
EB病毒核抗原3C提高Gemin3基因的表达
2012-09-07 09:15郭毅孔繁明赵楠
中国医科大学学报 2012年6期
郭毅,孔繁明,赵楠
(1.中国医科大学附属第一医院妇科,沈阳 110001;2.辽宁省宽甸县中心医院妇产科,辽宁 宽甸 118200)
EB病毒是第1个被发现的人类肿瘤病毒,与多种淋巴细胞增殖性疾病的发生相关。在体外,EB病毒可将静止的B细胞转化为具有无限增殖能力的潜伏感染的B细胞,即淋巴母细胞样B细胞系。此过程的关键调控因子包括EB病毒核抗原(Epstein-Barr nuclear antigen,EBNA)2、EBNA3A、EBNA3C 和LMP1 蛋白[1,2]。EBNA3C 作为转录调节因子能够与多种细胞及病毒因子相互作用。Gemin3是EBNA2、EBNA3C 及运动神经元生存因子的结合蛋白[3,4],属于DExD/H框RNA解旋酶家族,在RNA代谢过程中起重要作用[5,6]。Gemin3还可结合并调控多种细胞转录因子[7~11]。我们的前期研究结果显示:Gemin3可抑制p53介导的转录和凋亡途径,提示肿瘤病毒可能通过调控Gemin3靶基因的表达而抑制宿主细胞的凋亡。因此,本研究拟通过慢病毒载体介导发卡RNA干涉敲减EBNA3C基因的表达,探讨EBNA3C与Gemin3的相互作用。
1 材料与方法
1.1 材料
pA3F-EBNA3C质粒表达EBNA3C全长,羧基端由Flag标记。HEK293为经腺病毒DNA转化的人胚胎肾细胞系,LCL1和LCL2为体外转化的EBV阳性细胞系,表达EBNA3C的细胞系(BJAB7 and BJAB10)由pZipneo真核细胞表达载体介导转染EBNA3CcDNA,并经过新霉素选择建立,以上质粒及细胞系均由宾夕法尼亚大学医学院Erle Robertson教授馈赠。鼠抗Gemin3单抗由宾夕法尼亚大学医学院Gideon Dreyfuss惠赠,鼠抗Flag单克隆抗体M2购自Sigma-Aldrich公司,鼠抗EBNA3C单抗A10来自杂交瘤。
1.2 方法
1.2.1 细胞培养:HEK 293细胞采用 DMEM(Hy-Clone公司)培养基培养,BJAB,RAMOS和EBV阳性的细胞系采用RPMI1640培养基培养,分别加入10%胎牛血清,50 U/mL青霉素,50 μg/mL链霉素和2 mmol/L L-谷氨酰胺。在37℃、5%CO2、饱和湿度环境下培养。
1.2.2 转染:采用Bio-Rad Gene PulserⅡ型电穿孔仪瞬时转染细胞。收集对数生长期细胞(1.5×107个),PBS冲洗,悬浮于 400 μL已加入待转染EBNA3C质粒的培养基中,转入0.4 cm间距电转杯中,电容为 975 μF,转染 HEK 293和 SAOS-2电压为210 V,B淋巴细胞为220 V。将转染的细胞移至加入10 mL培养基的培养皿或培养瓶中,37℃培养36 h后收集细胞。
1.2.3 Western blot:收集细胞,PBS洗涤,0.5 mL冰冷RIPA缓冲液裂解,加入蛋白酶抑制剂,21 000g(10 min、4℃)离心,上清转移至新的微小离心管。用Laemmli上样缓冲液加热变性蛋白裂解物和免疫沉淀复合物,SDS聚丙烯酰胺凝胶电泳分离,转膜。用相关抗体检测醋酸纤维膜中的蛋白,再与相应的发射红外光的第2抗体共培育后,用Odyssey imager(LiCor Inc.,Lincoln,NE)扫描。
1.2.4 慢病毒shRNA载体的构建:将EBNA3C shRNA(CCATATACCGCAAGGAATA)插入 pGIPZ载体,按说明书(Open Biosystem公司)进行操作,表达EBNA3C小发卡状RNA的质粒缩写为sh-E3C。此外,选取与已知人mRNA无同源性的21个寡核苷酸序列插入相同载体作为对照,缩写为sh-C。
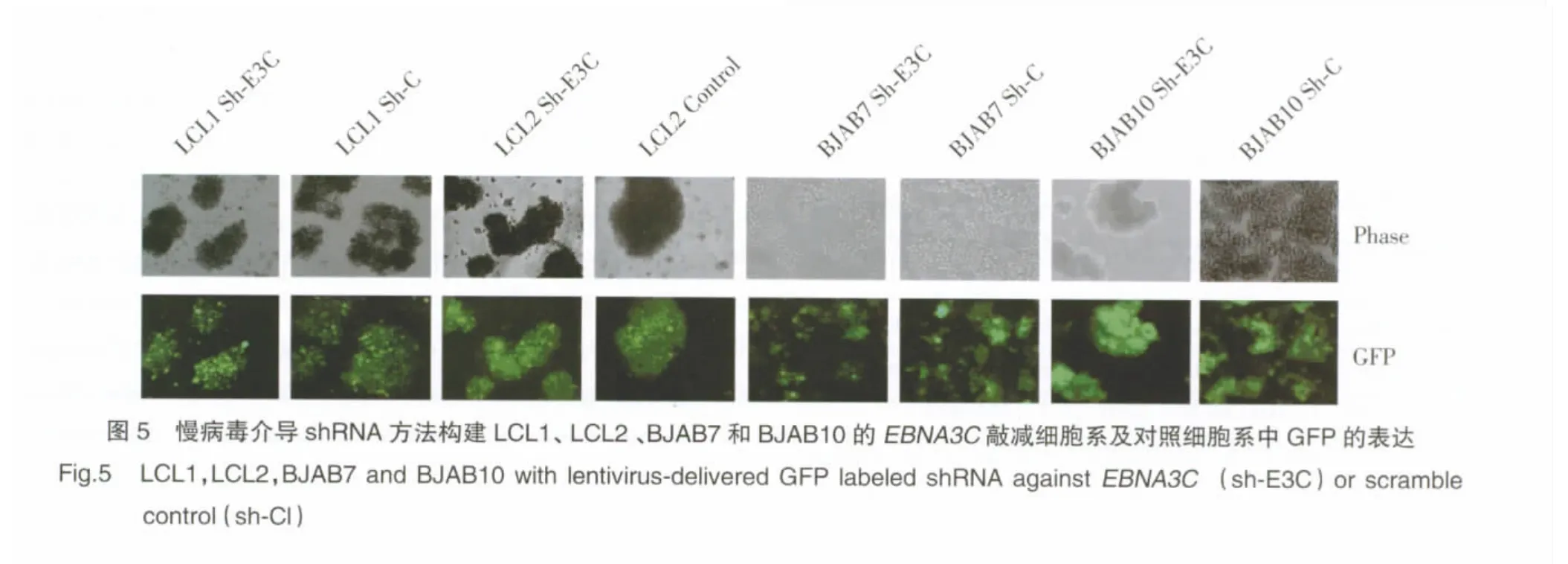
1.2.5 病毒的产生及感染B细胞:通过瞬时转染HEK 293T细胞产生慢病毒。将HEK 293T细胞(2×106个)培养于直径10 cm的培养皿中,采用添加10%FBS和1%抗菌防霉溶液的DMEM培养基,5%CO2培养24 h后转染。每个培养皿中加入需转染的20 μg DNA 质粒(包括 1.5 μg包膜质粒 pCMV-VSVG,3 μg包装结构质粒 pRSV-REV,5 μg包装结构质粒 pMDLg/Prre,10.5 μg的慢病毒载体质粒)。采用磷酸钙沉淀法转染HEK 293T细胞:上述质粒加蒸馏水共 438 μL 、2 mol/L Cacl262 μL 以及 2×HEPES 缓冲盐水液500 μL,室温放置30 min;转染前5 min加入氯喹使终浓度为25 μmol/L;12 h后用含有10%FBS、10 mmol/L HEPES及 10 mmol/L丁酸钠的DMEM更换培养液,10 h后用含有10%FBS和10 mmol/L HEPES的DMEM再次更换培养基。每隔12 h收集1次调整培养基,共4次,用0.45 μm孔径醋酸纤维素膜过滤后冻存,最后以70 000g离心2.5 h收集病毒。用RPMI重新悬浮病毒,加入20 μmol/mL聚凝胺后,感染细胞(1×106)。72 h后加入嘌呤霉素使终浓度达到2 μg/mL以筛选感染的细胞。采用Olympus IX71荧光显微镜在560 nm激发并645 nm发射滤过条件下检测绿色荧光蛋白。在2 μg/mL嘌呤霉素条件下,见细胞团饱和度达到80%时,收集细胞做Western blot检测。
2 结果
2.1 EBNA3C转染导致Gemin3表达增加
为了研究EBNA3C或EBV对Gemin3蛋白表达的影响,我们对Gemin3蛋白在不同细胞中的表达做了定量分析,结果显示:与对照细胞相比,EBNA3C和EBV阳性细胞系中Gemin3蛋白表达量明显增高(图 1,图 2)。

为确定Gemin3表达的增加是由于EBNA3C作用的结果,我们用EBNA3C质粒转染HEK 293细胞并检测Gemin3蛋白量,发现Gemin3蛋白表达水平随着EBNA3C转染量的增加而升高(图3)。

我们通过慢病毒介导shRNA敲减EBNA3C及EBV阳性细胞内EBNA3C基因的表达,经过筛选获明显降低(图4)。用Olympus IX71荧光显微镜观察细胞中绿色荧光蛋白,绿色荧光蛋白为慢病毒转染成功的指示蛋白,结果与Western blot一致(图5)。得稳定的携带sh-EBNA3C及对照的细胞系,采用Western blot检测并比较EBNA3C敲减细胞与对照细胞中Gemin3蛋白的表达水平。结果显示:与对照细胞相比,EBNA3C敲减细胞中Gemin3表达水平

3 讨论
EBNA3C蛋白由992个氨基酸组成,呈颗粒状散在于细胞核内,并具有多种功能:与RBP-Jκ形成稳定的复合物并抑制后者与DNA结合,具有转录调节作用[12,13];与活化的(Ha-)ras基因共同作用转化大鼠的胚胎纤维母细胞,具有癌基因的功能[14];通过蛋白之间直接相互作用调节细胞周期,通过召集泛素化连接酶E3复合物机制降解Rb或p27蛋白[15,16];稳定宿主细胞内的癌蛋白,如c-Myc和Mdm2[17,18]。
Gemin3最初是在寻找与EBNA2及EBNA3C相互作用的细胞因子的过程中通过酵母双杂交实验发现的[4]。无论是对于果蝇还是哺乳动物,Gemin3都是胚胎发育和成活的关键基因[19,20]。Gemin3 通过与其他分子作用而表现出不同的生物学功能:与类固醇生长因子1(SF-1)结合并在转录水平抑制后者的功能[7];与有丝分裂元Ets转录抑制因子METS/PE1或ERF形成复合物,通过召集组蛋白乙酰基转移酶控制细胞的增殖和分化[11,21];通过与早期生长反应因子2结合并部分依赖召集HDAC而抑制其转录活性[9];通过与叉头转录因子2结合诱导凋亡[10]。Gemin3与肿瘤相关的直接证据来源于膀胱癌危险因素与microRNA相关基因突变关系的临床分析,Gemin3基因的非同义单核苷酸多态性的同源性变异是膀胱癌发病的高危因素[22]。Gemin3具有抑制p53基因转录的功能,慢病毒介导shRNA敲减Gemin3并经过筛选的细胞经流式细胞检测显示凋亡增加[23]。
本研究通过共转染EBNA3C和Gemin3基因至HEK293细胞,依靠慢病毒载体介导发卡RNA干涉敲减EBNA3C基因的表达,并经嘌呤霉素筛选获得稳定的EBNA3C低表达细胞系,采用Western blot及荧光显微镜检测了EBNA3C对Gemin3蛋白表达的影响。结果表明EBNA3C能够上调具有癌蛋白功能的Gemin3表达水平。今后我们将在转录及蛋白修饰水平对其可能的调控机制开展进一步的研究。
[1]Rickinson AB,Kieff E.Epstein-Barr virus [M].Philadelphia:Lippincott Williams& Wilkins,2002:2575-2627.
[2]Zhao B,Sample CE.Epstein-barr virus nuclear antigen 3C activates the latent membrane protein 1 promoter in the presence of Epstein-Barr virus nuclear antigen 2 through sequences encompassing an spi-1/Spi-B binding site[J].J Virol,2000,74(11):5151-5160.
[3] Charroux B,Pellizzoni L,Perkinson RA,et al.Gemin3:a novel DEAD box protein that interacts with SMN,the spinal muscular atrophy gene product,and is a component of gems[J].J Cell Biol,1999,147(6):1181-1194.
[4]Grundhoff AT,Remmer EK,Tureci O,et al.Characterization of DP103,a novel DEAD box protein that binds to the Epstein-Barr virus nuclear proteins EBNA2 and EBNA3C[J].J Biol Chem,1999,274(27):19136-19144.
[5]Cordin O,Banroques J,Tanner NK,et al.The DEAD-box protein family of RNA helicases[J].Gene,2006,367(15):17-37.
[6]Rocak S,Linder P.DEAD-box proteins:the driving forces behind RNA metabolism[J].Nat Rev Mol Cell Biol,2004,5(3)P:232-241.
[7]Ou Q,Mouillet JF,Yan X,et al.The DEAD box protein DP103 is a regulator of steroidogenic factor-1[J].Mol Endocrinol,2001,15(1):69-79.
[8]Yan X,Mouillet JF,Ou Q,et al.A novel domain within the DEAD-box protein DP103 is essential for transcriptional repression and helicase activity[J].Mol Cell Biol,2003,23(1):414-423.
[9]Gillian AL,Svaren J.The Ddx20/DP103 dead box protein represses transcriptional activation by Egr2/Krox-20[J].J Biol Chem,2004,279(10):9056-9063.
[10]Lee K,Pisarska MD,Ko JJ,et al.Transcriptional factor FOXL2 interacts with DP103 and induces apoptosis [J].Biochem Biophys Res Commun,2005,336(3):876-881.
[11]Klappacher GW,Lunyak VV,Sykes DB,et al.An induced Ets repressor complex regulates growth arrest during terminal macrophage differentiation[J].Cell,2002,109(2):169-180.
[12]Robertson ES,Grossman S,Johannsen E,et al.Epstein-Barr virus nuclear protein 3C modulates transcription through interaction with the sequence-specific DNA-binding protein J kappa [J].J Virol,1995,69(5):3108-3116.
[13]Marshall D,Sample C.Epstein-Barr virus nuclear antigen 3C is a transcriptional regulator[J].J Virol,1995,69(6):3624-3630.
[14]Parker GA,Crook T,Bain M,et al.Epstein-Barr virus nuclear antigen (EBNA)3C is an immortalizing oncoprotein with similar properties to adenovirus E1A and papillomavirus E7[J].Oncogene,1996,13(12):2541-2549.
[15]Knight JS,Sharma N,Robertson ES.Epstein-Barr virus latent antigen 3C can mediate the degradation of the retinoblastoma protein through an SCF cellular ubiquitin ligase[J].Proc Natl Acad Sci USA,2005,102(52):18562-18566.
[16]Knight JS,Sharma N,Robertson ES.SCFSkp2 complex targeted by Epstein-Barrvirusessentialnuclearantigen[J].MolCellBiol,2005,25(5):1749-1763.
[17]Bajaj BG,Murakami M,Cai Q.Epstein-Barr virus nuclear antigen 3Cinteracts with and enhances the stability of the c-Myc oncoprotein[J].J Virol,2008,82(8):4082-4090.
[18]Saha A,Murakami M,Kumar P.Epstein-Barr virus nuclear antigen 3C augments Mdm2-mediated p53 ubiquitination and degradation by deubiquitinating Mdm2[J].J Virol,2009,83(9):4652-4669.
[19]Cauchi RJ,Davies KE,Liu JL.A motor function for the DEAD-box RNA helicase,Gemin3,in Drosophila [J].PLoS Genet,2008,4:e1000265.
[20]Mouillet JF,Yan X,Ou Q,et al.DEAD-box protein-103(DP103,Ddx20)is essential for early embryonic development and modulates ovarian morphology and function[J].Endocrinology,2008,149(5):2168-2175.
[21]Hester KD,Verhelle D,Escoubet-Lozach L,et al.Differential repression of c-myc and cdc2 gene expression by ERF and PE-1/METS[J].Cell Cycle,2007,6(13):1594-1604.
[22]Yang H,Dinney CP,Ye Y,et al.Evaluation of genetic variants in microRNA-related genes and risk of bladder cancer[J].Cancer Res,2008,68(7):2530-2537.
[23]郭毅,张晓光,赵楠,等.Gemin3基因通过抑制p53表达阻碍细胞凋亡[J].中华肿瘤杂志,2011,33(11):810-815.
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
华人时刊(2022年9期)2022-09-06
成都医学院学报(2022年4期)2022-08-19
江西农业学报(2021年4期)2021-04-20
华人时刊(2020年15期)2020-12-14
三农资讯半月报(2020年11期)2020-06-21
医学研究杂志(2015年9期)2015-07-01
癌变·畸变·突变(2015年4期)2015-02-27
中华胰腺病杂志(2014年1期)2014-08-04
中华介入放射学电子杂志(2014年1期)2014-02-02
