
TNF-α诱导SD大鼠胰岛细胞凋亡的研究*
2013-01-10 07:06王功玲
山东第一医科大学(山东省医学科学院)学报 2013年10期
王功玲
(泰山医学院附属泰山医院内分泌二科,山东 泰安 271000)
糖尿病(DM)是以慢性血糖升高为特点的代谢性疾病,伴有蛋白质及脂肪的代谢紊乱,久之可引起肾脏、眼、神经等组织严重的并发症。临床类型主要分为1型糖尿病(T1DM)和2型糖尿病(T2DM),T2DM以外周胰岛素抵抗、胰岛β细胞功能失调及其数量的减少为特征[1],而T1DM中,由于自身免疫导致巨噬细胞和T细胞浸润胰岛组织并分泌一些细胞因子,如IL-1β、TNF-α和IFN-γ等,可诱导β细胞凋亡,从而导致β细胞进行性减少[2]。近年来,TNF-α在糖尿病的发病机制越来越受到人们的重视,它是由激活的单核巨噬细胞和脂肪细胞分泌的一种细胞因子,研究[3]表明,它与糖尿病视网膜病变、胰岛素抵抗及β细胞凋亡都有重要的关系。本课题利用体外细胞培养,观察肿瘤坏死因子-α(TNF-α)对SD大鼠胰岛细胞凋亡的影响并探讨其机制。
1 材料和方法
1.1材料
1.1.1试剂 TNF-α为PeoroTechEc产品,胶原酶P为ROCHE产品,RPMI 1640培养基为GIBCO产品,FICOLL-为Pharmacia产品。Trizol试剂为Promega产品。逆转录聚合酶链式反应试剂盒为Fermentas产品。高保真PCR扩增试剂盒为上海生工生物工程有限公司产品。100 bp DNA Ladder为Fermentas产品。Bcl-2和 Bax引物为invitrogen产品。
1.1.2实验动物 从泰山医学院动物实验研究室购买50只体重约200~250 g成年雄性SD大鼠。
1.2方法
1.2.1大鼠胰岛细胞分离与培养 按董维平[4]法提取胰岛细胞。成年SD大鼠,经臀大肌注射1%戊巴比妥钠麻醉。 消毒,剖腹,经胆总管原位插管,注入预温的1%胶原酶P溶液(含7.5 mmol/L,hepes 10 mmol/L,pH 7.8)8 ml,分离已膨胀的胰腺,38℃震荡消化10 min, 于冰Hanks液中撕碎,使呈细砂状,20目不锈钢网丝过筛,加入冰Hanks液(含10%小牛血清)终止消化,4℃1000 r/min离心3 min,弃上清,同法洗二次,沉淀加25%Ficoll 4 ml混匀,依次加入23%、20%、11%Ficoll各2 ml和Hanks液2 ml,4℃3000 r/min离心20 min,吸出23%~20%,20%~11%界面的胰岛,经Hanks液洗三次,加入含10%小牛血清RPMI 1640培养液24 ml充分混匀。加入24孔培养板,每孔1 ml,于37℃,5%CO2培养。细胞铺成单层时,用台盼蓝鉴定细胞活性>85%。
1.2.2胰岛细胞分组与处理 将培养第7 d的胰岛细胞更换液体, 用Hanks液洗3次, 随机分组(每一组每一浓度为5孔), 依次分空白对照组,10 ng/ml TNF-α组,30 ng/ml TNF-α组,50 ng/ml TNF-α组。处理时加入上述不同浓度TNF-α培养24 h。
1.2.3TUNEL法检测细胞凋亡 按试剂盒说明操作:(1)收集贴壁的胰岛细胞先用4%多聚甲醛室温下固定30~60 min,用PBS和蒸馏水洗涤;(2)用新鲜配制3%H2O2室温处理10 min,蒸馏水洗2 min×3次;(3)加TBS 1∶200新鲜稀释ProteinaseK 37℃消化10~60 s,TBS洗2 min×3次;(4)每片加标记缓冲液20 μl /片,置于湿盒,37℃标记2 h,TBS洗2 min×3次;(5)加封闭液50 μl /片,室温下30 min,甩掉封闭液,不洗;(6)用封闭液1∶100稀释生物素化抗地高辛抗体,50 μl /片加至标本片上,置于湿盒中,37℃反应30 min,TBS洗涤2 min×2次;(7)用TBS 1∶100稀释SABC(1 ml TBS+10 μl SABC), 37℃反应60 min, TBS洗5 min×4次;(8)取1 ml蒸馏水,分别加入DAB试剂各一滴,显色10~30 min,水洗;风干、封片,光镜下观察, 拍照。
1.2.4用RT-PCR检测细胞内Bcl-2和 Bax mRNA表达水平 采用Trizol法提取总RNA,按说明书合成cDNA,在TaqDNA聚合酶催化下行聚合酶链反应。β-actin上游引物序列为5’-AGA GGG AAA TCG TGC GTG AC-3’,下游引物序列为5’-CGA TAG TGA TGA CCT GAC CGT-3’,扩增产物长度为138 bp。扩增条件:94℃预变性4 min,然后在94℃30 s,60℃30 s,72℃30 s 条件下循环30周,最后72℃延伸6 min。Bcl-2上游引物序列为5’-CGG GAG ATC GTG ATG AAG TA-3’,下游引物序列为5’-GGT AGC GAC GAG AGA AGT CA -3’,扩增产物长度为284 bp;Bax上游引物序列为5’-CTG CAG AGG ATG ATT GCT GA -3’,下游引物序列为5’-GAT CAG CTC GGG CAC TTT-3’,预计扩增产物长度为174 bp。扩增条件:94℃预变性3 min,96℃变性50 s,62℃复性50 s,70℃延伸50 s,循环30周,72℃延伸10 min。取6 μl反应产物经1.8%琼脂糖凝胶电泳后摄片,于凝胶图像分析仪扫描,分别计算Bcl-2及Bax与β-actin的灰度比值,以其比值分别代表Bcl-2及Bax mRNA的相对量。

2 结 果
2.1胰岛细胞凋亡形态学观察及凋亡率的计算 显微镜下,正常胰岛细胞细胞核DNA呈黄色,细胞质和核仁的RNA为桔黄色。而凋亡胰岛细胞,细胞核较小,固缩呈较浓的黄绿色,甚至为黄绿色碎块,并可见凋亡小体,具有凋亡细胞的形态特征。对照组很少见到凋亡胰岛细胞,10 ng/ml TNF-α组见到凋亡细胞增加不明显,与对照组比较P>0.05,而30 ng/ml和50 ng/ml组凋亡细胞显著增多,明显高于对照组,两者存在剂量依赖关系(图1~4)。每份样本于高倍镜下随机选取视野,计算100个细胞核,计算凋亡细胞百分率:其凋亡率=凋亡细胞数/计数的胰岛细胞数(表1)。

图1 对照组细胞(×400)

图2 10 ng/ml TNF-α TUNEL图片(×400)

图3 30 ng/m l TNF-α TUNEL图片(×400)

图4 50 ng/ml TNF-α TUNEL图片(×400)

组别凋亡细胞数凋亡细胞比例(%) 对照组9.94±0.653.92±0.4710 ng/ml TNF-α组11.42±0.804.58±0.4930 ng/ml TNF-α组46.52±1.31ab23.02±3.04ab 5 0ng/ml TNF-α组55.22±1.73 abc25.38±2.78abc
a:P<0.01与对照组比较。b:P<0.01与10 ng/ml TNF-α组比较。c:P<0.05与30 ng/ml TNF-α组比较。
2.2不同浓度的TNF-α对胰岛细胞Bcl-2、Bax mRNA表达的影响 各组β-actin mRNA表达水平相差不大,10 ng/ml TNF-α组Bcl-2 mRNA表达水平轻度上升,与对照组相比差异显著(P<0.05),30 ng/ml和50 ng/ml TNF-α组Bcl-2 mRNA表达水平明显下降,明显低于对照组(P<0.05,图5及表2)。而随TNF-α浓度的增加,Bax mRNA的表达水平呈上升趋势,明显高于对照组(P<0.05, 图6及表2)。
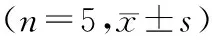
表2 不同浓度的TNF-α对胰岛细胞Bcl-2、Bax mRNA表达的影响
a:P<0.05与对照组比较;b:P<0.05与10 ng/ml TNF-α组比较;c:P<0.01与30 ng/ml TNF-α组比较;d:P<0.01与10ng/ml TNF-α组比较;e:P<0.01与30ng/ml TNF-α组比较。

图5 不同组Bcl-2 mRNA的表达

图6 不同组Bax mRNA的表达
M:Marker;0:正常对照组;10:10 ng/ml TNF-α组;30:30 ng/ml TNF-α组;50:50 ng/ml TNF-α组。
3 讨 论
细胞凋亡(apoptosis) 或细胞程序性死亡( Pro-grammed cell death, PCD) 是组织发育和功能维持的必需生理现象。胰岛β细胞凋亡和功能缺陷在糖尿病的发生和发展中起着非常关键的作用,研究显示2型糖尿病肥胖患者亚临床炎症以及1型糖尿病免疫损伤所导致的高细胞因子水平不仅与外周胰岛素抵抗密切相关[5],而且还可造成胰岛β细胞的功能缺陷和细胞凋亡[6]。尤其是1型糖尿病,目前认为是一种自身免疫性疾病,细胞免疫反应比体液免疫反应更为重要。细胞因子是细胞免疫的主要效应分子,而肿瘤坏死因子是细胞因子的重要成分之一。此外,动物实验证明,将TNF-α基因转入小鼠体内并使其只在胰腺内表达,2个月内即出现明显的胰腺炎,提示胰岛局部TNF-α表达增多与胰岛损伤有密切关系。TNF-α可引起胰岛细胞凋亡,对于这一现象报道也很多,但确切机制目前尚不清楚。
细胞凋亡是一个受基因调控的主动过程。Bcl-2 家族在调节细胞凋亡方面起着重要作用。 Bcl-2家族包括抑凋亡基因及促凋亡基因, 抑凋亡基因包括Bcl-2 , Bcl-xl , Bcl-xr,具有抗凋亡的作用;Bax也是Bcl-2家族的成员之一,属促凋亡基因,可拮抗Bcl-2 的抑凋亡作用。它们之间的表达量的变化与细胞是否存活有密切关系。Bcl-2 基因定位于18q21。Bcl-2基因可在胰岛中表达[7]。在胰腺β细胞株和βTC1细胞中,Bcl-2基因高表达能部分保护细胞因子诱导的细胞凋亡,Bcl-2/ Bax比率决定胰岛细胞对某些凋亡刺激因素的易感性[8]。研究发现,高浓度游离脂肪酸可诱导原代培养的大鼠胰岛细胞和βTc3细胞凋亡,在胰岛细胞凋亡过程中, Bcl-2/ Bax mRNA 表达的比率变化可能起重要作用[9]。
Bcl-2的抗凋亡机制为:(1)Bcl-2主要位于线粒体内膜,这是细胞内活性氧产生的主要场所。可能通过一种抗氧化剂的作用或抑制氧自由基的形成而抑制细胞死亡[10]。(2)Bcl-2蛋白与其它促凋亡蛋白如Bax相互作用,Bax表达水平增加可拮抗Bcl-2的作用,并且促进凋亡,故两者的比值决定细胞的死亡与生存[11]。
本实验以体外培养大鼠胰岛细胞作为受试对象,加入不同浓度的TNF-α处理,采用半定量RT-PCR和TUNEL法分别检测Bax和Bcl-2 mRNA表达变化及细胞凋亡率。结果发现, 10 ng/ml TNF-α组胰岛细胞凋亡数与对照组比较无显著差异(P>0.05),30 ng/ml和50 ng/ml TNF-α组胰岛细胞凋亡数与对照组比较显著增加(P<0.05)。不同浓度的TNF-α引起大鼠胰岛细胞凋亡百分率不同, 浓度越高,细胞凋亡越明显。说明TNF-α对胰岛细胞确实存在毒性,且呈剂量依赖关系。同时还发现:10 ng/ml TNF-α能增加Bcl-2 mRNA表达水平,而30 ng/ml及50 ng/ml TNF-α能显著降低Bcl-2 mRNA表达水平。而以上浓度的TNF-α均能使Bax mRNA表达水平增加。我们推测,低浓度TNF-α在诱导Bax表达的同时,可使Bcl-2表达一过性增加,这可能是细胞对抗凋亡的的一种自我保护机制;随浓度的增加,Bax mRNA表达水平持续增加,而Bcl-2 mRNA 表达水平明显降低,最终导致细胞凋亡。这也与Tran VV等[12]的研究一致,可见,在TNF-α所致的胰岛细胞凋亡中,Bcl-2 /Bax比率变化可能起着重要的作用。
总之,TNF-α所致的胰岛β细胞凋亡在糖尿病的发生和发展中可能起重要的作用,在这一过程中可能有多种基因和不同机制参与,其中Bcl-2/Bax基因表达的变化在上述异常中可能起着重要的作用。
[1] 周晓莉,饶一武,王朝辉,等.新诊断早发DM2患者胰岛素抵抗及胰岛β细胞功能分析[J].放射免疫学杂志,2012,25(4):417-418.
[2] Bruun C, Heding PE, Ronn SG, et al. Suppressor of cytokine signalling-3 inhibits Tumor necrosis factor-alpha induced apoptosis and signalling in beta cells[J].Mol Cell Endocrinol,2009, 13:311-328.
[3] Ohya M,Tanguchi A,Fukushima M, et al.Three measures of tumor necrosis factor alpha activity and insulin resistance in nonobese mLJapanese type 2 diabetic patients[J]. Metabolism, 2005,54(10):1297-1301.
[4] 董维平, 张洪德, 王煜非,等.胰岛移植物质量鉴定方法的研究[J]. 中华器官移植杂志,1998,19(4):205-207.
[5] El-Moselhy MA, Taye A, Sharkawi SS,et al. The antihyperglycemic effect of curcumin in high fat diet fed rats. Role of TNF-α and free fatty acids[J]. Food Chem Toxicol, 2011,49(5):1129-1140.
[6] Kharroubi I, Ladrière L, Cardozo AK, et al. Free fatty acids and cytokines induce pancreatic beta-cell apoptosis by different mechanisms: role of nuclear factor-kappaB and endoplasmic reticulum stress[J]. Endocrinology,2004,145(11):5087-5096.
[7] Hockenbery DM, Zutter M, Hickey W,et al. BCL2 protein is topographically restricted in tissues characterized by apoptotic cell death[J]. Proc Natl Acad Sci, 1991, 88: 6961-6965.
[8] Iwahashi H, Hanafusa T, Eguchi Y,et al.Cytokine-induced apoptotic cell death in a mouse pancreatic beta-cell line: inhibition by Bcl-2[J]. Diabetologia,1996,39:530-536.
[9] Mehmeti I, Lenzen S, Lortz S,et al. Modulation of Bcl-2-related protein expression in pancreatic beta cells by pro-inflammatory cytokines and its dependence on the antioxidative defense status[J]. Mol Cell Endocrinol,2011,332(1-2):88-96.
[10] Tao JL, Wen YB, Shi BY,et al.Endoplasmic reticulum stress is involved in podocyte apoptosis induced by saturated fatty acid palmitate[J]. Chin Med J, 2012,125(17):3137-3142.
[11] DU SC, Ge QM, Lin N,et al. ROS-mediated lipopolysaccharide-induced apoptosis in INS-1 cells by modulation of Bcl-2 and Bax[J]. Cell Mol Biol, 2012,23:1654-1659.
[12] Tran V V,Chen G,Newgard CB,et al.Discrete and complementary mechanisms of protection of beta-cells against cytokine-induced and oxidative damage achieved by bcl-2 overexpression and cytokine selection strategy[J].Diabetes,2003,52(6):1423-1432.
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
实用器官移植电子杂志(2022年5期)2022-03-24
现代临床医学(2021年4期)2021-07-31
生命科学研究(2018年1期)2018-05-29
罕少疾病杂志(2017年2期)2017-02-23
广西林业科学(2016年3期)2016-03-16
中西医结合心脑血管病杂志(2016年20期)2016-03-01
医学研究杂志(2015年12期)2015-06-10
肝胆胰外科杂志(2015年1期)2015-02-27
