
一类具有常数避难所与收获率的捕食-食饵模型的稳定性分析
2012-09-18 02:20:12周稻祥朱长荣
重庆理工大学学报(自然科学) 2012年12期
周稻祥,朱长荣
(重庆大学数学与统计学院,重庆 401331)
捕食-食饵模型一直都是生物数学研究的重要领域。近年来,由经典的捕食-食饵模型衍生出很多带各种新参数的种群模型,因为这些模型更能反映现实世界的真实状况,因此,引起了人们的广泛关注[1-119]。在这些模型中,带食饵避难所的捕食-食饵模型是重要的一种。研究结果指出:避难所在捕食者和食饵相互作用中发挥着稳定性的作用,并且避难所参数对系统的动力行为发挥着重要影响[2-9]。Olivares 和 Jiliberto[2]提出了如下的带食饵避难率的捕食者-食饵的种群模型:
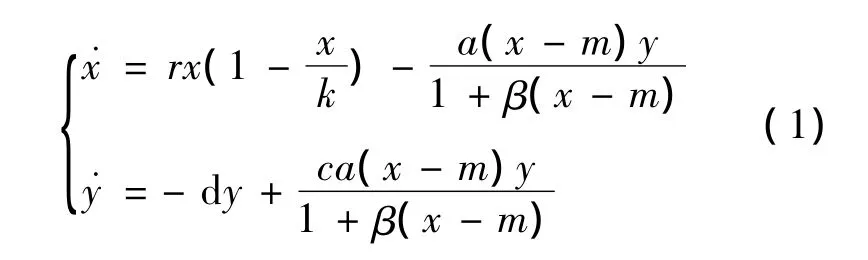
其中:所有参数均非负;x和y分别表示在时间t的食饵和捕食者的种群密度;r表示食饵的内禀增长率;k表示最大环境容量;c表示转化率;a(xm)y/(1+β(x-m))表示HollingⅡ功能反应函数;d是捕食者的死亡率;m表示食饵避难所。
系统(1)已有很多研究结果[2,6-7]。文献[2]利用微分方程定性理论的方法指出系统(11)有3个平衡点,内部平衡点是局部渐近稳定的,而且证明了极限环的存在性。文献[6]通过构造Dulac函数研究了系统(1)的全局稳定性,且用Poincare-Bendixson定理证明了极限环的唯一性。
在生物数学中,考虑到生物链的健康性、可持续性以及生物链产生的效益,近期有很多学者研究了带收获率的生物模型[13-14]。朱天晓[13]考虑了具有线性收获率的生物模型:
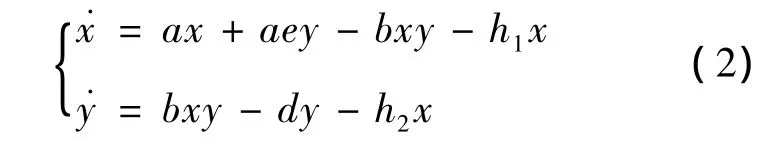
其中h1、h2为捕获强度。文献[13]利用微分方程定性理论,给出了系统(2)的平衡点的稳定性条件,通过构造Dulac函数的方法,利用Bendixson-Dulac判定定理,证明了系统(2)不存在极限环。
在模型(1)和(2)的基础上,本文研究带线性功能反应曲线、常数食饵避难所和线性收获率的捕食-食饵模型。具体来说,本文考虑以下的捕食-食饵模型:
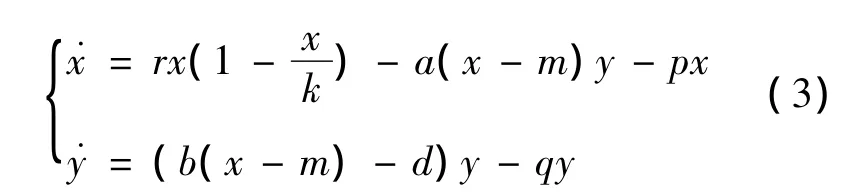
其中:x、y、r、k、d 的生物意义和模型(1)一样;(x -m)y表示线性功能反应函数;a、b为捕食系数;px、qy表示线性收获率。
本文讨论了系统(3)的全局与局部动力性态。利用Gronwall不等式,证明了系统(3)的解的一致有界性。利用平面系统的动力性质理论,研究了系统(3)的平衡点的类型及稳定性。分析发现:系统在第1象限及其边界上至多有3个平衡点,其中2个为边界平衡点,1个为内平衡点。边界平衡点可以为双曲鞍点、双曲结点,也可以为非双曲的鞍-结点;内平衡点的动力性态较丰富,对不同的参数取值范围,可以是稳定结点、稳定焦点。证明了内平衡点的全局稳定性。最后进行了数值模拟,模拟结果与理论结果完全吻合。
1 系统解的一致有界性
因为生物系统中的捕食者与食饵的数量都非负,而且系统(3)带有食饵避难率,因此x>m。实际上,系统(3)在 Ω ={(x,y)|x>m,y >0}中是一致有界的。
定理1 系统(3)在Ω中的解一致有界。
证明 令W=x+ay/b。对方程两边关于时间求导,并将式(3)代入,可得

上式右边是一个开口向下的二次函数,当 x=k(β +r-p)/2r时+βW 达到最大值:

上式中令 t→∞,有0<W <μ/β,即系统(3)的解是一致有界的。
定理1表明,系统(3)中捕食者和食饵的初始种群密度在Ω中,捕食者和食饵的种群密度都降落在Ω的一个有界集内。这表明捕食者和食饵可以在Ω中长期共存。
2 平衡点的动力性态
本节将研究系统(3)的动力性态。很显然,x轴的正半轴是系统(3)的不变流形,y轴的正半轴不是系统(3)的不变流形,但是在y轴的正半轴上点的向量场是(+,-),所以经过y轴正半轴的轨线,都要从第2象限进入到第1象限。因而,第1象限及其边界是系统(3)的不变集。下面将在第1象限及其边界上讨论系统(3)的动力性态。
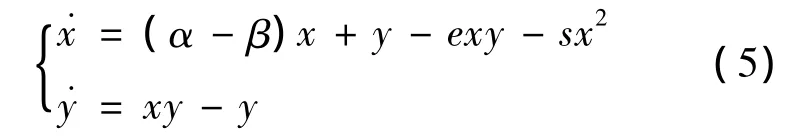
其中 α =r/ρ,β =p/ρ,e= ρ/bm >1,s=r/bk。
平衡点在讨论系统的动态行为时起着十分重要的作用,对平衡点的动力行为的研究关系着系统的局部稳定性、全局稳定性等。分2种情况计算系统(5)的平衡点。
1)当y=0时,系统有2个平衡点:A=(0,0)和B=((α -β)/s,0)。注意到 α=β时,A与B 重合,并且捕食者与食饵均非负,因此B存在,当且仅当α>β。所以仅在α>β的条件下讨论系统(5)在B处的动力性态。
2)当y≠0时,系统有平衡点C=(x*,y*)=1,(α-β-s)/(e-1())。注意到x*>0,y*>0,且e>1,所以C在第1象限内,当且仅当α>β+s。因此仅在α>β+s的条件下,讨论系统(5)在C处的动力性态。
在讨论平衡点的动力性态时,平衡点处的雅可比矩阵起着重要作用。令(x0,y0)是系统(5)的平衡点,则系统(5)在平衡点处的雅可比矩阵为
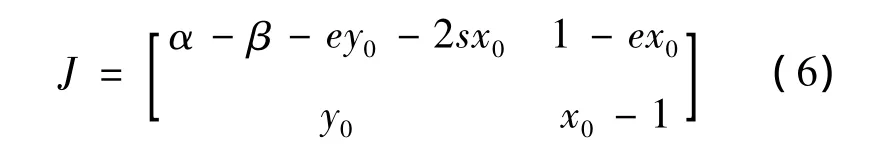
首先讨论平衡点A处的动力性态。
定理2 1)当 e>1,s>0,α >β>0时,A为不稳定鞍点;2)当 e>1,s>0,0<α<β时,A为稳定结点;3)当 e>1,s>0,α=β>0时,A为鞍 -结点。
证明 系统(5)在平衡点A处的雅可比矩阵为

所以特征值为:λ(A)1= -1<0,λ(A)2=α-β。
1)当e>1,s>0,α >β>0时,有 λ(A)2>0,则A为不稳定鞍点。
2)当e>1,s>0,0<α <β时,有 λ(A)2<0,则A为稳定结点。
3)当e>1,s>0,α=β>0时,有,在平衡点 A处作变换 x=u-v,y=v,则系统(5)的变为
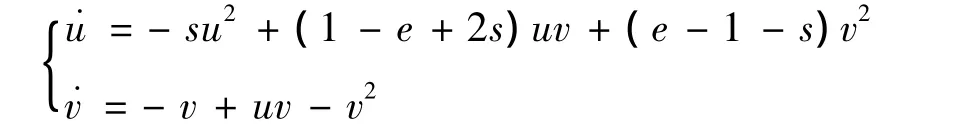
依照文献[8]中2.11的定理1知A是鞍-结点。
因为平衡点B存在,当且仅当α>β,在α>β的条件下考虑B点处的动力性态。
定理3 1)当 e>1,s>0,α >s+β 时,B 为不稳定鞍点;2)当 e>1,s>0,β<α <β+s时,B 为稳定结点;3)当 e>1,s>0,α=β+s时,B 为鞍 -结点。
证明 系统(5)在平衡点B处的雅可比矩阵为

所以特征值为:λ(B)1= -(α -β)<0,λ(B)2=(α - β)/s-1。
1)当e>1,s>0,α >s+β 时,有 λ(B)2>0,则B为不稳定鞍点。
2)当e>1,s>0,β <α <β+s时,有 λ(B)2<0,则B为稳定结点。
3)当 e>1,s>0,α =β +s时,λ(B)2=0,且平衡点B=(1,0)。将平衡点 B移到原点,令 x=u+1,y=v,则系统化为
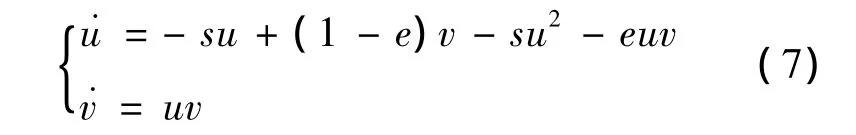
对系统(7)引入变换 u=x+(1-e)y,v=sy,则系统(7)变为
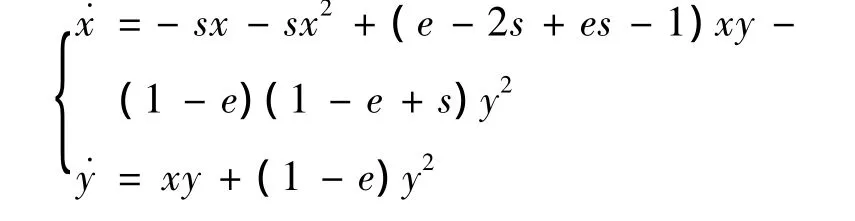
依照文献[8]中2.11的定理1知B是鞍-结点。
最后,因为C在第1象限内,当且仅当α>β+s,所以在α>β+s的条件下讨论系统(5)在C处的动力性态。
定理4 1)当 s>0,β+s<α≤β+s+s2/4,e>1时,C 为稳定结点;2)当 s>0,β+s+s2/4<α时,令e0=(α-β-s)/(2-s)>0,则:① 如果1<e≤e0+1,则C为稳定结点;② 如果e>e0+1,则C是稳定焦点。
证明 系统(5)在平衡点C处的雅可比矩阵为
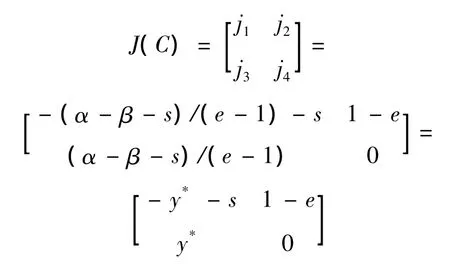
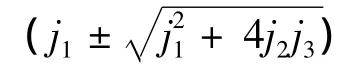

1)当 s>0,β+s<α≤β+s+s2/4,e>1时,+4j2j3≥0,λ(C)1、2为负实数,故 C 为稳定结点。
2)当s>0,β+s+s2/4<α时,注意到ξ>0且+4j2j3和 g(ξ)的符号相同,有下面的结论:① g(ξ)≥0,当且仅当0 <ξ≤ξ2,即1 <e≤e0+1,则λ(C)1、2为负实数,故 C 为稳定结点;② g(ξ)<0,当且仅当 ξ>ξ2,即e>e0+1,则λ(C)1、2为具有负实部的复数,故C是稳定焦点。
从定理4看到,内部平衡点C总是局部渐近稳定的。实际上,还能得到C点的全局稳定性:当s>0,α >β+s,e>1时,平衡点 C 是全局稳定的。这是因为:将平衡点C移到原点,令x=u+x*,y=v+y*,则系统(5)化为

由文献[4]中定理10.6知平衡点 C是全局稳定的。
3 数值模拟
现在通过数值模拟来验证理论分析结果。为了模拟此系统,令 α =0.469,β =0.352,e=2.13,s=0.0166,得到系统在平衡点C(1,0.088849)处是渐近稳定的。在初值为(2,0.3)时得到的相图见图1。捕食和食饵随时间的变化见图2。捕食者和食饵随时间的变化见图3、4。

图1 系统(5)在平衡点C(1,0.088849)处的相图
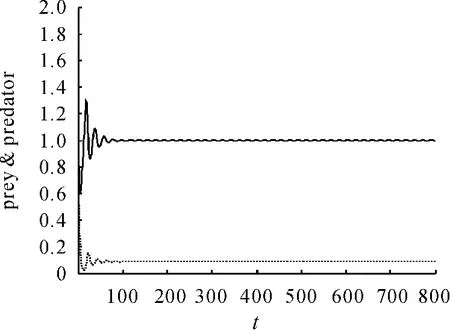
图2 捕食和食饵随时间的变化
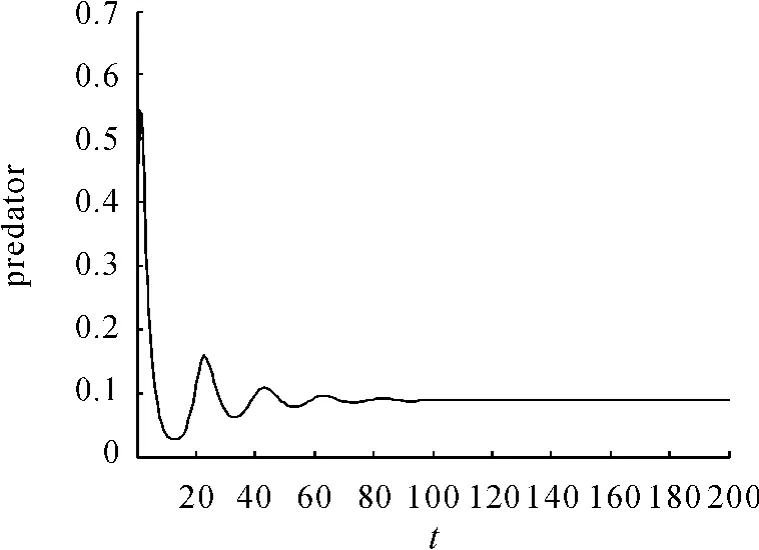
图3 捕食随时间的变化
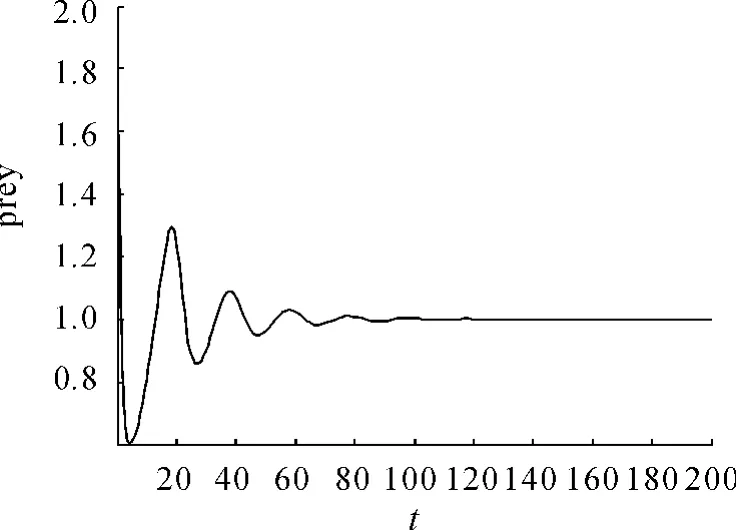
图4 食随时间的变化
4 结束语
本文进行了系统解的一致有界性以及平衡点稳定性的定性分析。分析指出增加避难所的数量就可以增加食饵的密度,且稳定性和动态行为受到避难所参数m的影响,内部平衡点C是全局稳定的,即无论开始捕食和食饵的数量是多少,最终将要稳定在一定的数量之内。数值模拟也很好地印证了这些结果。
[1]Birkoff G,Rota G C.Ordinary differential equations[M].Ginn:[s.n.],1982.
[2]González-Olivares E,Ramos-Jiliberto R.Dynamic consequences of prey refuges in a simple model system:More prey,fewer predators and enhanced stability[J].Ecological Modeling,2003,166:135 -146.
[3]Dai G,Tang M.Coexistence region and global dynamics of a harvested predator-prey system[J].SIAM Journal on Applied Mathematics,1998(1):193 -210.
[4]Grank J,Martin H G,Melluish D M.Nonlinear ordinary differential equations[M].USA:[s.n.],1977.
[5]Yuri K.Elements of Applied Bifurcation Theory[M].Second Edition.New York:Springer,1998:79 -100.
[6]Chen L,Chen F.Qualitative analysis of a predator prey model with Holling type II functional response incorporating a constant prey refuge[J].Nonlinear Analysis Real World Applications,2008(10):125 -127.
[7]Ji L L,Wu C Q.Qualitative analysis of a predator-prey model with constant-rate prey harvesting incorporating a constant prey refuge[J].Nonlinear Analysis Real World Applications,2010(11):2285 -2295.
[8]Perko L.Differential Equation and Dynamical Systems[M].Sencond Edition.New York:Spinger,1996:146-152.
[9]Kar T K.Modeling and analysis of a harvested prey-predator system incorporating a prey refuge[J].Journal of Computational and Applied Mathematics,2006,185:19-33.
[10]Kar T K.Stability analysis of a prey-predator model incorporating a prey refuge[J].Communications in Nonlinear Science and Numerical Simulation,2005(10):681-691.
[11]Tao Y D,Wang X,Song X Y.Effect of prey refuge on a harvested predator-prey model with generalized functional response[J].Commun Nonlinear Sci Numer Simulat,2011,16:1052 -1059.
[12]张芷芬,丁同仁,黄文灶,等.微分方程定性理论[M].北京:科学出版社,1985:49-90.
[13]朱天晓.具有线性收获率的Diekman模型稳定性[J].长春大学学报,2007(3):1-4.
[14]刘娟,李医民.两种群都有收获率的微分生态系统的定性分析[J].徐州师范大学学报,2011(2):32-35.
[15]赵琳,雒志学,王利红.在容量较小的污染环境中种群的最优捕获问题[J].重庆理工大学学报:自然科学版,2011,25(2):122 -126.
[16]张靖.食饵具有流行病的捕食-被捕食(SIS)模型的分析[J].四川兵工学报,2011(9):153-156.
[17]张志军,刘启宽,李映辉.具有线性收获率且包含食饵避难所的一类食饵-捕食者模型的定性分析[J].重庆理工大学学报:自然科学版,2011,25(9):104-113.
[18]李盈科,樊小琳,陈亮.带有非线性传染率的阶段结构捕食食饵模型的持久性[J].四川兵工学报,2011(7):151-154.
[19]刘启宽,罗廷友,李映辉.一类具功能反应且常数投放的食饵-捕食系统的定性分析[J].重庆理工大学学报:自然科学版,2011,25(3):1118 -122.
猜你喜欢
数学物理学报(2022年1期)2022-03-16 06:15:06
数学物理学报(2021年5期)2021-11-19 07:01:24
科学咨询(2021年25期)2021-09-14 01:39:04
云南大学学报(自然科学版)(2021年1期)2021-02-05 08:04:04
应用数学(2020年4期)2020-12-28 00:37:02
数学物理学报(2020年4期)2020-09-07 09:14:24
数学物理学报(2019年4期)2019-10-10 02:40:04
数学物理学报(2019年4期)2019-10-10 02:39:24
数学物理学报(2019年3期)2019-07-23 01:15:30
太原师范学院学报(自然科学版)(2018年2期)2018-08-17 12:21:50
