
不同光照条件下观光木茎干液流密度季节变化及其相关因素分析
2012-12-14 06:57:14郑怀舟王振兴郑群瑞黄儒珠
植物资源与环境学报 2012年4期
汪 滢,王 健,郑怀舟,王振兴,郑群瑞,黄儒珠
(1.福建师范大学:a.生命科学学院,b.地理科学学院,c.湿润亚热带生态地理过程省部共建教育部重点实验室,福建福州350108;2.福建万木林自然保护区,福建建瓯353100)
光是影响植物生长和分布的重要环境因子之一[1],植物对不同光环境响应策略的差异是植物群落演替和更新的重要内在驱动力之一[2]。木兰科(Magnoliaceae)单种属植物观光木(Tsoongiodendron odorum Chun)分布于江西南部、福建、广东、海南、广西及云南东南部[3-4]。该种分布范围虽广,但种群较小,难以成为森林建群种[4],目前已被列为国家稀有濒危植物[5]。近年来,有关观光木的染色体核型[6]、花器官发育[7]、不同部位的化学成分及生物活性[8-9]、群落特征[4,10-14]、遗传多样性[15-16]、抗性生理[17]以及光合特性[18-19]等方面已有许多研究报道,但尚未见有关观光木蒸腾耗水对光环境变化的响应和适应的研究报道。森林光环境的变化可导致树木的水分生理指标发生相应变化[20],因而研究观光木水分生理指标对不同光环境的响应特征,对于评价和预测在未来森林环境变化条件下这一濒危树种群落的动态变化具有重要的理论和实践意义。
茎干液流是指植物体内由叶片蒸腾作用引起的土壤水分通过植物木质部从根部运输到叶片的上升流。茎干液流技术是研究植物水分生理的有效手段之一,目前应用最广泛、最先进的方法是热技术法。迄今为止,国内研究者大多运用该技术进行高大乔木的植株耗水研究[21-26],但对直径较小的林木幼树的耗水研究较少[27-28]。茎热平衡法(stem heat balance,SHB)主要是利用以热平衡为原理的包裹式茎流探头确定茎干液流运动所产生的热传输和散发至周围环境中的辐射热能量,具有无需标定和直接获得测定结果两大优点[29],对于研究不同光环境下木本植物幼树茎干液流具有明显的优越性。
作者应用Flow 4包裹式植物茎流计和Li-6400便携式光合测定仪测定了在不同遮光条件(100%、40%和8%自然光强)下观光木3年生幼树的茎干液流速率、叶片蒸腾速率、气孔导度以及相关的环境因子(水汽压亏缺、大气温度、大气相对湿度和光合有效辐射强度),并分析了观光木茎干液流的变化特征及其与蒸腾速率、气孔导度和相关环境因子的关系,旨在为观光木的水分生理研究和迁地保育工作提供实验基础和科学依据。
1 研究区概况和研究方法
1.1 研究区概况
本实验在福建万木林自然保护区苗圃内进行。该保护区地理坐标为东经 118°08'22″~118°09'23″、北纬 27°02'28″~27°03'32″,海拔 234 ~556 m,森林覆盖率高达96.1%;该保护区属中亚热带季风气候,年均气温18.7℃,极端最高气温41.4℃,极端最低气温-7.3℃,≥10℃的年均积温为5 889.4℃;年均降水量1 670 mm,年均蒸发量1 466 mm,大气相对湿度81%,降水多集中在春季和夏季;年均霜期75 d,冬季有寒潮影响,但较弱;土壤为花岗岩发育的红壤。植被组成主要为壳斗科(Fagaceae)、樟科(Lauraceae)、山茶科(Theaceae)、金缕梅科(Hamamelidaceae)、木兰科、杜英科(Elaeocarpaceae)、冬青科(Aquifoliaceae)和山矾科(Symplocaceae)等常绿阔叶树种[4,30]。
1.2 材料
供试观光木均为3年生幼树,2008年移植前株高(76.43±5.35)cm、基径(0.88±0.07)cm,2010 年株高(105.78±14.98)cm、基径(1.30±0.30)cm。
1.3 方法
1.3.1 幼苗栽植及遮光处理 于2008年8月在福建万木林自然保护区苗圃内选取长势基本一致的3年生观光木幼树18株,平均分为3组,分别栽植于100%自然光照、40%自然光照(覆盖1层黑色尼龙网布)和8%自然光照(覆盖2层黑色尼龙网布)条件下,定期进行正常田间管理,以保证充足的水、肥及稳定的光照。分别于2010年10月下旬(秋季)、2011年1月中旬(冬季)、2011年4月下旬(春季)和2011年7月下旬(夏季),每处理选择3个样株定株进行有关参数的同步测定。
1.3.2 茎干液流密度测定 采用Flow 4包裹式植物茎流计(美国Dynamax公司,探头为SGA-9探头,直径8~12 mm)进行观光木茎干液流密度(sap flux density,SFD)测定,茎流计的安装与相关参数的数据采集参考许浩等[31]的方法。在观光木茎干基部平滑无节处用磨砂纸轻轻磨去表皮,露出韧皮部,将打磨区清理干净后用游标卡尺精确测定茎干直径;直接将涂有导热硅脂的探头固定在打磨区上,包上保鲜膜,分别裹上保温和防辐射材料后用绑带扎紧;用数据传输电缆连接探头与数据采集器,接通电源,启动茎流计,每隔15 min采集1次数据。在每个季度的测定月份不间断连续测定3 d(晴天),观光木茎干液流密度为3 d的平均值。
1.3.3 其他参数测定 在进行茎干液流密度测定的同时采用Li-6400便携式光合测定仪(美国Li-COR公司)测定同一样株的其他相关参数。于每天6:00至18:00,选择样株冠层中上部完全展开的3片叶片,测定叶片的蒸腾速率(Tr)、气孔导度(Gs)、水汽压亏缺(VPD)以及大气温度(Ta)、大气相对湿度(RAH)和光合有效辐射强度(PAR);每片叶片随机选取3个点(避开主叶脉)进行测量,1h测定1次,连续测定3 d(晴天),结果均为3 d的平均值。
1.4 数据处理
参照文献[32-33]计算SFD,计算公式为:SFD=F/A。式中,A为茎干横截面积;F为茎干液流速率。F值计算公式为:F=Qf/(CP·dT)=(Pin-Qv-Qr)/(CP·dT)。式中,Qf为茎干液流带走的热量;Qv为径向与水流热交换损失的热量;Qr为轴向以辐射形式向四周散发的热量;Pin为茎流计热源的恒定功率;CP为树液比热容(为水的比热容4.186 J·g-1·℃-1);dT为上下2个探针间的温差。Qv、Qr和dT均可通过传感器测量并经自动计算得出。
采用Excel 2003和SPSS 13.0统计分析软件对实验数据进行处理和分析,并采用Origin 8.0软件制作图表。
2 结果和分析
2.1 不同光照条件下观光木茎干液流密度的四季变化特征
在3种光照条件下,一年四季观光木幼树茎干液流密度均呈明显的昼夜周期和连日变化规律,且四季观光木茎干液流日变化趋势基本一致,仅液流启动时间、液流峰值及液流密度大小等特征有一定差异,因而,以秋季观光木茎干液流密度的变化规律(图1)为例进行详细分析。
由图1可见:在不同光照条件下,秋季观光木茎干液流的启动时间均稍有差异。其中,在100%与8%自然光照条件下茎干液流密度的日变化趋势基本一致,基本上表现为液流启动时即达到液流密度的峰值,在10:00至15:00间液流密度虽有小幅波动但比较稳定,15:00后缓慢下降,17:00至17:15液流密度降为0 g·cm-2·h-1,液流停止并持续至次日再次启动;而在40%自然光强条件下液流密度日变化呈明显的“单峰型”,即液流启动后液流密度逐渐增大,峰值出现在9:15至13:30,峰值过后快速下降,在17:30至 18:45 降为 0 g·cm-2·h-1,液流停止。
在不同光照条件下四季观光木茎干液流密度的变化特征见表1。由表1可以看出:观光木茎干液流密度具有明显的光强差异和季节差异。在同一季节,在100%自然光照条件下观光木茎干液流密度的日均值和日峰值均最高,而在8%自然光照条件下均最低。方差分析结果表明:在秋季,观光木茎干液流密度的日均值和日峰值在3种光照条件下均有极显著差异(P<0.01)。在冬季,在100%和40%自然光照条件下,茎干液流密度的日均值无显著差异但日峰值有极显著差异;而在40%与8%自然光照条件下,茎干液流密度日均值有极显著差异(P<0.01),但日峰值无显著差异。在春季,在不同光照条件下茎干液流密度日均值均无显著差异;而在100%自然光照条件下茎干液流密度日峰值与40%和8%自然光照条件下有极显著差异(P<0.01),但后两者间差异不显著。在夏季,因探头安装失误,缺失在40%自然光照条件下茎干液流密度的数据;而在100%与8%自然光照条件下,茎干液流密度日均值和日峰值均有极显著差异(P<0.01)。

图1 秋季不同光照条件下观光木茎干液流密度的日变化曲线Fig.1 Daily change curve of sap flux density of Tsoongiodendron odorum Chun under different light conditions in autumn
从不同季节来看,在100%自然光照条件下,夏季与秋季间观光木茎干液流密度的日均值无显著差异,而其他季节间均有极显著差异(P<0.01);春季与秋季间、夏季与冬季间茎干液流密度的日峰值无显著差异,其他季节间均有极显著差异(P<0.01)。在40%自然光照条件下,在秋、冬、春三季间茎干液流密度的日均值和日峰值均有极显著差异(P<0.01)。在8%自然光照条件下,在夏季和冬季间以及秋季和春季间茎干液流密度的日均值差异均不显著,在其他季节间有极显著差异(P<0.01);而在春季和冬季间以及秋季和冬季间茎干液流密度的日峰值有极显著差异(P<0.01),在其他季节间差异均不显著。
2.2 不同光照条件下观光木叶片水分生理指标的四季变化特征
在3种光照条件下一年四季观光木叶片蒸腾速率和气孔导度的日变化曲线分别见图2和图3。由图2可以看出:仅夏季在100%自然光照条件下观光木叶片的蒸腾速率呈“双峰型”曲线,而在其他光照条件下各季节的叶片蒸腾速率均呈“单峰型”曲线。其中,在40%和8%自然光照条件下同季节叶片蒸腾速率峰值出现的时间基本重叠,秋季和春季均为12:00,冬季为14:00,而夏季略有差异,分别为11:00和12:00。在100%自然光照条件下,秋季和春季叶片蒸腾速率峰值出现的时间均为13:00,冬季为14:00,夏季2个峰值出现的时间分别为11:00和13:00。由图2还可看出:一年四季观光木叶片的蒸腾速率均随光强的减弱而降低。
由图3可以看出:一年四季观光木叶片的气孔导度均随光强的减弱而降低。在40%和8%自然光照条件下,不同季节观光木叶片气孔导度的日变化趋势与蒸腾速率的日变化趋势基本一致,也呈“单峰型”曲线,仅春季气孔导度峰值的出现时间较蒸腾速率提前1 h,其他季节气孔导度峰值的出现时间均与蒸腾速率峰值出现时间同步;在100%自然光照条件下,仅冬季观光木的气孔导度呈“单峰型”曲线,其他3个季节均呈“双峰型”曲线,说明在100%自然光照条件下,秋季、春季和夏季观光木叶片均出现光合“午休”现象。
表1 不同季节在不同光照条件下观光木茎干液流密度特征值比较(±SD)Table 1 Comparison of characteristic values of sap flux density of Tsoongiodendron odorum Chun under different light conditions in different seasons(±SD)
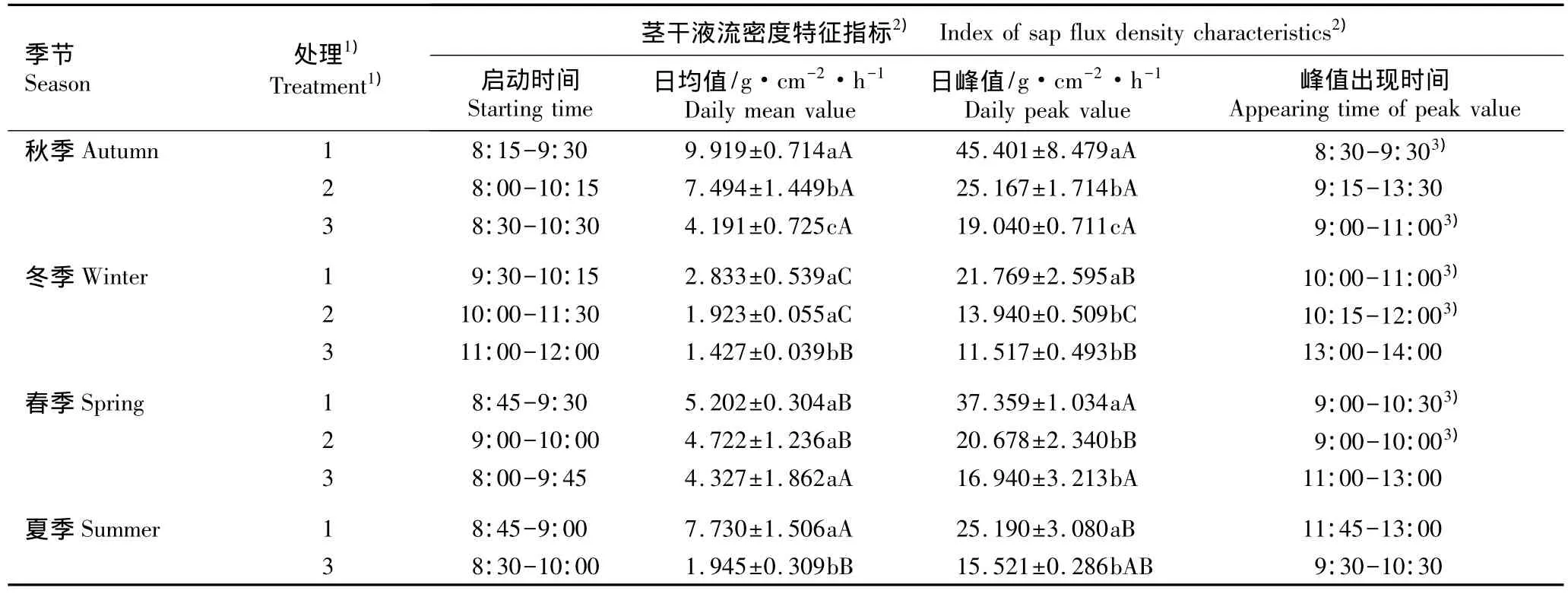
表1 不同季节在不同光照条件下观光木茎干液流密度特征值比较(±SD)Table 1 Comparison of characteristic values of sap flux density of Tsoongiodendron odorum Chun under different light conditions in different seasons(±SD)
1)1:对照,100%自然光照Control,100%of natural light;2:40%自然光照40%of natural light;3:8%自然光照8%of natural light.因探头安装失误,未能获得夏季在40%自然光照条件下茎干液流密度数据 The datums of sap flux density under 40%of natural light condition in summer could not be obtained due to mistake of the probe installation.2)同列中不同的小写字母表示同一季节不同处理间差异极显著(P<0.01)Different small letters in the same column indicate the extremely significant difference among different treatments in the same season(P<0.01);同列中不同的大写字母表示在同一处理条件下不同季节间差异极显著(P<0.01)Different capitals in the same column indicate the extremely significant difference among different seasons under the same treatment(P<0.01).3)茎干液流密度的“晨峰”“Morning peak”of sap flux density.
?

图2 不同季节在不同光照条件下观光木叶片蒸腾速率的日变化曲线Fig.2 Daily change curve of transpiration rate of Tsoongiodendron odorum Chun leaf under different light conditions in different seasons

图3 不同季节在不同光照条件下观光木叶片气孔导度的日变化曲线Fig.3 Daily change curve of stomatal conductance of Tsoongiodendron odorum Chun leaf under different light conditions in different seasons
2.3 观光木茎干液流密度与叶片水分生理指标及环境因子的相关性分析
2.3.1 与叶片水分生理指标的相关性分析 对不同光照条件下不同季节全天6:00至18:00观光木茎干液流密度的平均值与叶片的蒸腾速率和气孔导度的平均值进行相关性分析,相关系数见表2。由表2可见:仅夏季在8%自然光照条件下茎干液流密度与叶片蒸腾速率呈显著(P<0.05)正相关,其他季节在不同光照条件下茎干液流密度与叶片蒸腾速率均呈极显著(P<0.01)正相关。在100%自然光照条件下,仅冬季茎干液流密度与气孔导度无显著相关性,其他季节二者间均呈显著或极显著正相关;在40%自然光照条件下,秋季、春季和冬季茎干液流密度与气孔导度均呈极显著正相关;在8%自然光照条件下,秋季和冬季茎干液流密度与气孔导度呈极显著正相关,春季二者之间呈显著正相关,而夏季二者间无显著相关性。
2.3.2 与环境因子的相关性分析 对不同季节不同光照条件下观光木茎干液流密度与相关环境因子的相关性进行分析,相关系数见表3。由表3可见:叶片水汽压亏缺、大气温度、大气相对湿度及光合有效辐射强度等环境因子与不同光照条件下观光木茎干液流密度的相关性较为复杂,总体上,茎干液流密度与大气相对湿度呈负相关,与其他3个指标呈正相关。
秋季,在不同光照条件下茎干液流密度与叶片水汽压亏缺、大气温度及光合有效辐射强度均呈极显著(P<0.01)正相关,与大气相对湿度呈极显著负相关。
冬季,在100%自然光照条件下,茎干液流密度与光合有效辐射强度呈极显著正相关;在40%自然光照条件下,茎干液流密度与大气温度和光合有效辐射强度呈极显著正相关,与叶片水汽压亏缺呈显著正相关;在8%自然光照条件下,茎干液流密度与叶片水汽压亏缺呈极显著正相关,与大气温度和光合有效辐射强度呈显著(P<0.05)正相关。而在不同光照条件下茎干液流密度与大气相对湿度均呈不显著的负相关。
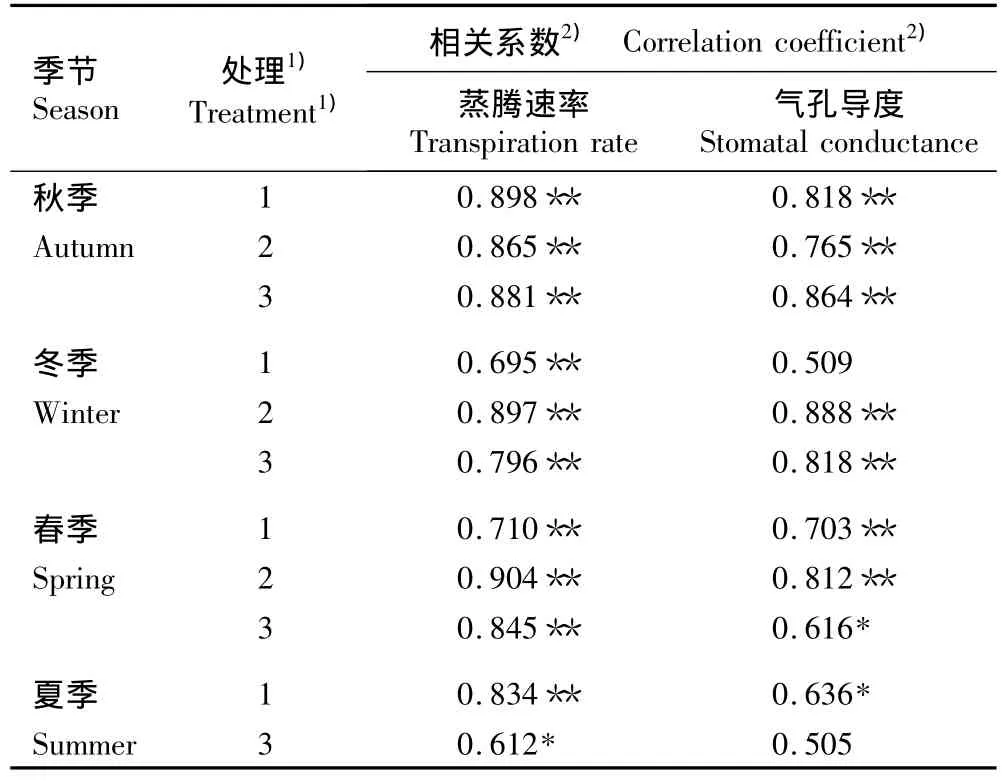
表2 不同季节在不同光照条件下观光木茎干液流密度与叶片蒸腾速率和气孔导度的相关系数Table 2 Correlation coefficient of sap flux density with transpiration rate and stomatal conductance of leaf of Tsoongiodendron odorum Chun under different light conditions in different seasons

表3 不同季节在不同光照条件下观光木茎干液流密度与4个环境因子的相关系数Table 3 Correlation coefficient of sap flux density of Tsoongiodendron odorum Chun with four environment factors under different light conditions in different seasons
春季,在40%和8%自然光照条件下,茎干液流密度与叶片水汽压亏缺、大气温度及光合有效辐射强度呈极显著正相关,与大气相对湿度呈极显著负相关;而在100%自然光照条件下,茎干液流密度仅与光合有效辐射强度呈极显著正相关。
夏季,在100%自然光照条件下,茎干液流密度与叶片水汽压亏缺、大气温度及光合有效辐射强度呈极显著正相关,与大气相对湿度呈极显著负相关;而在8%自然光照条件下,茎干液流密度与光合有效辐射强度呈极显著正相关,与叶片水汽压亏缺呈显著正相关,与大气相对湿度呈显著负相关,与大气温度相关性不显著。
综合分析结果表明:在不同光照条件下,光合有效辐射强度是影响不同季节观光木茎干液流密度的最主要环境因子。
3 讨 论
本研究中,在3种光照条件下一年四季观光木茎干液流密度均存在明显的昼高夜低的准“单峰型”日变化规律,且其日均值和日峰值均随光照强度的降低而减小,均表现为在100%自然光照条件下最高、8%自然光照条件下最低的趋势。除春季外,不同光照条件下观光木茎干液流密度日均值基本上均有极显著(P<0.01)差异。
液流变化是植物自身生理特性与周围环境因子共同作用的结果。本研究中,观光木叶片蒸腾速率和气孔导度均表现为在100%自然光照条件下最高、在8%自然光照条件下最低的趋势,且除了出现“晨峰”现象外,不同光照条件下观光木茎干液流密度与蒸腾速率和气孔导度有相似的日变化规律。在不同光照条件下,各季节观光木茎干液流密度与叶片蒸腾速率均呈显著或极显著的正相关,但各季节气孔导度与观光木茎干液流密度的相关性有一定的差异。其中,在100%自然光照条件下,仅冬季观光木茎干液流密度与气孔导度呈不显著正相关,其他季节间均存在显著或极显著的正相关性;在40%自然光照条件下,秋季、冬季和春季茎干液流密度均与气孔导度呈极显著正相关;在8%自然光照条件下,在秋季和冬季茎干液流密度与气孔导度呈极显著正相关,在春季则呈显著正相关,而在夏季无显著相关性。说明光照是影响蒸腾作用的最主要外界因素,因为光照一方面直接影响植物气孔的开闭,使气孔开放、气孔阻力减小、蒸腾作用加快;另一方面光照可提高叶片温度,使叶内外蒸汽压差增大,从而使蒸腾作用加强。另外,在不同光照条件下,在秋季、冬季和夏季茎干液流密度的日均值总体上有极显著差异,说明蒸腾作用产生的拉力是植物液流上升的主要动力,而根压只有在早春芽叶尚未展开之前才对水分上升起主导作用[34]。此外,上午8:00前观光木茎干液流均没有启动,但在6:00左右植株就有微弱的蒸腾作用,表明液流的启动落后于冠层的蒸腾作用。而余峰等[35]的观察结果显示:在宁夏中部干旱带,柠条(Caragana korshinskii Kom.)的茎干液流与蒸腾作用几乎同步启动。推测造成上述研究结果差异的原因为:在干旱区,白天气温高、空气干燥,柠条蒸腾作用强烈,夜间仍需维持较低的液流以补充白天蒸腾作用引起的水分亏缺;而观光木夜间液流几乎为0 g·cm-2·h-1,白天的蒸腾作用并没有过度消耗观光木体内的水分,这是观光木液流启动时间落后于蒸腾作用的主要原因。
从相关分析结果看,在不同光照条件下观光木茎干液流密度与水汽压亏缺、大气温度及光合有效辐射强度等环境因子均呈正相关,与大气相对湿度呈负相关,但在不同光照条件下不同季节仅光合有效辐射强度与茎干液流密度的相关性达到显著或极显著水平,说明光合有效辐射强度是影响不同光照条件下观光木茎干液流密度的最主要环境因子。这一结果与马长明等[23]和马玲等[24]的研究结果基本一致,但与陈冰冰等[25]和史梅娟等[26]的研究结果有所不同。陈冰冰等[25]认为:空气相对湿度是影响藤本植物瓜馥木〔Fissistigma oldhamii(Hemsl.)Merr.〕茎液流速率的最主要初始环境因子;史梅娟等[26]则认为:水汽压亏缺是影响乔木树种木荷(Schima superba Gardn.et Champ.)、米槠〔Castanopsis carlesii(Hemsl.)Hayata〕和杉木〔Cunninghamia lanceolata(Lamb.)Hook.〕液流的最主要环境因子。造成这一差异的原因可能与植物生长的小生境不同有关,藤本植物和乔木树种的冠层均不受遮挡,因此,光合有效辐射强度不是影响其液流的主要因子。
此外,在3种光照条件下,虽然供试的观光木立地条件完全一致,但在有些季节(如:在100%光照条件下在秋季、冬季和春季,在40%光照条件下在冬季和春季,在8%光照条件下在秋季)茎干液流密度均出现明显的“晨峰”现象,即日出(茎干液流启动)后不久即出现1个陡峰。结合张友焱等[21]、于红博等[22]、Lu 等[36]的研究结果,表明此现象不具有普遍性。然而这一现象是否与光照存在关联,则有待进一步的深入研究。
[1]ALERIC K M,KIRKMAN L K.Growth and photosynthetic responses of the federally endangered shrub,Lindera melissifalia(Lauraceae),to varied light environments[J].American Journal of Botany,2005,92(4):682-689.
[2]HULME P E.Phenotypic plasticity and plant invasions:is it all Jack?[J].Functional Ecology,2008,22(1):3-7.
[3]中国科学院中国植物志编辑委员会.中国植物志:第三十卷第一分册[M].北京:科学出版社,1996:194.
[4]郑群瑞,张兴正,姚清潭,等.福建万木林观光木群落学特征研究[J].福建林学院学报,1995,15(1):22-27.
[5]傅立国,金鉴明.中国植物红皮书:稀有濒危植物(第一册)[M].北京:科学出版社,1992:454-455.
[6]吴文珊,刘剑秋,张清其,等.观光木染色体核型的研究[J].福建师范大学学报:自然科学版,1997,14(1):90-92.
[7]付 琳,曾庆文,徐凤霞,等.观光木的花器官发生[J].热带亚热带植物学报,2007,15(1):30-34.
[8]何开跃,李晓储,张双全,等.观光木叶片挥发油成分及其对超氧阴离子抑制与清除活性研究[J].林业科学研究,2007,20(1):58-62.
[9]宋晓凯,吴立军,屠鹏飞.观光木树皮的生物活性成分研究[J].中草药,2002,33(8):676-678.
[10]吴承祯,洪 伟,郑群瑞.福建万木林保护区观光木群落物种相对多度模型的拟合研究[J].热带亚热带植物学报,2001,9(3):235-242.
[11]潘文钻.福建牛姆林自然保护区观光木群落特征的初步研究[J].林业勘察设计,2002(1):13-16.
[12]邓贤兰,曾晓辉,吴新年,等.井冈山观光木所在群落特征研究[J].井冈山大学学报:自然科学版,2010,31(4):113-117.
[13]许 涵,庄雪影,黄久香,等.广东省南昆山观光木种群结构及分布格局[J].华南农业大学学报,2007,28(2):73-77.
[14]许 涵,黄久香,唐光大,等.南昆山观光木所在群落优势树种的种间联接性[J].华南农业大学学报,2008,29(1):57-62.
[15]黄久香,庄雪影.观光木种群遗传多样性研究[J].植物生态学报,2002,26(4):413-419.
[16]黄久香,庄雪影.华南三地观光木遗传多样性的RAPD分析[J].华南农业大学学报,2002,23(2):54-57.
[17]何开跃,李晓储,黄利斌,等.干旱胁迫对木兰科5树种生理生化指标的影响[J].植物资源与环境学报,2004,13(4):20-23.
[18]孙谷畴,赵 平,曾小平,等.不同光强下焕镛木和观光木的光合参数变化[J].植物生态学报,2002,26(3):355-362.
[19]钟 圣,王 健,郑怀舟,等.福建万木林3种常绿乔木的光合生理特征比较[J].福建师范大学学报:自然科学版,2010,26(4):110-114.
[20]郭志华,张旭东,黄玲玲,等.落叶阔叶树种蒙古栎(Quercus mongolica)对林缘不同光环境光能和水分的利用[J].生态学报,2006,26(4):1047-1056.
[21]张友焱,周泽福,党宏忠,等.利用TDP茎流计研究沙地樟子松的树干液流[J].水土保持研究,2006,13(4):78-80.
[22]于红博,杨 劼,臧春鑫,等.皇甫川流域中国沙棘树干液流日变化及其相关因子[J].生态学杂志,2008,27(7):1071-1076.
[23]马长明,翟明普.干季核桃树干液流特征及其与气象因子的关系[J].林业科学研究,2007,20(6):883-886.
[24]马 玲,赵 平,饶兴权,等.马占相思树干液流特征及其与环境因子的关系[J].生态学报,2005,25(9):2145-2151.
[25]陈冰冰,郑怀舟,王 健,等.瓜馥木茎液流特征及其与环境因子的关系[J].亚热带资源与环境学报,2010,5(1):27-33.
[26]史梅娟,郑怀舟,王 健,等.3种优势树种深秋树干液流特征与环境因子的关系[J].福建师范大学学报:自然科学版,2010,26(3):57-61.
[27]郭树江,徐先英,杨自辉,等.干旱荒漠区沙冬青茎干液流变化特征及其与气象因子的关系[J].西北植物学报,2011,31(5):1003-1010.
[28]王 翠,王传宽,孙慧珍,等.移栽自不同纬度的兴安落叶松(Larix gmelinii Rupr.)的树干液流特征[J].生态学报,2008,28(1):136-144.
[29]岳广阳,张铜会,刘新平,等.热技术方法测算树木茎流的发展及应用[J].林业科学,2006,42(8):102-108.
[30]朱锦懋,姜志林,郑群瑞,等.福建万木林自然保护区森林群落物种多样性[J].生态学杂志,1997,16(2):1-6.
[31]许 浩,张希明,闫海龙,等.塔克拉玛干沙漠腹地多枝柽柳茎干液流及耗水量[J].应用生态学报,2007,18(4):735-741.
[32]STEINBERG S,VAN BAVEL C H M,MCFARLAND M J.A gauge to measure mass flow rate of sap in stems and trunks of woody plants[J].Journal of the American Society for Horticultural Science,1989,114(3):466-472.
[33]严昌荣,ALEC D,韩兴国,等.北京山区落叶阔叶林中核桃楸在生长中期的树干液流研究[J].生态学报,1999,19(6):793-797.
[34]王宝山.植物生理学[M].北京:科学出版社,2004:22.
[35]余 峰,陈 林,王 峰,等.宁夏中部干旱带人工柠条茎流及蒸腾特征研究[J].水土保持研究,2011,18(2):143-147.
[36]LU P,URBAN L,ZHAO P.Granier’s thermal dissipation probe(TDP)method for measuring sap flow in trees:theory and practice[J].Acta Botanica Sinica,2004,46(6):631-646.
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
南京林业大学学报(自然科学版)(2022年1期)2022-11-29 10:10:16
节水灌溉(2021年5期)2021-05-26 06:49:38
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
农业机械学报(2020年3期)2020-04-01 08:11:10
中国农业科技导报(2020年12期)2020-03-15 10:50:08
种子(2019年5期)2019-07-02 11:37:44
天然产物研究与开发(2016年11期)2016-06-15 20:29:15
