
APPβ分泌酶切割位点特异性单链抗体的制备和鉴定
2012-12-06 08:03周丽荣孙红霞黄晓刚
中国药理学通报 2012年3期
黄 巍,周丽荣,周 婷,孙红霞,黄晓刚,刘 桥
(武汉市医学科学研究所基础医学研究室,湖北武汉 430014)
阿尔采末病(Alzheimer’s disease,AD)发病机制复杂,其中 β-淀粉样蛋白(amyloid β protein,Aβ)胞内外异常过量产生、聚集和沉积,是导致AD老年斑形成和神经元毒性的确切病理现象和重要诱因。Aβ由I型跨膜的淀粉样前体蛋白(amyloid precursor protein,APP)经分泌酶水解代谢产生,按作用位点可分为α、β和γ 3种,α和β分泌酶(β-site APP-cleaving enzyme,BACE)先竞争性的水解 APP,然后γ分泌酶再水解剩下的APP片段产生各自代谢产物。Aβ由β和γ分泌酶依次水解APP产生[1],具有毒性;α和γ分泌酶依次水解APP,产生的可溶性sAPPα肽和P3肽段则具有神经营养等作用。若APP的α和β代谢失去平衡,产生过多的Aβ,就容易诱发AD。
APPβ分泌酶切割位点是Aβ产生的关键位点,容易受到周围空间环境影响,如将Aβ抗体与APP结合,当抗体识别位点靠近Aβ的N端,即靠近APP的β切割位点,则Aβ的产生会受到干扰[2],抗体识别位点正好为APPβ分泌酶切割位点序列时则作用更直接[3-4],提示APPβ位点可以作为一个药物设计的良好靶点。我们在研制抗人APPβ分泌酶水解位点的单克隆抗体(mAb)的基础上[5],进一步研制特异性单链抗体(ScFv),ScFv相对mAb具有免疫原性弱,组织穿透力强,更容易通过血脑屏障等特点。APPβ分泌酶切割位点抗体与Aβ抗体在识别序列和减低Aβ机制上都有所不同,前者的目的是阻碍β分泌酶进入作用位点,切割APP产生Aβ,后者则是激活免疫系统,溶解清除已产生的Aβ,见Fig 1。
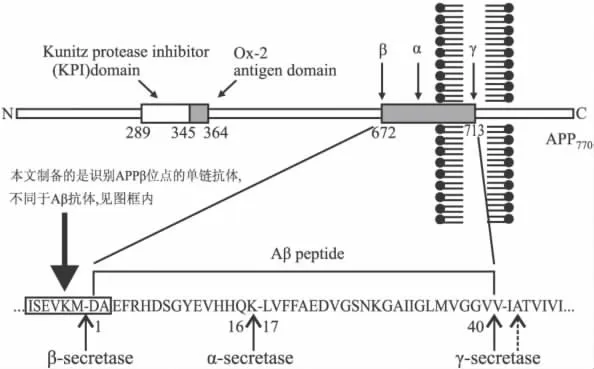
Fig 1 The delineation of APP degradation
1 材料与方法
1.1 材料 分泌抗人APPβ分泌酶水解位点单克隆抗体细胞株1A7和2H10[5],以及稳定转染pEGFP-hAPP751(+4G)的 CHO细胞株(过表达人APP751和GFP融合蛋白)[6]由本室制备保存。限制性内切酶BamH I和EcoR I、pMD19-T克隆载体和逆转录酶M-MLV等为大连宝生物公司产品。引物合成与测序为上海英骏生物技术公司,高保真pfu酶和细胞裂解液购自碧云天生物技术研究所,镍NTA Resin为上海博彩生物科技有限公司产品,增强型HRP-DAB底物显色试剂盒为北京天根生化公司产品。APPβ位点序列多抗原短肽[ISEVKMDA]8由上海波泰生物科技公司合成,抗His的单克隆抗体为Santa Cruz公司产品,抗His的兔多克隆抗体和Aβ肽(1-42)为北京博奥森生物公司产品。
1.2 方法
1.2.1 引物合成 由于含linker序列的引物不利于扩增抗体的重链可变区VH和轻链可变区VL基因,故先设计不含linker序列的引物扩增抗体的VH和VL片段,再以此为模板用含linker序列的引物扩增VH和VL,引物分别为:VHF(重链VH上游引物):5'-ATAGGATCCCAGGTSMARCTGCAGSAGTCWGG-3'(含 BamH I位点;W=A/T,S=G/C,M=A/C,R=A/G);VHB1(重链 VH下游引物,不含 linker序列):5'-TGAGGAGACGGTGACCGTGGTCC-3';VHB(重链 VH下游引物,含 linker序列):5'-GCCAGAGCCACCTCCGCCTGAACCGCCTCCACCTGAGGAGACGGTGACCGTGGTCC-3';VLF1(轻链 VL上游引物,不含 linker序列):5'-GACATCGAGCTCACCCAGTCTC-3';VLF(轻链VL上游引物,含linker序列):5'-CAGGCGGAGGTGGCTCTGGCGGTGGCGGATCGGACATCGAGCTCACCCAGTCTC-3';VLB(轻链VL下游引物):5'-CGCGAATTCCCGTTTGATTTCCAGCTTGGTCCC-3'(含EcoR I位点)。引物VHB1和VLF1分别是VHB和 VLF中3'端序列的一部分,VHB和VLF中5'端部分为含有互补关系的linker序列,VHF和VLB分别含有BamH I和EcoR I酶切位点,便于拼接出来的ScFv构建载体。
1.2.2 VH和VL基因的克隆 分别提取1A7和2H10总 RNA,逆转录合成 cDNA。以 cDNA为模版,分别用VHF/VHB1和VLF1/VLB引物扩增出不带linker的序列,命名为VH1和VL1,回收纯化VH1和VL1片段,再以此为模板,用VHF/VHB和VLF/VLB引物,在高保真pfu酶作用下,分别扩增含linker序列的VH和VL片段。
1.2.3 ScFv的拼接 将VH和VL片段回收纯化,通过重叠延伸PCR(splicing by overlapping extension PCR,SOE-PCR)技术连接起来,即为ScFv片段,其主要过程如下:取等量纯化后的VH和VL片段与适量dNTP和Taq酶混匀,进行PCR反应,条件为:94℃ 1 min,60℃ 1 min,72℃1 min,共进行 7 个循环。然后,不必纯化直接取适量SOE-PCR反应液作为模板,用VHF/VLB引物大量扩增 ScFv片段。将PCR的产物与 pMD19-T克隆载体连接,转化DH5α,挑选阳性克隆测序。
1.2.4 ScFv表达载体的构建 以测序正确的ScFv克隆载体为模板,用VHF/VLB引物再次大量扩增ScFv片段,BamH I和EcoR I双酶切,回收纯化后与同样双酶切的pET-28a表达载体相连接,转化感受态DH5α,通过PCR、BamH I和 EcoR I双酶切和测序鉴定阳性克隆,命名为pET-ScFv(APPβ)。
1.2.5 ScFv的表达纯化及鉴定 将鉴定正确的pET-ScFv(APPβ)质粒转化表达菌株BL21(DE3),筛选阳性菌落进行IPTG诱导表达,超声破碎离心收集上清和沉淀,进行12%SDS-PAGE电泳,考马斯亮蓝染色分析。结果表达产物以包涵体形式居多,目的蛋白带有his标签,通过镍柱纯化和复性,Western blot检测,一抗为抗His的单克隆抗体,增强型HRP-DAB底物显色试剂盒显色。
1.2.6 可溶性ScFv生物活性的分析 通过ELISA分别检测ScFv对人APPβ分泌酶位点序列短肽的结合能力和对Aβ的交叉反应:以APPβ位点序列多抗原短肽[ISEVKMDA]8或 Aβ(1-42)包被酶标板,5 mg·L-1,每孔 100 μl,4℃过夜。洗板后,加可溶性ScFv,然后依次加抗His的兔多克隆抗体、HRP标记抗兔二抗,TMB显色并读取A450值。通过Western blot检测ScFv对人全长APP的结合能力:培养过表达人APP751和GFP融合蛋白的CHO细胞和过表达GFP的CHO细胞,裂解后行10%SDSPAGE电泳,湿转至NC膜,5%脱脂奶粉37℃封闭2 h,可溶性ScFv 4℃孵育过夜,洗膜后加抗His的单克隆抗体37℃ 2 h,洗膜,再加HRP酶标二抗37℃2 h,试剂盒显色。
2 结果
2.1 VH和 VL基因的克隆及 ScFv的拼接 从2H10中扩增出预期的约370 bp的VH1和330 bp的VL1条带,回收纯化后以各自为模板,分别扩增出含linker序列的大小分别约为410 bp和380 bp的VH和VL条带。将VH和VL回收纯化,经SOE-PCR拼接得到约750 bp的ScFv片段,见Fig 2。将2H10的ScFv与pMD19-T克隆载体连接,转化DH5α,挑选阳性克隆测序,测序正确的序列用于ScFv表达载体的构建。从1A7中没有扩增出预期的VH1,该株克隆中止。
2.2 表达载体pET-ScFv(APPβ)的构建和鉴定经筛选得到的阳性重组体命名为pET-ScFv(APPβ),通过PCR可以扩增出约750 bp的条带,BamH I和EcoR I双酶切显示两个片段,分别是约5.5 kb和750 bp,与预计结果相同,见Fig 3。测序分析表明ScFv序列全长744 bp,编码248个氨基酸,轻重链基因序列符合小鼠抗体可变区的结构特点,表达载体阅读框正确,见Fig 4。
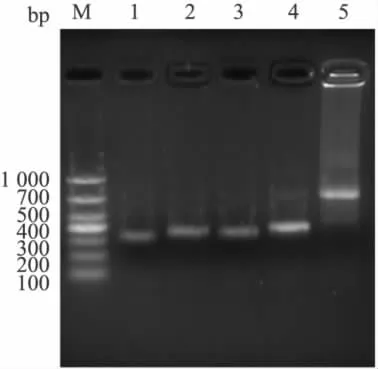
Fig 2 PCR product of VL1,VH1,VL,VHand ScFv gene from hybridoma 2H10
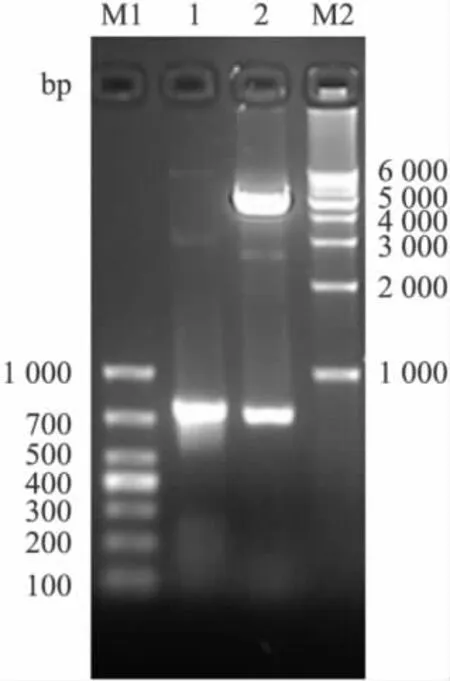
Fig 3 PCR product and the results of reconstruction vector pET-ScFv(APPβ)digested by BamH I and EcoR I
2.3 ScFv的表达纯化和鉴定 诱导表达产物进行12%SDS-PAGE分析,在约29 ku位置处出现新增条带。诱导表达菌经超声破碎离心收集上清和沉淀,进行SDS-PAGE分析,结果表明表达产物主要为包涵体形式。通过镍柱纯化并复性后,可得到分子量约29 ku、纯度在90%以上的条带。目的蛋白通过his标签进行Western
blot检测,结果显示在分子量大小为29 ku处有特异条带,与预期ScFv蛋白情况相符,见Fig 5。
2.4 可溶性ScFv生物活性的分析 ELISA检测结果表明,分别包被APPβ位点序列多抗原短肽[ISEVKMDA]8和Aβ,ScFv能够与前者结合,与 Aβ 的结合反应则未能检测到。过表达人APP751和GFP融合蛋白的CHO细胞和过表达GFP的CHO细胞,裂解后行10%SDS-PAGE电泳,湿转至NC膜,封闭后,依次加可溶性ScFv、抗His单克隆抗体和酶标二抗孵育,增强型HRP-DAB显色。结果过表达APP细胞在约156 ku的地方出现条带,与预期的全长APP(约130 ku)+GFP标签(约26 ku)分子量之和一致,而稳定转染 GFP的细胞无任何条带,排除ScFv与GFP结合的可能,表明ScFv是可以与全长APP结合的,见Fig 6。
3 讨论
目前,有关阻断或清除胞内外异常增多Aβ的研究主要集中在APPβ及γ分泌酶抑制剂或RNA干扰[7-9]、Aβ抗体或疫苗等方向,但存在一些问题,如APP分泌酶除APP外,还有其它在正常生理活动中发挥重要功能的底物,抑制分泌酶活性有可能带来严重的副作用。APPβ分泌酶切割位点抗体为减少Aβ提供新的思路,当其与作用位点结合后,发挥空间位阻效应,阻碍β-分泌酶进入位点,致使Aβ无法产生,此方法通过阻碍水解酶进入特定位点而抑制其特定作用,由于不直接抑制水解酶的活性,特异性高副作用小,可为AD或其他新型药物设计或疫苗研究提供借鉴。
Aβ的产生有两大途径——分泌途径和内吞途径,β-分泌酶和位于高尔基体、细胞表面和内吞体等处的APP均能相互作用,在APP的分泌过程和胞膜APP内吞体形成与循环过程中水解APP生成Aβ[10-11]。APPβ 分泌酶位点单克隆抗体主要在内吞体循环过程中抑制Aβ产生,抑制率约50%[12]。本文进一步研制APPβ分泌酶位点小分子的ScFv,期望以后通过基因转染等方式将其导入胞内,还可在APP的分泌过程中抑制Aβ产生。本文制备的ScFv在抑制Aβ产生的同时还可能具有促进APPα代谢的潜能。虽然有报道指出APPβ位点单克隆抗体不能在阻断BACE作用的同时使APP朝α代谢途径发展[3],但小分子的单链抗体或胞内抗体则不然,不但不会影响α位点水解,还有利于APP与α酶结合使其向α分泌途径发展。这是因为APP的α和β位点距离很近,α酶和β酶(或β位抗体)与相应位点结合时可能出现类似竞争性结合的情况。当β酶或β位单克隆抗体与β位点结合后,由于两者分子较大都会阻碍α酶与α位点的结合,故影响APP的α代谢;而小分子抗体由于空间位阻小,与β位点结合后既阻碍β酶进入β位点又方便α酶与α位点的结合,从而有利于APP的α代谢。关于这一点,识别位点靠近β位的抗Aβ胞内抗体的研究提供了有力的证据[2],研究表明该Aβ抗体对α位点的水解有促进作用,因此,可以推断距离α位点较该Aβ抗体更远的β位点单链抗体或胞内抗体,对APPα分泌酶与α位点结合切割过程将会更有利。

Fig 4 The sequence of pET-ScFv(APPβ)
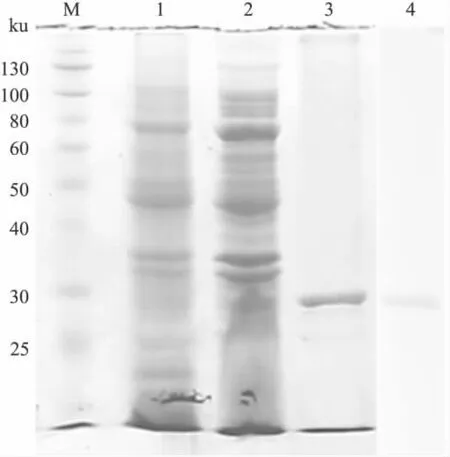
Fig 5 SDS-PAGE analysis and Western blot assay of pET-ScFv(APPβ)

Fig 6 Western blot analysis of the binding of soluble ScFv to the full length of hAPP
本文采用的是传统克隆抗体可变区的方法,即用一组简并引物进行扩增,结果两株杂交瘤细胞中只有2H10可以扩增出VH和VL基因并成功拼接出ScFv序列,而1A7扩增不出预期大小的VH(简并引物扩增VH时,有一条约700 bp的带,资料未显示),说明抗体可变区高度可变,简并引物法不适用于某些抗体可变区序列的扩增,需要采用其他方法钓取抗体的可变区序列。在扩增VH和VL基因片段时,不宜直接使用含linker序列的引物VHB和VLF,否则由于引物太长导致二级结构复杂不利于片段扩增,导致扩增失败或效率过低不利于回收。本文先用不含linker序列的短引物VHB1和VLF1与各自配对引物扩增出目的片段VH1和VL1,经纯化后再各自为模板,用含linker序列的引物扩增相应的VH和VL片段,效果很好。用于扩增VH和VL的模板VH1和VL1必须要纯化,否则不利于VH和VL的扩增;而SOEPCR产物应直接用于PCR扩增ScFv序列,可以扩增出很亮的ScFv带,若纯化后再扩增ScFv,则容易出现带拖尾不集中等情况。ScFv与pET-28a表达载体相连接并在大肠杆菌中诱导表达,载体上His标签与ScFv融合,方便ScFv通过镍柱纯化和复性。ELISA和Western blot检测结果表明,可溶性ScFv与人APPβ分泌酶水解位点序列短肽和全长APP都有结合活性,对Aβ没有交叉反应,可以推断ScFv能够与全长APP的β分泌酶切割位点结合,这为进一步研究ScFv干预分泌酶对APP的水解过程奠定了基础。
[1]李守业,朱 妍,焦 悦,等.阿尔采末病APP分泌酶及其靶向干预研究进展[J].中国药理学通报,2011,27(10):1349-53.
[1]Li S Y,Zhu Y,Jiao Y,et al.Progress of APP secretase and targeted intervention on Alzheimer’s disease[J].Chin Pharmacol Bull,2011,27(10):1349-53.
[2]Paganetti P,Calanca V,Galli C,et al.beta-site specific intrabodies to decrease and prevent generation of Alzheimer’s Abeta peptide[J].J Cell Biol,2005,168(6):863-8.
[3]Arbel M,Yacoby I,Solomon B.Inhibition of amyloid precursor protein processing by beta-secretase through site-directed antibodies[J].Proc Natl Acad Sci,2005,102(21):7718-23.
[4]Thomas R S,Liddell J E,Kidd E J.Anti-amyloid precursor protein immunoglobulins inhibit amyloid-β production by steric hindrance[J].FEBS J,2011,278(1):167-78.
[5]黄 巍,庞蓓蓓,周 婷,等.抗人APPβ水解位点单克隆抗体的制备及其初步鉴定[J].细胞与分子免疫学杂志,2010,26(12):1235-7.
[5]Huang W,Pang B B,Zhou T,et al.Preparation and characterization of monoclonal antibody against β-site of human APP[J].J Cell Mol Immunol,2010,26(12):1235-7.
[6]黄 巍,庞蓓蓓,熊 波,等.Kozak序列(+4G)对携带GFP标签的人APP751表达的影响[J].生物技术通讯,2011,22(2):149-53,196.
[6]Huang W,Pang B B,Xiong B,et al.Effect of kozak sequnce+4G on the expression of hAPP751 and GFP fusion gene[J].Lett Biotechnol,2011,22(2):149-53,196.
[7]Elvang A B,Volbracht C,Pedersen L Ø,et al.Differential effects of gamma-secretase and BACE1 inhibition on brain Abeta levelsin vitroandin vivo[J].J Neurochem,2009,110(5):1377-87.
[8]孙 琰,周文霞,张永祥.RNAi技术在阿尔采末病防治研究中的应用[J].中国药理学通报,2009,25(11):1408-11.
[8]Sun Y,Zhou W X,Zhang Y X.Applications of RNAi technology in Alzheimer’s diseases[J].Chin Pharmacol Bull,2009,25(11):1408-11.
[9]张红梅,张 雄,李 昱.姜黄素对阿尔采末病中APP淀粉样酶切途径的调控作用[J].中国药理学通报,2009,25(3):361-6.
[9]Zhang H M,Zhang X,Li Y.The regulation of Curcumin on the amyloidogentic pathaway of APP in Alzheimer’s disease[J].Chin Pharmacol Bull,2009,25(3):361-6.
[10]Yan R,Han P,Miao H,et al.The transmembrane domain of the Alzheimer’s beta-secretase(BACE1)determines its late Golgi localization and access to beta-amyloid precursor protein(APP)substrate[J].J Biol Chem,2001,276(39):36788-96.
[11]Kinoshita A,Fukumoto H,Shah T,et al.Demonstration by FRET of BACE interaction with the amyloid precursor protein at the cell surface and in early endosomes[J].J Cell Sci,2003,116(Pt 16):3339-46.
[12]Thomas R S,Liddell J E,Murphy L S,et al.An antibody to the beta-secretase cleavage site on amyloid-beta-protein precursor inhibits amyloid-beta production[J].J Alzheimers Dis,2006,10(4):379-90.
猜你喜欢
计算机应用与软件(2022年6期)2022-07-12
自然灾害学报(2022年2期)2022-05-10
中国学校体育(2021年10期)2021-04-26
昆明医科大学学报(2021年2期)2021-03-29
中国现代医药杂志(2020年10期)2020-12-14
中学化学(2019年3期)2019-07-08
天然产物研究与开发(2018年7期)2018-08-21
中学化学(2016年2期)2016-05-31
课程教育研究·下(2016年2期)2016-03-25
实验流体力学(2014年6期)2014-07-10
