
肿瘤自噬效应及其在非小细胞肺癌分子靶向治疗中的潜在应用
2012-12-06 08:03陈美娟
中国药理学通报 2012年3期
陈美娟,詹 瑧,张 旭,2
(1.南京中医药大学基础医学院中西医结合基础学科,2.江苏省方剂研究重点实验室,江苏 南京 210046)
肺癌是严重危害人类健康的恶性肿瘤之一,其中非小细胞肺癌(non-smallcelllungcancer,NSCLC)约占肺癌的80%。目前临床上常用的手术、放化疗等治疗手段其疗效仍然相当不足,临床治疗失败的主要原因是复发和发生重要器官的转移,因此,迫切需要寻找其它治疗手段以弥补目前治疗的不足。
1 NSCLC分子靶向治疗的主要方向及存在问题
随着分子生物学的发展和对癌症发生机制的深入研究,以特定分子为靶点的抗肿瘤药物逐渐受到关注。例如表皮生长因子受体(epidermal growth factor receptor,EGFR),由于其在多种上皮肿瘤中均过度表达,而且在调节肿瘤细胞的生长、修复和生存及新生血管的生成、侵袭和转移中具有重要的作用,因此成为抗肿瘤药物的重要靶点之一。由于大多数NSCLC都表达EGFR及其天然配体,目前发现约62%的鳞癌和腺癌中存在EGFR的过度表达[1],并且EGFR的活化可进一步激活对肿瘤的发生、发展都起关键作用的Ras/MAPK、PI3K/Akt和 JAK/STATs等3 条信号通路[2],因此针对EGFR的分子靶向治疗成为NSCLC临床中治疗的一个重要方向。以吉非替尼(gefitinib)和厄洛替尼(erlotinib)为代表的表皮生长因子受体酪氨酸激酶抑制剂(EGFR-TKI)和以西妥昔(cetuximab)为代表的EGFR单克隆抗体已应用于NSCLC的临床治疗,并且显示出了一定的疗效,但随着天然或获得性抵抗的发生,其疗效会大大降低。据最新报道,这些药物产生临床耐药性某种程度上和它们诱导肿瘤细胞产生自噬(autophagy)作用有关[3-4]。
2 自噬在肿瘤治疗中的双重作用
自噬是溶酶体对细胞内的部分细胞质、细胞器等进行一系列降解过程的统称。其形态学的最主要特征是细胞内出现大量具有双层膜的内部存在胞质及细胞器的吞噬泡。自噬在肿瘤的发生、发展中具有双重的作用。在某些情况下自噬可能成为保护肿瘤细胞免受损伤的应激保护机制,如当肿瘤细胞缺乏营养供应时,尤其在肿瘤的中心地带,细胞内自噬作用会增加,通过降解蛋白质和细胞器可以给其他肿瘤细胞供应氨基酸、脂肪酸和核苷酸等营养物质以提高其生存的机会和侵袭的能力[5]。因此,在某些情况下抑制自噬可以增加药物的抗癌作用。如Wu等[6]发现在人恶性胶质瘤细胞中,抑制自噬会明显降低胶质瘤细胞在营养被剥夺状况下的生存能力,而通过过表达真核延长因子-2激酶(eEF-2K)提高自噬水平,可以提高肿瘤细胞在营养缺乏状况下的生存能力,并且发现mTOR通路参与eEF-2K对自噬的调节。Amaravadi等[7]给动物接种缺少核p53基因从而能抵抗凋亡的肿瘤细胞造成淋巴瘤动物模型,发现动物整体水平通过它莫西芬诱导激活p53后,肿瘤先衰退,然后出现复发。进一步研究发现,p53的激活很快伴随凋亡细胞的出现,但在存活的细胞中产生了自噬。而通过氯喹或针对Atg5进行shRNA干扰抑制自噬,会提高肿瘤细胞p53激活的水平和烷化剂诱导肿瘤细胞死亡的能力。Carew等[8]也发现,通过氯喹阻断自噬,可以提高组蛋白脱乙酰酶抑制剂——SAHA诱导的过氧化物的产生水平,促进其重新定位,并明显增加其在溶酶体蛋白酶内的含量,减少其底物硫氧还蛋白的表达,从而提高其抗癌活性。另外,在某些放化疗治疗中,自噬也成为肿瘤细胞逃避损伤的机制[9-10]。
与上述自噬保护肿瘤细胞的情况相反,许多研究发现自噬也可以在抗肿瘤方面发挥重要作用。因为自噬可以通过清除受损细胞器,避免有害自由基及突变的发生,从而避免更大范围的损伤;还可以限制DNA损伤,维持基因组完整性;另外还能抑制细胞生长,诱发肿瘤细胞发生凋亡式程序性死亡。如Scott等[11]发现,Atg1高表达所诱导的自噬可使肿瘤细胞明显缩小,从而控制其生长。同时发现自噬可诱发肿瘤细胞死亡,这种细胞死亡具有明显的凋亡特征。在其他的疗法或药物抗肿瘤的过程中,也发现自噬过程发挥了积极作用,物理疗法如Kessel等[12]报道的光动力疗法;抗癌药物如烷化剂——替莫唑胺(temozolomide)、顺铂;mTOR抑制剂——rapamycin、temsirolimus、everolimus;天然药物——伴刀豆球蛋白A(Con A)、姜黄素、染料木黄酮(genistein)、维生素 D 类似物;NF-κB 抑制剂——BAY11-7082,Parthenolide;植物生物碱——紫杉醇、长春碱;甚至还有其他一些化合物如 As2O3、糖皮质激素、Nelfinavir等[13]。
3 自噬效应在非小细胞肺癌分子靶向治疗中的潜在应用
在NSCLC临床常用药物gefitinib和erlotinib抗药性产生机制的研究中,Han等[3]发现,gefitinib和erlotinib可通过抑制PI3K/Akt/mTOR信号通路诱导肺腺癌细胞A549和H1299发生较高水平的自噬,甚至在用siRNAs干扰降低EGFR的表达后,gefitinib、erlotinib依然能诱导自噬。而当用自噬抑制剂氯喹(chloroquine,CQ)或用siRNAs对自噬相关基因Atg5和Atg7进行干扰后,则会明显增加gefitinib和erlotinib的细胞毒性。另外,Li等[4]也报道,西妥昔单抗的临床抗药性的产生也和其诱导自噬相关,其主要也是通过抑制PI3K/Akt/mTOR信号通路,诱导肺癌细胞产生自噬,细胞自噬的同时还伴随HIF-1α和Bcl-2水平的降低。用PI3K-Ⅲ抑制剂3-MA可以抑制西妥昔单抗诱导的自噬,而敲除自噬调节基因beclin1或Atg7,或用溶酶体抑制剂氯喹处理细胞会增加西妥昔单抗诱导的细胞凋亡。
由此表明自噬作用在NSCLC的临床治疗中同样是一个不可忽略的重要环节,在NSCLC治疗的各个阶段都应考虑到自噬作用的影响。
在肿瘤发生的早期阶段,自噬可表现出抑制肿瘤的作用,因此在肺癌的早期诊断和治疗中,应将自噬相关基因beclin1、p53等纳入肺癌的临床分型指标体系。其中,beclin1是自噬最主要的正调节因子,在多种人类癌细胞中发现beclin1的缺失[14]。此外,值得重视的是对自噬具有双重调节能力的抑癌基因p53。在正常情况下p53为野生型(wt p53),而在大部分癌细胞包括肺癌细胞中,p53表现为突变型(mt p53)[15]。在NSCLC中,p53突变的比例约占50%,而且其发生水平在鳞癌中相对最高,在腺癌中较低[16-19]。p53对自噬的双重调节作用主要表现在:在细胞核中,p53作为核转录因子,通过与 AMPK、DAPK-1、DRAM(damage-regulated autophagy modulator)、Bad、Bax、BNIP3、PUMA、Sestrin1/2和TSC2等自噬调节分子的基因启动子区结合,可以促进相关基因的转录,从而促进自噬;而在细胞质中,p53不依赖于其核转录因子的角色,而主要是通过激活mTORC1,抑制自噬。而且,在营养缺乏或缺氧的情况下,增加的自噬可能促成p53缺失的癌细胞的生存[20]。
由于在肿瘤发展的后期,自噬可能对肿瘤的进展起促进作用,如处于肿瘤组织中心部位的肿瘤细胞可以通过自噬在低氧供应和低营养状态下获得生存;另外,自噬还可通过封闭凋亡通路以保护某些肿瘤细胞的抗癌治疗。在该类情况下,则应该抑制自噬、激活凋亡,从而让肿瘤细胞对治疗手段变得敏感。
另外,还可以有针对性地探索一些单纯诱导肿瘤细胞发生自吞噬式细胞死亡(即Ⅱ型程序性细胞死亡)的药物和治疗手段。寻找该类药物的方向主要从调控自噬的相关信号通路和相关分子靶标入手。目前发现正向调节自噬的信号通路或相关分子主要有:Ras/Raf1/ERK1/2、ClassⅢPI3K/Beclin1、TRAIL-R(TNF-related apoptosis-inducing factor)/TNF-R/FADD(Fas-associated death domain protein),LKB1/AMPK、JNK、DAPK(death-associated protein kinase),DRP-1(death-associated related protein kinase-1);负向调节自噬的信号通路主要有:ClassIPI3K/AKT/mTOR/p70s6k、p38MAPK等(Fig 1)[21]。
其中,在负向调节自噬的信号通路中,mTOR是最主要的信号分子。mTOR是哺乳动物雷帕霉素的靶点,其对细胞的生长、增殖、存活都具有重要的调节作用。mTOR既可以与 Raptor、mLST8和 PRAS40组成 mTOR复 合 体 1(mTORC1),也可以和 Rictor、mSIN1和 Protor-1组成 mTOR复合体2(mTORC2),其中只有mTORC1对雷帕霉素敏感。
在mTORC1的上游,调节mTORC1的信号通路主要有3条:即 ClassIPI3K/AKT、Ras/Raf1/ERK1/2和 LKB1/AMPK。其中前两条通路激活 mTORC1,最后一条通路抑制mTORC1。PI3K/Akt对mTOR的激活一是通过抑制TSC1/TSC2复合体来激活 Rheb,从而激活 mTORC1[22];二是通过磷酸化激活mTORC1的结合蛋白PRAS40,使其与mTORC1解离,同时结合 14-3-3蛋白,从而 mTORC1被激活[23]。LKB1(丝氨酸/苏氨酸激酶11)下调mTORC1主要是在细胞遇到能量短缺压力时,被PARP-1[poly(ADP-ribose)polymerase-1]激活的 LKB1首先将 AMPK磷酸化,然后活化的AMPK将TSC2磷酸化激活,活化的TSC2再激活 Rheb的GAP,从而抑制 mTORC1[24]。
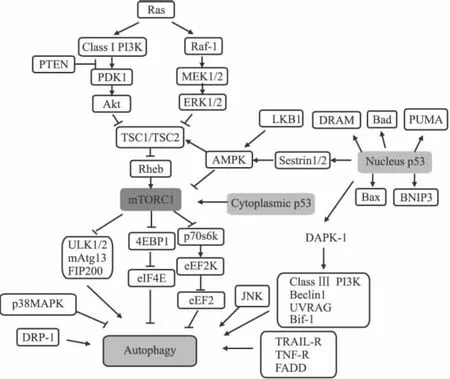
Fig 1 Diagram of autophagy signaling
在mTORC1的下游,mTORC1也通过3条途径影响自噬:其一,通过阻止哺乳动物自噬相关蛋白13(mAtg13)和ULK的结合,导致 ULK稳定性下降,使依赖于 ULK的FIP200的磷酸化被抑制,从而诱导自噬;其二,通过促进4EBP1的磷酸化,利于其与真核翻译起始因子4E(eIF4E)解离,从而上调cap依赖的翻译过程,抑制自噬;其三,真核延长因子2(eEF2)可以促进肽链延长,但eEF2激酶(eEF2K)可磷酸化eEF2并使其失活,从而抑制肽链延长。但磷酸化的p70s6k可以磷酸化eEF2K,使上述活性失效,从而促进翻译。因此mTORC1通过磷酸化p70s6k,促进翻译,抑制自噬[25]。
4 结语
总之,在NSCLC的临床治疗中,应充分考虑并利用好自噬效应,适当情况下,可以将其作为NSCLC分子靶向治疗的方向之一,从而一方面使各种治疗手段的临床疗效得以提高,另一方面结合自噬相关信号通路,开发出新的靶标治疗药物,使得它们共同为NSCLC的临床治疗更有效地发挥作用。
[1]Hirsch F R,Varella-Garcia M,Bunn P A Jr,et al.Epidermal growth factor receptor in non-small-cell lung carcinomas:correlation between gene copy number and protein expression and impact on prognosis[J].J Clin Oncol,2003,21(20):3798-807.
[2]熊 飞,詹 瑧,唐于平,等.PI3K/Akt信号转导通路在非小细胞肺癌中的作用[J].中国药理学通报,2010,26(10):1264-7.
[2]Xiong F,Zhan Z,Tang Y P,et al.The effect of PI3K/Akt signal transduction pathway in non-small cell lung cancer[J].Chin Pharmacol Bull,2010,26(10):1264-7.
[3]Han W D,Pan H M,Chen Y,et al.EGFR tyrosine kinase inhibitors activate autophagy as a cytoprotective response in human lung cancer cells[J].Plos One,2011,6(6):e18691.
[4]Li X Q,Fan Z.The EGFR antibody cetuximab induces autophagy in cancer cells by downregulating HIF-1α and Bcl-2 and activating the beclin-1/hVps34 complex[J].Cancer Res,2010,70(14):5942-52.
[5]刘 畅,郝淑玲,于忠和.细胞自噬与肺癌关系研究进展[J].
临床肺科杂志,2011,16(6):916-8.
[5]Liu C,Hao S L,Yu Z H.Research progresses on relationship between autophagy and lung cancer[J].J Clin Pulm Med,2011,16(6):916-8.
[6]Wu H,Yang J M,Jin S,et al.Elongation factor-2 kinase regulates autophagy in human glioblastoma cells[J].Cancer Res,2006,66(6):3015-23.
[7]Amaravadi R K,Yu D,Lum J J,et al.Autophagy inhibition enhances therapy-induced apoptosis in a Myc-induced model of lymphoma[J].J Clin Invest,2007,117(2):326-36.
[8]Carew J S,Nawrocki S T,Kahue C N,et al.Targeting autophagy augments the anticancer activity of the histone deacetylase inhibitor SAHA to overcome Bcr-Abl-mediated drug resistance[J].Blood,2007,110(1):313-22.
[9]Lomonaco S L,Finniss S,Xiang C,et al.The induction of autophagy by gamma-radiation contributes to the radioresistance of glioma stem cells[J].Int J Cancer,2009,125(3):717-22.
[10]O’Donovan T R,O’Sullivan G C,McKenna S.Induction of autophagy by drug-resistant esophageal cancer cells promotes their survival and recovery following treatment with chemotherapeutics[J].Autophagy,2011,7(6):509-24.
[11]Scott R C,Juhsz G,Neufeld T P.Direct induction of autophagy by Atg1 inhibits cell growth and induces apoptotic cell death[J].Curr Biol,2007,17(1):1-11.
[12]Kessel D,Vicente M G H,Reiners J J.Initiation of apoptosis and autophagy by photodynamic therapy[J].Autophagy,2006,2(4):289-90.
[13]Levy J M,Thorburn A.Targeting autophagy during cancer therapy to improve clinical outcomes[J].Pharmacol Therapeut,2011,131(1):130-41.
[14]Mizushima N,Levine B,Cuervo A M,et al.Autophagy fights disease through cellular self-digestion[J].Nature,2008,451(7182):1069-75.
[15]Vousden K H,Lane D P.p53 in health and disease[J].Nat Rev Mol Cell Biol,2007,8(4):275-83.
[16]Bodner S M,Minna J D,Jensen S M,et al.Expressionof mutant p53 proteins in lung cancer correlates with the class of p53 gene mutation[J].Oncogene,1992,7(4):743- 9.
[17]Takahashi T,Nau M M,Chiba I,et al.p53:a frequent target for genetic abnormalities in lung cancer[J].Science,1989,246(4929):491-4.
[18]Kishimoto Y,Murakami Y,Shiraishi M,et al.Aberrations of the p53 tumor suppressor gene in human non-small cell carcinomas of the lung[J].Cancer Res,1992,52(17):4799- 804.
[19]Tammemagi M C,McLaughlin J R,Bull S B.Meta-analyses of p53 tumor suppressor gene alterations and clinicopathological features in resected lung cancers[J].Cancer Epidem Biomar,1999,8(7):625-34.
[20]Gao W,Shen Z,Shang L,Wang X.Upregulation of human autophagy-initiation kinase ULK1 by tumor suppressor p53 contributes to DNA-damage-induced cell death[J].Cell Death Differ,2011,18:1598-607.
[21]Codogno P,Meijer A J.Autophagy and signaling:their role in cell survival and cell death[J].Cell Death Differ,2005,12:1509- 18.
[22]Sini P,James D,Chresta C,et al.Simultaneous inhibition of mTORC1 and mTORC2 by mTOR kinase inhibitor AZD8055 induces autophagy and cell death in cancer cells[J].Autophagy,2010,6(4):553-4.
[23]Li M,Jiang X,Liu D,et al.Autophagy protects LNCaP cells under androgen deprivation conditions[J].Autophagy,2008,4(1):54-60.
[24]Vander Haar E,Lee S I,Bandhakavi S,et al.Insulin signaling to mTOR mediated by the Akt/PKB substrate PRAS40[J].Nat Cell Biol,2007,9(3):316- 23.
[25]Wang S Y,Yu Q J,Zhang R D,Liu B.Core signaling pathways of survival/death in autophagy-related cancer networks[J].Int J Biochem Cell Biol,2011,43:1263-6.
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07
波谱学杂志(2022年1期)2022-03-15
天津医科大学学报(2021年4期)2021-08-21
现代临床医学(2021年2期)2021-03-29
汽车维修与保养(2021年8期)2021-02-16
天津医科大学学报(2019年6期)2019-08-13
天然产物研究与开发(2018年7期)2018-08-21
分析化学(2017年12期)2017-12-25
安徽医科大学学报(2015年9期)2015-12-16
安徽医科大学学报(2015年9期)2015-12-16
