含营养,有补血活血,清肺益气,滋补强身的功效,深受广大消费者喜爱。黑龙江省伊春、牡丹江等地区已形成一定水平的规模化生产[2]。
我国黑木耳的人工栽培技术走在了世界前列,而菌种分类鉴定等基础研究严重滞后,菌种质量良莠不齐,品质无从考证,成为黑木耳生产中的严重障碍。20世纪70 年代初蛋白质标记在遗传上应用较多[3],近十年来其在真菌的分类鉴定研究中用于食药用菌品种的分类鉴定的同工酶以酯酶同工酶为主[4~6]。在黑木耳的菌株鉴别、亲缘关系分析等研究中也以酯酶同工酶研究较多[7~9]。本文选用6个本研究所选育的经国家认证的黑木耳品种和4个采集自黑龙江省不同地区的野生菌种为材料,研究分析野生菌株的形态和酯酶同工酶酶谱特性,旨在为野生资源收集和分类鉴定、良种选育、种性鉴定及菌种管理等提供科学依据。
1 材料和方法
1.1 供试菌株 供试菌株共10个,1~6号为国家认定品种;7~10号为野生菌株:“晨明4号”采自黑龙江省伊春市晨明镇,“塔河2号”采自黑龙江省大兴安岭塔河县,“双2号”采自黑龙江省双鸭山市,“加格达奇1号”采自黑龙江省大兴安岭加格达奇区。所有菌株均由黑龙江省科学院微生物研究所菌种保藏中心提供,菌株编号见表1。

表1 供试菌株
1.2 培养基 母种、菌丝培养和拮抗试验培养基(cPDA) :土豆200 g(煮汁1 000 mL),琼脂粉14 g,葡萄糖20 g,磷酸二氢钾3 g,硫酸镁1.5 g,蛋白胨0.5 g,维生素B110 mg,pH自然。同工酶制备液体培养基:元葱200 g(煮汁1 000 mL),蔗糖10 g,阿魏酸0.06 g,pH值自然。
1.3 试验方法
(1)菌丝形态及生长速度。将供试菌株分别同时接种于直径为88 mm的cPDA平板培养基上,接种块大小为2×2(mm),25 ℃恒温培养,观察菌丝生长情况。每3天测一次菌丝生长速度,并记录数据。
(2)菌丝拮抗试验。按照无菌操作的要求,将10个供试菌株点接于cPDA平板培养基上,接种点大小为2×2(mm),每个平板均匀接入 3种菌株,25 ℃下恒温培养,15 d后观察任意两个菌种相交区域之间是否有拮抗现象发生。
(3)菌株同工酶鉴定。①同工酶样品的制备: 分别于250 mL三角瓶中装入80 mL同工酶制备液体培养基,0.10 MPa灭菌20 min,冷却后每瓶中分别接入母种菌块(1.5 mm×1.5 mm)10块,25 ℃静止培养20 d,200目尼龙网过滤收集菌丝体,用蒸馏水冲洗干净,加入0.1 mol/L TBE缓冲液(菌丝体g/缓冲液V:1/1),-20 ℃下冰冻16 h。冰浴研磨成糊状,12 000 r/min低温(4 ℃)离心20 min,取上清液,为同工酶样品。置于-20 ℃冰箱中保存备用。②电泳:垂直板聚丙烯酰胺凝胶电泳,分离胶浓度7.5%,pH 8.9;浓缩胶浓度2.5%,pH 6.7;电极缓冲液 Tris-Glycine系统,pH 8.3;点样40 μL,溴酚兰指示剂,4 ℃下电泳。浓缩胶电压120 V,分离胶电压200 V,电泳4.0 h。③染色:采用醋酸-α-萘酯和坚牢兰染色,缓冲液为0.1 mol/L、pH 6.4的磷酸盐缓冲液。
(4)数据处理与分析。相对迁移率(Rf):电泳结束后先在指示剂移动的位置( 前沿)作标记;染色后,量出指示剂移动的距离和酶带移动的距离。Rf= X2/ X1。X1:固定染色前凝胶中指示剂的迁移距离;X2:固定染色后凝胶中酶蛋白区带的迁移距离。测量Rf时,以酶带的中部位置为准。
相似系数:将凝胶上出现的条带分类,绘图。按照条带有无分别赋值,有带记为1,无带记为0。按Nei 或Li(1997)公式计算菌株间的相似系数Se。其公式为:Se=2Mxy/(Mx+My+2Mxy),式中Mx为x菌株所独有的谱带数,My为y菌株所独有的谱带数,Mxy为x和y菌株所共有的谱带数。
聚类分析:根据供试黑木耳菌株两两间的遗传相似系数用NTSYS-PC软件进行聚类分析,构建黑木耳菌株相似系数树状图谱。
2 结果与分析
2.1 菌丝形态 由表2可知,被试菌株菌落外部特征差异较大,主要表现在菌落形态、气生菌丝、菌丝颜色,及在培养基质中分泌色素情况等方面。1、8号菌丝生长浓密、粗壮,绒毛毡状,气生菌丝生长势旺。2、4、6号菌丝粗壮,成束。3、5、9号菌丝生长稀疏、细弱,绒毛毡状,气生菌丝较弱。7、10号菌丝较弱,成束,气生菌丝长势较弱。除2、4号外,其他菌株都不同程度地产生色素。生长速度最快的为4号,其次是1号,9号菌丝生长最慢,速度差异性分析表明,除9号外,其他处理间差异不显著。
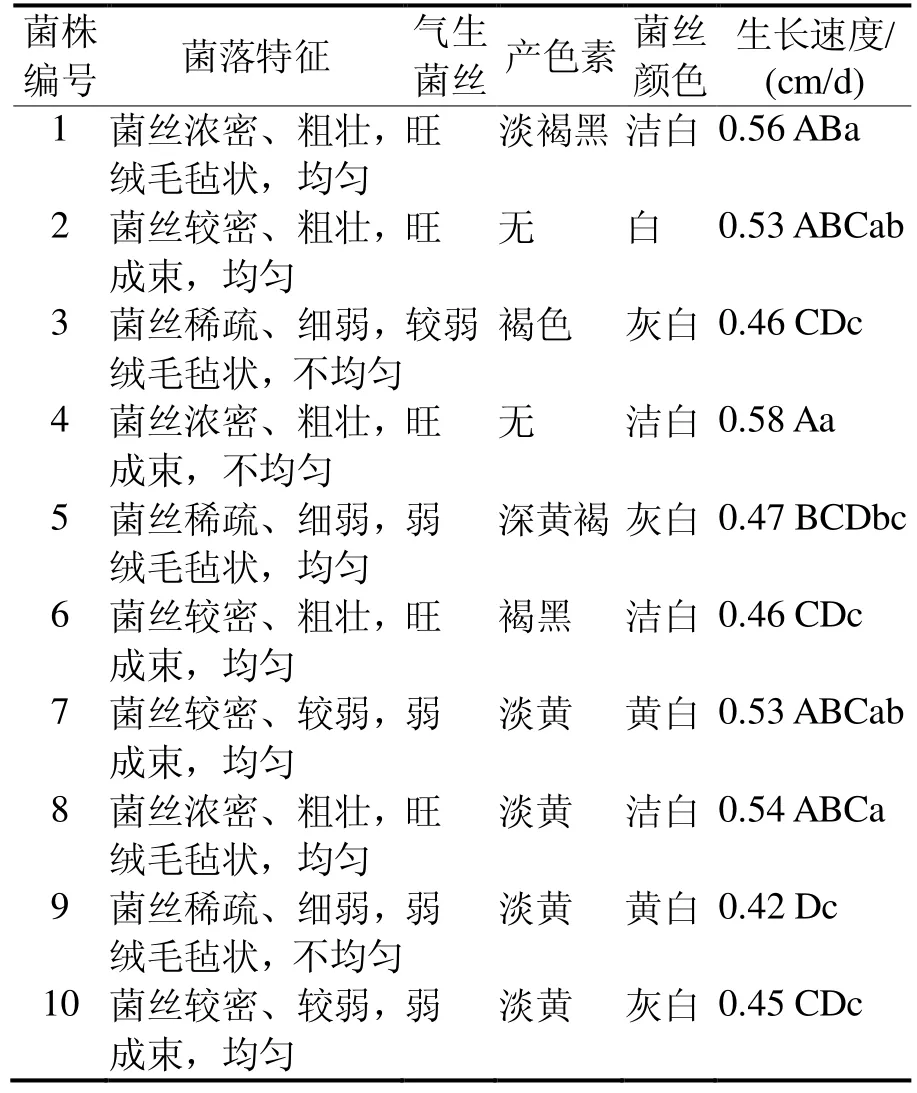
表2 菌株菌丝形态
2.2 菌丝拮抗 试验结果表明,被试菌株除1号和2号,4号和5号,5号和6号间无拮抗外,其他菌株间都存在拮抗反应。1、2号和3号,4号和6号,7号和8号间拮抗不显著,野生菌株与认证菌株间拮抗明显。表明它们与认证菌株亲缘远,为完全独立的异源菌株。
2.3 野生菌株与栽培菌株酯酶同工酶 菌株酯酶同工酶酶谱见图1,10个菌株共检测93条酯酶同工酶谱带,菌株迁移率不同的谱带有15条,所有的谱带均在Rf 0.195~0.766之间,其中Rf 0.195、0.578、0.633、0.668、0.695五处供试菌株都有谱带。10个菌株得到了6个酶谱类型,其中1、2、3号为一个带型;4、5、6号为一个带型。7、8、9、10号菌株各为一个带型。
2.4 酯酶同工酶的相似系数 10个菌株两两间遗传相似系数(Se)的分布范围在0.600~1.000之间,其中2号和3号、5号和6号相似系数为1.000,其他菌株间相似系数较小;相似性最小的为9号和10号,相似系数仅为0.600。
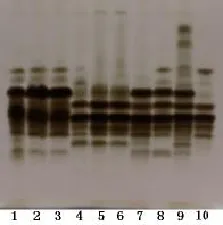
图1 供试菌株的酯酶同工酶酶谱
2.5 聚类分析 NTSYS软件聚类分析得到的酯酶同工酶聚类分析图见图2,遗传相似水平在0.72~1.00之间。从聚类分析树状图可以看出,当遗传相似水平为79%时,测试的10个黑木耳菌株可分为3大类群;当遗传相似水平为81%时,测试菌株可分为4大类群;当遗传相似水平为88%时,测试菌株分为6个类群,第一类群:1、2、3、8号;第二类群:5、6号;第三类群为4号;第四类群为7号;第五类群9号;第六类群为10号。
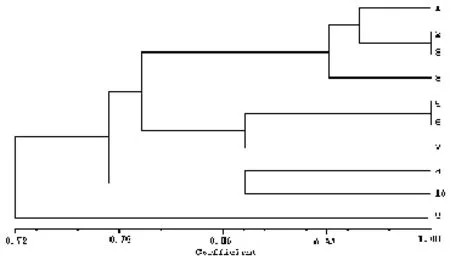
图2 供试菌株的酯酶同工酶聚类分析树状图
3 讨 论
菌丝拮抗与同工酶谱分析皆可在一定程度上反映菌株间的亲缘性,他们间存在着一定的同一性,而菌丝生长形态则很难反映菌丝间的差异。本试验的菌丝拮抗性与酯酶同工酶鉴定结果基本一致,拮抗反应差异显著的菌株,其同工酶谱带差异也较大;反之亦然。菌丝形态受外界环境的影响较大,因此表现型尚不能完全体现其基因型。同工酶判定则是在蛋白、基因水平上反映菌株间的异同,能更准确地反映菌株生化遗传特性。
本试验的野生分离菌株与现存栽培菌株间同工酶的谱带差异显著,又各自独立,表明来源于不同地区的野生菌株在调控基因上存在地域差异。供试的野生菌株地域较远,亲缘关系也较远,证明这些野生菌株在自然界生长的过程中,未受到栽培菌株的杂交干扰,是遗传育种等基础研究的很好材料,应作为珍贵的野生资源保藏。
[1] 杨新美. 食用菌栽培学[M]. 北京:中国农业出版社, 1995: 1~251.
[2] 于文志. 黑龙江省森工企业黑木耳产业发展方向[J]. 中国林副特产, 2007, 91(6): 93-95.
[3] 胡能书, 万贤国. 同工酶技术及其应用[M]. 长沙: 湖南科学技术出版社, 1985: 1-7.
[4] 杜萍, 陈艳秋. 7个桑黄菌株酯酶同工酶的研究[J].中国食用菌, 2006, 25(2): 40-42.
[5] 秦俊哲, 姚艳芳, 孙伟. 18个灵芝菌种的酯酶同工酶分析[J]. 陕西科技大学学报, 2006, 24(6): 65-69.
[6] 傅安涛, 宋爱荣, 丁伟, 等. 15个灰树花菌株的酯酶同工酶分析[J]. 中国食用菌, 2007, 26(1): 40-42.
[7] 韩增华, 张介弛, 戴肖东, 等. 六株黑木耳两种同工酶的研究[J]. 中国食用菌, 2002, 21(6): 42-45.
[8] 唐利华, 郭倩, 王瑞娟, 等. 中国黑木耳主栽菌株酯酶同工酶的研究[J]. 食用菌学报, 2007, 14(4): 37-40.
[9] 张丕奇, 韩增华, 戴肖东, 等. 培养时间对黑木耳酯酶同工酶的影响[J]. 菌物研究, 2008, 6(2): 106-109.
现代农业产业技术体系建设专项资金资助。
高娃,高级工程师,主要从事食用菌菌种收集、分类、鉴定等研究。E-mail:gw_6019@163.com。
**通讯作者:戴肖东,副研究员。 E-mail:heiweihlj@126.com。
猜你喜欢
遗传(2023年8期)2023-08-25 06:59:00
原子与分子物理学报(2021年1期)2021-03-29 07:28:26
宁夏医学杂志(2020年3期)2021-01-21 08:23:38
江苏农业科学(2019年5期)2019-09-02 14:01:46
天然产物研究与开发(2018年8期)2018-09-10 05:48:24
天然产物研究与开发(2018年4期)2018-05-07 06:47:45
广东农业科学(2017年5期)2017-08-29 10:37:31
应用海洋学学报(2014年1期)2014-11-22 07:17:44
中成药(2014年9期)2014-02-28 22:28:55
东北师大学报(自然科学版)(2014年1期)2014-02-27 08:02:15
