
马耳他布鲁菌omp25的原核表达与免疫原性检测
2012-08-08 06:13吴树清李东兴刘艳琴张明月
中国兽医杂志 2012年2期
吴 杰,吴树清,李东兴,刘艳琴,张明月,海 岩
(1.内蒙古农业大学兽医学院,内蒙古 呼和浩特 010018;2.内蒙古疾病预防控制中心,内蒙古 呼和浩特 010018)
布鲁菌病是由布鲁菌属细菌引起的动物源性疾病,是一种人畜共患传染性疾病,该病在我国发病呈逐年上升趋势[1]。它主要引起人类波状热和慢性感染以及反刍动物流产和睾丸炎等,目前尚无根治方法。大量研究表明,布鲁菌的外膜蛋白具有很强的免疫原性[2-3]。omp25是布鲁菌ompA家族成员之一,omp25基因在布鲁菌各个种属之间有很高的保守性[4]。本试验克隆了马耳他布鲁菌omp25基因,在大肠埃希菌Rosetta中表达,用纯化的表达产物进行Western-blot分析,发现重组蛋白具有免疫原性。为检测布鲁菌病及研制布鲁菌亚单位疫苗提供了科学依据。
1 材料与方法
1.1 菌株和质粒 马耳他布鲁菌为内蒙古农业大学微生物实验室分离株。pET-32a、Rosetta购自宝生物工程(大连)有限公司。
1.2 试剂 T4连接酶,DNA Marker、蛋白Marker、IPTG和限制性内切酶Eco RⅠ、Xho lⅠ购自宝生物工程(大连)有限公司;辣根过氧化物酶(HRP)标记兔抗羊IgG,购自Sigma公司。引物由上海生工生物工程技术服务有限公司合成,浓度2OD/管,储存液浓度100μmol/L。羊种布鲁菌阳性血清和标准阴性血清由本实验室保存,其他常规试剂均为国产试剂。
1.3 目的片段的扩增 根据GenBank上发表的M16布鲁菌的omp25的基因序列设计马耳他布鲁菌上下游引物P1:5′cgcctcgagttagaacttgtagt 3′,P2:5′ccgggatccatgaaatccgtaat 3′(分别加入Eco RⅠ和Xho lⅠ酶切位点)。PCR反应条件为:95℃预变性5 min,94℃变性40s,55℃复性45s,72℃延伸45s;30个循环,最后72℃延伸10min。同时设去离子水对照。反应结束后用1%琼脂糖凝胶电泳检测并观察结果。
1.4 omp25基因表达载体的构建 PCR产物经限制性内切酶Eco RⅠ、Xho lⅠ双酶切处理,1%琼脂糖凝胶电泳,回收预期的omp25基因片段与同样处理的pET-32a表达载体16℃连接过夜,转化至Rosetta中,涂LB平板(氨苄青霉素和氯霉素抗性),37℃培养过夜,挑取单克隆菌落37℃过夜摇菌,碱变性法提取质粒,进行PCR和Eco RⅠ、Xho lⅠ双酶切鉴定。挑取阳性单克隆,送上海生工生物工程技术服务有限公司测序,并与GenBank报道的omp25基因序列进行同源性比较。将重组质粒命名为pET-32a-omp25。
1.5 pET-32a-omp25重组蛋白的诱导表达 含重组质粒的菌液按1∶100的量接种在LB培养基中(含氨苄青霉素,氯霉素50μg/mL),37℃振荡培养至OD600值约为0.6~1.0,再加入IPTG使其终浓度达到1mmol/L。37℃继续培养,经12%分离胶检测表达情况。
1.6 表达产物的纯化和Western-blot检测 将上述的诱导表达样品以10000r/min离心收集菌体,沉淀物用原收集菌液量1/5体积的PBS悬浮,置冰浴中超声波处理10min,10000r/min(4℃)离心10 min,分别收集上清和沉淀。按照His GraviTrap和His GraviTrap Kit的说明书,对融合蛋白进行重力流纯化,对纯化产物进行SDS-PAGE分析。将纯化蛋白经SDS-PAGE后再转到NC膜上,5%脱脂奶粉4℃封闭过夜,PBST缓冲液洗3次,加入布鲁菌病阳性血清(1∶100倍稀释)37℃作用2h,PBST缓冲液洗3次,再加入兔抗羊IgG辣根过氧化物酶(HRP)标记抗体(1∶2000倍稀释),37℃作用1h,PBST缓冲液洗3次,DAB显色液中避光显色2~10min。
2 结果
2.1 pET-32a-omp25重组质粒的鉴定 pET-32aomp25重组质粒经PCR和Eco RⅠ、XhoⅠ双酶切鉴定,得到了预期大小的642bp片段。测序结果表明,目的基因omp25正向插入pET-32a载体(见图1)。
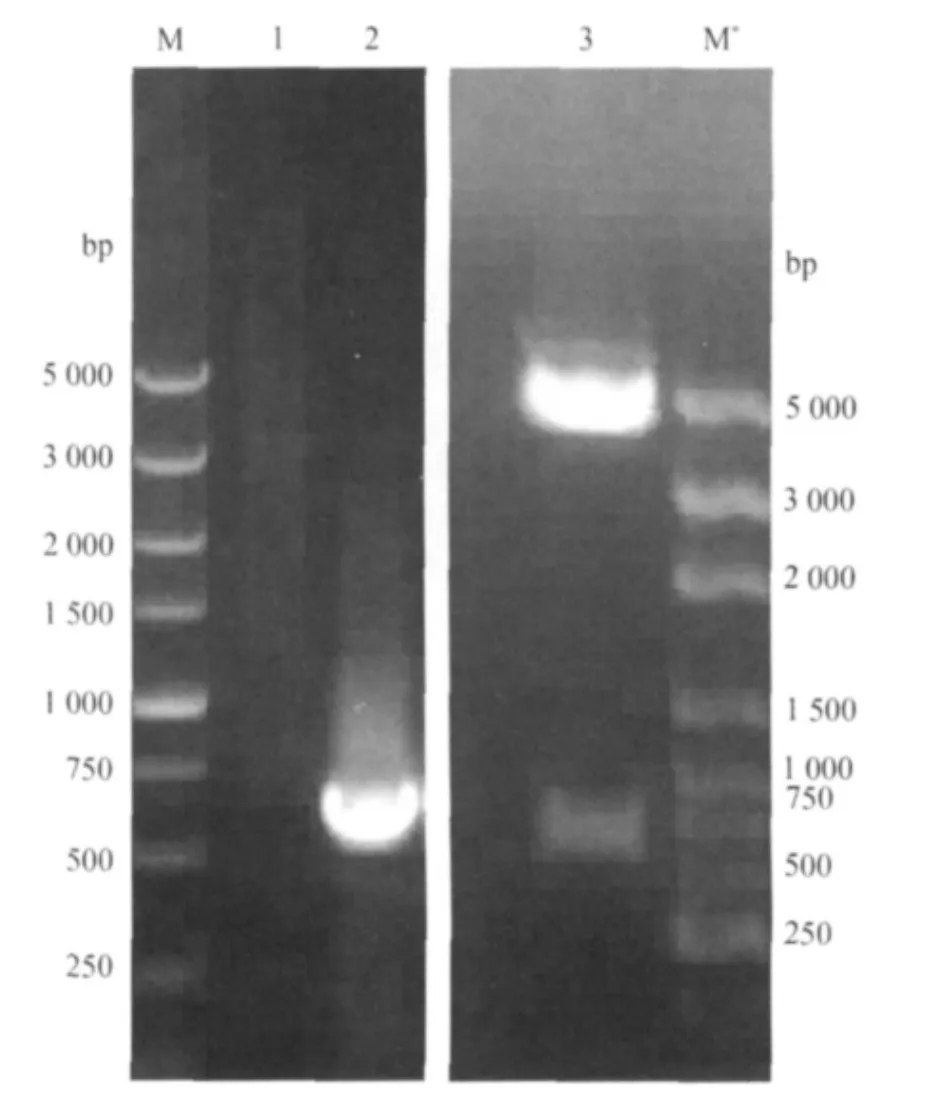
图1 重组质粒pET-32a-omp25的PCR及双酶切鉴定
2.2 表达产物的SDS-PAGE分析 SDS-PAGE分析结果表明,经1mmol/L的IPTG诱导的5h基因工程菌获得表达,产生约43kDa的特异性蛋白条带(见图2)。
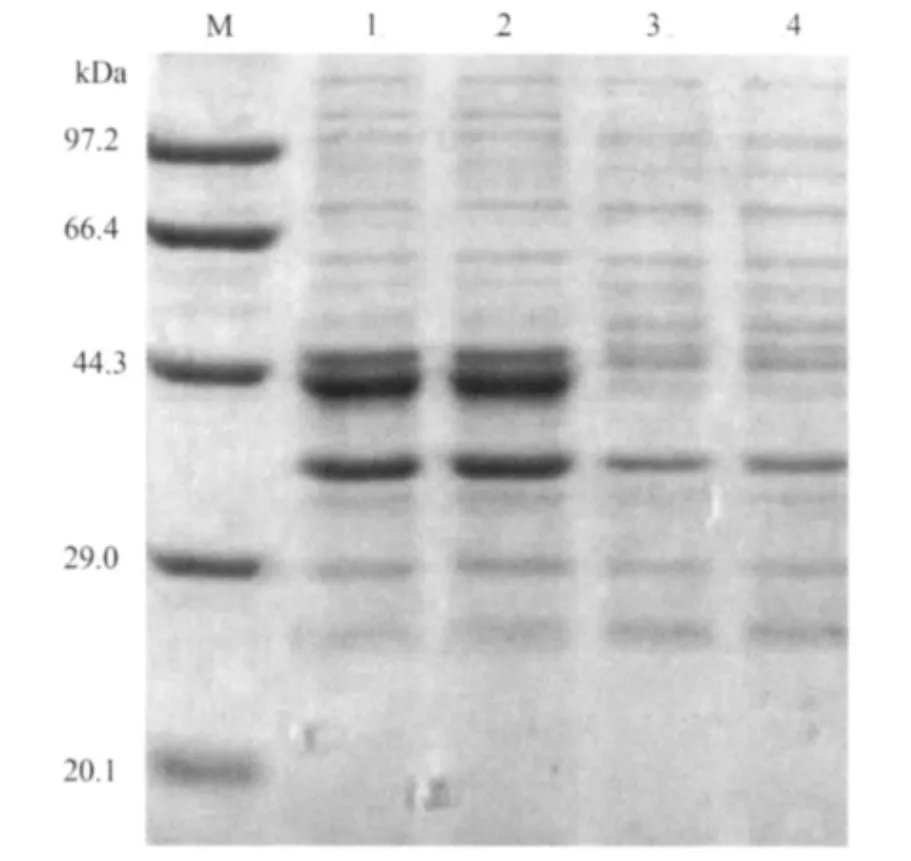
图2 表达产物的SDS-PAGE
2.3 表达产物的纯化及Western-blot检测结果纯化产物经SDS-PAGE电泳分析表明获得了高纯度的目的蛋白。Western-blot分析结果表明,表达的目的蛋白能与马耳他布鲁菌阳性血清发生特异性反应,43kDa处出现特异免疫反应条带(见图3)。
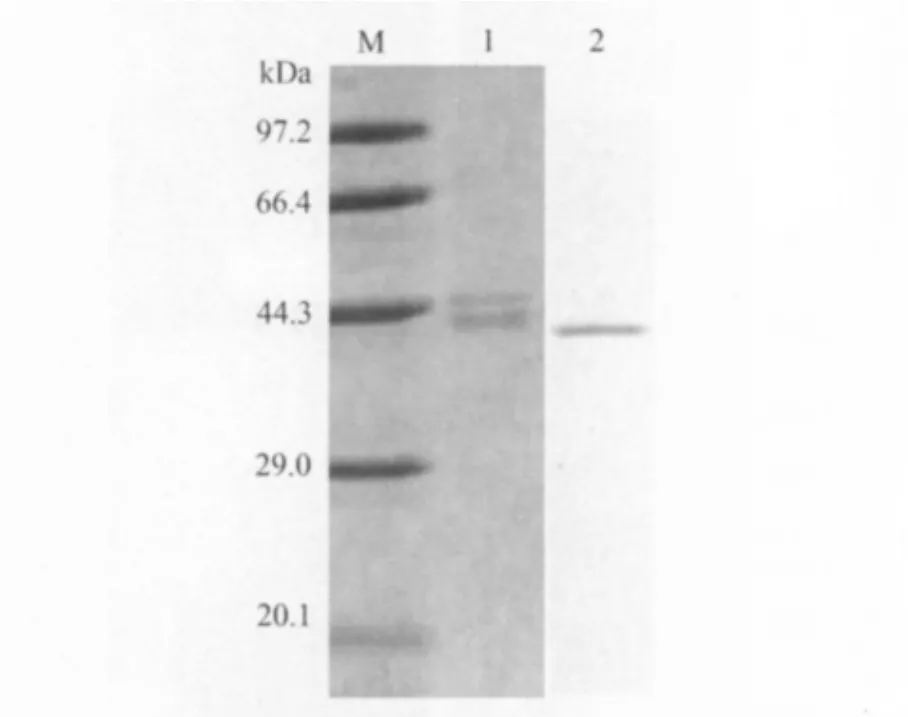
图3 表达产物纯化的SDS-PAGE及Western-blot分析
3 讨论
omp25是布鲁菌外膜结构蛋白[5],是具有代表性的第3组外膜蛋白[6]。在布鲁菌各个种属之间具有高度的保守性,同源性达98%以上。布鲁菌omp25基因的缺失可引起布鲁菌的毒力减弱,并能够对宿主起到免疫保护作用[7]。Philpper从B.abortus544基因库中,以抗omp25单克隆抗体调出了omp25基因,并分析了其脱氧核糖核苷酸序列omp25基因长约917bp,在起始密码子6个碱基前有7个核苷酸序列(TAAGGAG)与大肠杆菌16sRNA序列具有同源性,估计是核糖体结合位点。布鲁菌细胞壁膜结构是大多数细菌识别宿主的独特标志物,能绑定胞外的许多基质蛋白,特别是纤维连接素和玻璃体粘连蛋白,这也是许多其他的病原微生物普遍存在的一个属性。研究证实布鲁菌表面主要的抗原成分是LPS和omps。到目前为止,己发现有7种主要omps[8],它以共价键的形式与细胞外膜的肤聚糖(PG)层紧密结合。
本试验证实omp25蛋白表达量高,占总菌体蛋白的含量高,具有良好的免疫反应原性,具备作为免疫学活性抗原的潜力和优势,为疫苗的开发和免疫学检测方法的建立提供了候选抗原。
[1]文学忠,于瑞华,姜秋杰.布鲁氏菌病近况[J].吉林畜牧兽医,2007,5:20-23.
[2]Bowden R A,Cloeckaert A,Zygmunt M S,et al.Evaluation of immunogenicity and protective activity in BALB/c mice of the 25-kDa major outer-membrane protein of Brucella melitensis(OMP25)expressed in Escherichia coli[J].J Med Micobiol,1998,47:39-48.
[3]Brocker B J,Tabatabai L B,Mayfeld J E.Conserbation of antigenicityin a 31-kDa Brucella protein[J].Vet Microbiol,1998,18:313-525.
[4]Matthew D,Edmonds,Cloeckaert A,et al.Brucella species lacking the major outer membrane protein Omp25are attenuated in mice and protect against Brucella ovis[J].Vet Micobiol,2002,88:205-221.
[5]Bowden R A,Verger J M,Grayon M,et al.Rspid identification of rough Brucella isolates bu a latex coagglutination assay with the 25-lilodalton outer membrane protein and rough-lipopolysaccharide specific monoclonal antibodies[J].Clin Diagn Lsb Innunol,1997,4(5):611-614.
[6]Oliveira S C,Splitter G A.Immunization of mice with recombinant L7/L12ribosomal protein confers protein confers protection against Brucella abortus infection[J].Vaccine,1996,14(10):959-962.
[7]Edmonds M D,Cloeckaert A,Booth N J,et al.Attenuation of a Brucella abortus mutant lacking a major 25kDa outer membrane protein in cattl[J].Ameri J Vet Res,2001,62(9):1461-1466.
[8]Cloeckaert A,Zymunt M S,Wergifosse P,et al.Demonstration of pertidoglycan-associated Brucella outer-membrane proteins by use of monoclonal antibodies[J].J Gen Microbiol,1992(138):1543-1550.
猜你喜欢
检验医学与临床(2022年20期)2022-10-24
成都医学院学报(2022年4期)2022-08-19
江西农业学报(2021年4期)2021-04-20
中国动物传染病学报(2020年6期)2020-11-18
环球时报(2020-09-30)2020-09-30
三农资讯半月报(2020年11期)2020-06-21
灾害医学与救援(电子版)(2018年1期)2018-06-05
中国全科医学(2015年33期)2015-12-02
中国当代医药(2015年9期)2015-03-01
