
人和动物生命质能方程和寿命曲线
2012-08-04 09:16:22刘春春李树蕾李艳超张艳凤谭百宏
中国老年学杂志 2012年13期
刘春春 李树蕾 陈 刚 李艳超 张艳凤,4 谭百宏
(吉林大学白求恩医学院基础医学实验教学中心,吉林 长春 130021)
按照世界卫生组织调查数据,人的寿命15%取决于遗传因素,10%取决于社会因素,8%取决于医疗条件,7%取决于气候影响,60%取决于个体生活方式和行为习惯〔1〕。现代科学证明:生命过程中存在一个生物能量传递过程,生物消耗能量越多,寿命越短〔2,3〕。爱因斯坦通过一个理想的实证得到了一个方程E=M·C2〔4〕。当物体能量的变化为△E时,一定会伴随着质量变化(△M),△E=△M·C2,这一关系反过来也成立,这就是著名的质能方程,其所涉及能量形式可推广到所有形式〔5〕。那么,爱因斯坦质能方程能否且如何解释生命体能量变化呢?本文将通过推演生命质能方程和绘制质能时间曲线对人和动物寿命的奥秘作一次开创性探索。
1 材料与方法
1.1 生命质能方程建立 按照质能方程,质量即是能量,即使是极小的一块物质,由于具有质量,都蕴藏着巨大能量〔4〕。体重对应的质量也属于能量,这就意味着体重增加,能量也增加。但是绝大部分体重只是能量的一种储备形式,其中只有很少一部分可以被调用(△M可调)(主要参与氧化反应),转化成为活动的能量。不参与氧化的质量(设为Wi)对应的能量是“静”的能量,Wi=体重-△M可调。人类骨骼在17~18岁左右成熟,此时对应的体重为骨龄成熟标准体重。达到骨龄成熟点后骨龄标准体重基本不变或略有下降(图1)〔6〕。引入骨龄标准体重可以排除肥胖因素对寿命方程的干扰,因此所用的体重均为骨龄标准体重。
1.2 生命质能方程建立的前提 ①生命体某一时点可调用能量(E可调)为机体在单位时间内△M可调参与氧化反应所产生的能量。E可调是生命体中最活跃的能量,用来满足各种生命活动,所以它是“活”的能量。E可调=△M可调·C2(1)。②生命体某一时点E消耗包括:1)基础代谢耗能(Eb)。生命体某一时点在清醒而又极端安静状态下,不受肌肉运动、环境、温度、食物及精神紧张(意识活动)等影响时所消耗的能量为Eb〔7〕。Eb与体重(W+△M可调)大小关系密切,对于某一生命个体而言,体重增加,Eb也相应增加,所以设定某一时点Eb为Ebi=Kb·(Wi+△Mi可调)(2)。在狭义相对论中,质量、时间和能量是相互联系在一起的,并且都与速率有关,即E=M·C2,所以Kb为Eb速率的平方〔(m/s)2〕。一般认为基础代谢率与体重的0.75次方成正比〔8〕,这是通过研究不同种属动物体重和基础代谢率所得出的一个经验数值,并未去除肥胖因素。与基础代谢率的定义不同,我们所定义的Eb是针对同一个体不同骨龄体重而设定的。机体是细胞的集合,细胞越多,基础耗能越大,因此它与该个体骨龄标准体重正相关。2)生存活动必须耗能(Ebp)。生命体某一时点为了获得与Eb相当的能量必须付出的最小耗能。这一耗能包括觅食和进食、吸收和排泄、呼吸运动等为维持生命存在所消耗的能量。因为Eb与骨龄标准体重成正比,所以生存活动Ebp也与骨龄标准体重成正比,设某一时点生存活动Ebpi=Kbp·(Wi+△M可调)(3)。因为E=M·C2,所以Kbp为生存活动Ebp速率的平方〔(m/s)2〕。3)性欲耗能(ES)为生命个体某一时点因为性欲所消耗的能量。ES与体重和意识活动等多种因素有关,因此不能简单地用骨龄标准体重来衡量。4)高级活动耗能(En)。生命个体某一时点通过主观意识所参与的各种活动而消耗的能量总和,称为En。En是生命体在满足了生存的基础上的额外耗能,如为了实现人生目标、为了控制他人或为了更好地满足身体需要。En与生命体的意识等多种因素有关,因此也不能简单地用骨龄标准体重来衡量。本研究目的主要是研究生命体在健康条件下的寿命,即假想某种属的个体在良好状态下的生存年限(健康期望寿命)〔7,9〕。因为排除了遗传因素、社会因素、医疗条件、天灾人祸和气候等因素对寿命的影响,而是只考虑生命体自身活动对寿命的影响,所以设定生命体除了上述耗能外,没有其他方式耗能。根据1)、2)、3)和4),计算生命体某一时点的总耗能(见图1)。△E=(Kb+Kbp)(Wi+△M可调)+ES+En(4)。△E在生命体的不同时点不断变化,一般而言,△E随增龄有增加趋势(图1)。根据质能方程和方程式(4),计算生命体某一时点所具有的能量Ei=Wi·C2+△M可调·C2-(Kb+Kbp)(Wi+△Mi可调) -ES-En(5)。
1.3 生命存在的3个关键时间点 ①起始点:受精卵点。寿命的定义是生物从出生到死亡的时间〔10〕。但是因为同种属动物不同个体之间出生时间有所差异,即存在早产和晚产的情况,而不同种属的动物出生时间差异则更为显著,所以为了便于比较,我们将寿命定义为生物体从受精卵形成到死亡的时间。设受精卵形成的一刹那为时间起点(0 s),体重为受精卵的质量W0(kg),能量为E0(J)。②成熟点:从受精卵开始生命体经过生长发育达到骨龄成熟时,称为骨龄成熟点。骨龄评估能较准确地反映个体生长发育水平和成熟程度,当骨骼发育成熟时称为骨龄成熟点,它反映生命个体已经发育成熟。人骨龄成熟以后骨龄标准体重基本不变或略微降低,通过引入骨龄标准体重可以排除肥胖因素对推演寿命方程式的干扰。设骨龄成熟点时间为Tg(s),体重为Wg(kg),能量为Eg(J)。③死亡点。衰老是指机体随着时间推移对外界环境适应能力逐渐退化的过程〔11〕。尽管有各种理论来解释死亡(如端粒变短学说、心跳总数不变学说、细胞分裂次数不变学说)〔3,12〕,但机体死亡原因还是一个待解的谜。从耗能曲线和骨龄体重变化曲线(图1)来看,E消耗和体重变化均不能揭示死亡本质,但可以肯定的是,生命在死亡点时,M可调=0,E消耗=0。设死亡点时间为Td(s),体重为Wd(kg),能量为Ed(J)。机体死亡后经过降解而消失,最终质量变为0,能量变为0(图1),从死亡点到机体完全分解为自然降解过程,不属于生命存在范畴,所以不予考虑。按照实验中所设定的条件和参数计算各时点能量,并根据生命体各生存时间点和该时点对应的能量,绘制生命质能和生存时间曲线(即以时间为横轴,能量为纵轴,如图2)。
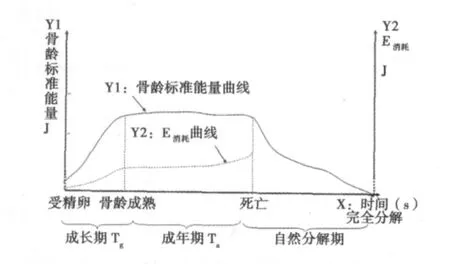
图1 骨龄标准能量曲线和能量消耗曲线
2 结果
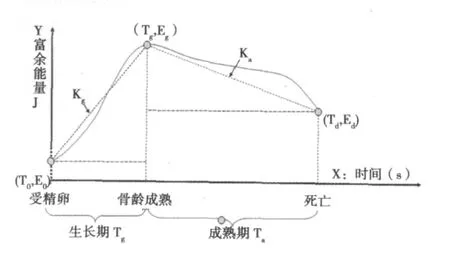
图2 富余能量和期望寿命关系曲线
2.1 各时点的能量 ①受精卵 (T0,E0)。根据方程式Ei=Wi·C2+△M可调·C2-(Kb+Kbp) · (Wi+△Mi可调)-ES-En(5)。在生命体起始点,Kb=0,Kbp=0,ES=0,En=0,所以受精卵点,生命体所具有的能量E0=W0·C2(6)。②骨龄成熟点(Tg,Eg)。从受精卵到骨龄成熟点的时间为成长时间Tg,设此时骨龄成熟标准体重为Wg,能量为Eg。根据方程式(5),在骨龄成熟点,生命体所具有的能量Eg=Wg· C2+△M可调·C2-(Kb+Kbp)· (Wg+△M可调)-ES-En(7)。在骨龄成熟点,当ES=0,即性活动开始之前,且En趋近最小时,生命体所具有的能量最大,所以理想的最大能量Eg=Wg·C2+△M可调·C2-(Kb+Kbp)·(Wg+△M可调)(8)。③死亡(Ta,Ed)。从骨龄成熟点到死亡点的时间为成熟期Ta,死亡时体重为Wd,能量为Ed。根据方程式(5),死亡点时△M可调=0,Kb=0,Kbp=0,ES=0,En=0,所以死亡时所具有的能量Ed=Wd·C2(9)。
2.2 根据生命存在的3个关键时间点计算成长速率(Kg)和衰老速率(Ka) ①Kg=(Eg-E0)/Tg,根据方程式(6)和(7),计算Kg=〔Wg· C2+△M可调·C2-(Kb+Kbp)·(Wg+△M可调)-ES-En-W0·C2〕/Tg(10)。②Ka=(Eg-Ed)/Ta。根据方程式(7)和(9),计算 Ka=〔Wg·C2+△M可调·C2-(Kb+Kbp)·(Wg+△M可调)-ES-En-Wd·C2〕/Ta(11)。
2.3 生存时间和能量的关系 TLife=Tg+Ta。根据方程式(11)计算寿命TLife=Tg+〔Wg·C2+△M可调·C2-(Kb+Kbp)·(Wg+△M可调)-ES-En-Wd· C2〕/Ka(12)。根据方程式(8)和(12),最长的理想寿命(健康期望寿命)TLife为:=Tg+〔Wg·C2+△M可调·C2-(Kb+Kbp) ·(Wg+△M可调)-Wd· C2〕/Ka=Tg+〔(Wg+△M可调)·(C2-Kb-Kbp)-Wd· C2〕/Ka(13)。
3 讨论
由于遗传、社会、医疗和气候等因素均非个人所能控制,所以我们在推演寿命方程时假设这些不可控因素对生命体总体而言是相同且不变的,即只考虑了个人行为对寿命的影响〔13〕。虽然健康期望寿命只考虑生存时间而不考虑生存质量,但了解影响健康期望寿命的因素,对提高生存质量依然具有重要指导意义。寿命曲线始于受精卵点。尽管受精卵质量很小,但质能并不为零,其数值大小取决于父母(遗传因素),代表个体的先天和种属差异。生命存在的基本特征不在于能量绝对值大小,而在于能量不断变化,机体E可调和机体E消耗均为零代表了生命终止〔14〕。从曲线看,死亡时绝对质量要远远超过受精卵质量,这意味着生长期积累的能量在死亡时仍有大量储备。生命体死后完全分解时质能为零,但是死亡后已不再属于生命存在范畴,因此不算入寿命。
需要说明的是,前提和假设人为地排除了肥胖和环境等多种因素,这导致能量和寿命方程不可避免地存在以下几点缺陷:①脂肪是机体能量的一种储备形式,但增加脂肪储备的同时使Eb和Ebp相应增加。虽然我们引用了骨龄标准体重去掉了肥胖因素对寿命的影响,但是却造成了我们的寿命方程式不能直接解释肥胖和寿命的关系。②我们将骨龄标准体重分解为Wi和△M可调,Wi和△M可调在实际上会发生相互转化。一个完美的寿命模型应体现这两种质量转化关系,因此我们的模型有待进一步完善。③我们假设遗传、社会、医疗和气候等不可控因素对于生命总体是相同且不变的,这虽然使生命个体的行为和寿命的关系更为简单化,但毋庸置疑的是,上述因素一直在影响着生物个体寿命。④我们没有对Es和En进行更深入的探讨,因为这两种活动在高等动物特别是人类都有主观意识的参与,而意识耗能是能量变化的更高形式,不可能用一个简单公式表达。尽管我们的能量和寿命方程存在上述不足,但是因为通过一系列假设将寿命和生命个体因素关系更简单化,因此得出的结论反而更透彻和直接。
从生命质能和寿命方程式,可以得出以下几点重要结论:①性活动和高级生命活动对寿命的负面影响。方程式(7)代表骨龄成熟点时生命体所具有的能量,因为生命存在的基本特征在于能量的不断变化,即(Kb+Kbp)不能为0,所以生命体所具有的能量最大值应在ES=0和En=0时。ES=0意味着生命体能量最大值应在性活动开始之前,即性活动开始越早,骨龄成熟时积累能量越少。En=0则意味着生命体各种高级活动的E消耗应该最小,即En越多,骨龄成熟时积累能量越少。根据方程式(12),因为(Kb+Kbp)不能为0,所以在其他变量保持不变的情况下,如果要延长寿命,就应该使ES和En最小,即减少ES和En。昆虫学家早就注意到性活动和寿命的关系:如果强制雄果蝇独身,其平均寿命约为65 d;若允许雄果蝇每天跟一只雌果蝇交配,寿命缩短14%;如果雄果蝇每天交配的雌果蝇数增至8只,寿命会缩短38%〔15〕。对人类而言,没有性活动不能简单地等于压制性欲。事实上,压制性欲是意识参与的高级生命活动,即使没有性活动,但是由于性能压抑会产生各种不良身心反应,其结果反而导致En增加,使寿命缩短〔10〕。脑的意识活动是能量变化的最高形式,要消耗机体近四分之一的能量,与此结论相一致的是,精神紧张和压力正在变成现代人发生猝死的重要诱因〔16,17〕。②在理想的假设条件探讨某一种属动物的寿命。根据方程式(12),当ES=0,En=0时,生命体所具有的能量最大,即理想的最大能量。对于在同一环境下生活的同种属动物而言,Wg、Wd、Ka及 Kb、Kbp、ES、En均为相对稳定的数值,因此方程式(13)可对不同种属动物寿命长短进行比较。
在寿命方程中,生长期长短与寿命的关系最为明确,即生长期Tg越长,该类动物寿命越长。如人的生长期为20~25年,而健康期望寿命为100~175年;与人相比,鼠生长期为6~8 w,其寿命仅为1~2年;犬生长期为2年,寿命为10~14年〔12〕。1794年Buffon考察了31种哺乳动物的首次繁殖期(性成熟期)和最长寿命时发现,动物生长期等于性成熟期的2倍,哺乳动物最高寿命约为其生长期的5~7倍。Robine等〔18〕将Buffon的31种哺乳动物增加为564种,依然证实了Buffon理论的正确性。生长期是性成熟期2倍的这一论点对人类很有启发。人类性成熟期为13~15岁〔6,12〕,那么人类生长期应该在26~30岁,按照方程式(7)和(12),在生长期尚未结束之前过早出现ES和En,不仅影响能量积累,还会缩短寿命。
寿命方程式(12)中Wg+△M可调即为体重,所以骨龄成熟时标准体重越大,寿命越长。许多统计数据的确表明动物体型越大,寿命越长〔12〕。如人平均体重为60 kg,而寿命为70~80年;与人相比,小鼠平均体重为0.03 kg,其寿命仅为1~2年。然而从寿命方程式(12)来看,体重与寿命的关系并不是简单的正相关,体重对寿命长短的影响还与多种因素有协同关系。因为〔(Wg+△M可调)(C2-(Kb+Kbp)-Wd·C2〕/Ka≥0,对于相同种属而言,每一个体生长期、Ka及死亡时骨龄体重(Wd)是相近的。因为C2为定值,所以Kb+Kbp值减少时,也就是基础耗能和生存Ebp减少时,寿命相应延长。
通过比较动物寿命和单位体重单位时间耗氧量(相当于Eb和Ebp)的关系,人们发现,小鼠代谢较快,寿命较短,而人代谢相对较慢,所以寿命较长〔10〕。在人类,女性基础代谢比男性低,女性寿命也比男性长3~5年〔3,10〕。因为基础代谢和生活节奏有关,所以有学者认为,生活速度快的动物相对短寿〔19〕。值得一提的是,现代人生活速度快,寿命却比古代人长,其原因应该归功于生存环境和医疗条件的改变〔10,12〕。
综上,生命体的健康期望寿命与生长期、骨龄成熟时标准体重,性活动及高级生命活动密切相关。由寿命与能量关系的曲线得到结论是,如果要长寿,那么在保障生存必需耗能之外应尽量减少性活动和高级活动能量支出。
1 郭普远.多视角看影响中国人寿命的因素〔J〕.中国老年保健医学,2008;6(4):3-4.
2 庞小峰.关于生物能量在生命体系中的传递的研究(Ⅰ)〔J〕.西南民族学院学报(自然科学版),1997;23(1):73-84.
3 吴华根.能量消耗决定人的寿命〔J〕.中共乐山市委党校学报,2011;13(4):92-5.
4 Einstein A.Does the inertia of a body depend on its energy content〔J〕?Ann Phys Lpz,1905;18:639-41.
5 罗旭东,马红孺.质能方程的一种简单推导方法〔J〕.江西科学,2005;23(4):418-20.
6 张绍岩,张继业,马振国,等.青少年骨龄标准身高、体重和体重指数生长图表〔J〕.中国法医学杂志,2009;24(5):308-11.
7 Degen A.Ecophysiology of small Dsert Mammal〔M〕.Berlin:Springer Verlag Berlin Heidelberde,1997:163-236.
8 Kleirer M.Body size and metabolism〔J〕.Hilgardia,1932;6:315-53.
9 方积乾,刘淑霞,王心旺.健康期望寿命的综合评价〔J〕.广州医院学报,2004;32(4):29-31.
10 吕维善,林竟成.人类的寿命研究〔J〕.国外医学·社会医学分册,1987;2(2):85-90.
11 周 彤,金锡鹏.职业与衰老〔J〕.中华劳动卫生职业病杂志,1996;14(3):170-2.
12 侯俊林,陈苇菁,连至诚.动物和人类的寿命与百岁老人〔J〕.中医药信息,2009;26(5):4-9.
13 Dever GE.Community health analysis:a holistic approach〔M〕.2nd ed.New York:Aspen Publishers Inc,U.S.1991:384.
14 田爱景,李宗荣,林雨霖,等.论生命及非生命信息运动的一般规律〔J〕.医学信息,1998;11(10):21-5.
15 Service PM.The effect of mating status on lifespan,egg laying,and starvation resistance in Drosophila melanogaster in relation to selection on longevity〔J〕.J Insect Physiol,1989;35(5):447-52.
16 刘静儒.试述精神因素对人体健康的影响〔J〕.陕西中医学院学报,1988;11(2):40-2.
17 曹保卫.28例青壮年冠心病猝死临床分析〔J〕.当代医学,2009;15(13):96-7.
18 Robine JM,Petersen HC,Jeune B.Buffon and the longevity of species in the light of history〔J〕.Dan Medicinhist Arbog,2008;36:97-108.
19 Hulbert AJ.Metabolism and longevity:is there a role for membrane fatty acids〔J〕?Integr Comp Biol,2010;50(5):808-17.
猜你喜欢
保健与生活(2022年10期)2022-05-06 04:15:13
中学生数理化·中考版(2021年10期)2021-11-22 07:26:46
科学24小时(2020年5期)2020-08-02 10:50:47
小哥白尼(趣味科学)(2020年5期)2020-05-22 06:29:38
妈妈宝宝(2018年9期)2018-12-05 02:19:48
心血管外科杂志(电子版)(2018年1期)2018-11-08 02:54:30
大自然探索(2017年7期)2017-07-25 00:26:55
小樱桃·童年阅读(2016年10期)2017-04-21 00:26:33
科学生活(2015年4期)2015-04-27 13:49:40
长江大学学报(自科版)(2013年18期)2013-04-14 01:25:46
