
缺血再灌注大鼠脑皮质Bcl-2、Bax和Caspase-3的动态表达
2012-09-12 06:07曲丽莹刘春春于海洋薛晓飞
中国老年学杂志 2012年13期
赵 航 王 敏 曲丽莹 刘春春 蔡 睿 于海洋 安 林 薛晓飞 刘 玲
(吉林大学白求恩医学院人体解剖学教研室,吉林 长春 130021)
近年,脑缺血再灌注损伤成为医学界研究的热点问题。研究显示,在脑缺血再灌注损伤的病理变化过程中细胞凋亡是主动的可逆的过程〔1〕。因此,对脑缺血再灌注损伤引起的神经细胞凋亡机制进行深入研究具有重要意义。细胞凋亡过程受多种基因的调控,Bcl-2、Bax、Caspase-3是凋亡信号传导途径中重要的调控基因。本实验通过建立大鼠脑缺血再灌注模型,观察脑缺血再灌注后Bcl-2、Bax、Caspase-3蛋白表达与再灌注时间的关系,初步探讨了脑缺血再灌注过程中的神经保护机制。
1 材料与方法
1.1 材料 氯化三苯基四氮唑(TTC)购自北京鼎国生物;RTPCR检测试剂盒购自 TaKaRa公司;Bcl-2、Bax、Caspase-3抗体均购自Santa Cruz公司;免疫组织化学检测试剂盒购自北京中衫生物技术有限公司。
1.2 方法
1.2.1 实验动物及分组 选择健康成年雄性Sprague-Dawley大鼠57只,重量250~280 g,由吉林大学动物实验中心提供。饲养环境温度18℃ ~22℃,光暗周期12 h。随机分为:假手术对照组(12 只)和缺血再灌注2、6、12、24、48 h 组(每组 9 只)。缺血再灌注组制备大脑中动脉闭塞(MCAO)模型,假手术组的手术过程与缺血再灌注组一致,但只暴露颈总动脉,并不进行插线。
1.2.2 MCAO大鼠模型的制备 应用Zea-Longa线栓法:经左侧颈外动脉-颈内动脉插线建立MCAO 2 h再灌注动物模型。实验进程中个别死亡的、蛛网膜下腔出血的大鼠剔除在外,确保每组完成实验的动物达到7只。
1.2.3 TTC染色法 假手术组和模型组术后不同时间点各取1只大鼠,立即断头取脑,放在-20℃冰箱10 min待脑组织稍硬后,沿脑垂直轴做冠状切片,片厚约2 mm,再置于2%的TTC溶液中,37℃避光孵育30 min。然后转移至4%的多聚甲醛中固定。
1.2.4 RT-PCR检测 Bcl-2、Bax、Caspase-3 mRNA表达 按照TaKaRa提取试剂盒操作说明从冷冻的脑组织(每组3只)中提取总RNA,测定RNA浓度,取1 μg总RNA进行逆转录合成cDNA。RT-PCR特异引物由上海生工合成,引物序列见表1。扩增产物经1.5%琼脂糖凝胶电泳分离,利用全自动数码凝胶分析仪(Tanon 2500R)检测目的cDNA。

表1 Bcl-2、Bax、Caspase-3、GAPDH 上下游引物序列
1.2.5 免疫组化检测 Bcl-2、Bax、Caspase-3蛋白表达 每组MCAO模型大鼠各3只,大鼠以37℃ 0.01 mol/L pH7.4磷酸盐缓冲液(PBS)200 ml心脏灌流,4℃预冷4%多聚甲醛内固定后,迅速分离大脑,置4%多聚甲醛固定24 h,石蜡包埋并切片(5 μm)。常规脱蜡,按试剂盒说明书操作,二氨基联苯胺(DAB)显色,苏木素轻度复染,脱水,透明,封片。
2 结果
2.1 TTC染色 TTC染色结果显示脑缺血再灌注侧脑组织有梗死灶,白色为梗死区,主要位于大脑皮质、基底节区。见图1。
2.2 RT-PCR 假手术组 Bcl-2、Bax、Caspase-3 mRNA仅有微量表达,与其相比,Bcl-2 mRNA在缺血再灌注2 h后表达开始增多,6 h明显增多,24 h达高峰,48 h开始减少;Bax mRNA表达较Bcl-2延后,2 h开始微量表达,6 h开始增多,增高趋势持续至48 h;Caspase-3 mRNA从6 h后表达微量增多,12 h略有增多,24 h明显增多,持续增高至48 h。见图2。
2.3 免疫组织化学染色 假手术组Bcl-2、Bax、Caspase-3阳性细胞表达少,与其相比,Bcl-2表达的阳性细胞数在缺血再灌注后2 h开始增多,6 h阳性细胞数显著增多,24 h阳性细胞数增多最为明显,48 h未见减少;Bax表达的阳性细胞数在缺血再灌注后2 h微量增多,6 h开始明显增多,增高趋势至48 h;Caspase-3阳性细胞数在缺血再灌注后6 h微量表达12 h逐渐增多,24 h增加明显,持续增多至48 h。见图3。

图1 TTC染色,白色区域为梗死区
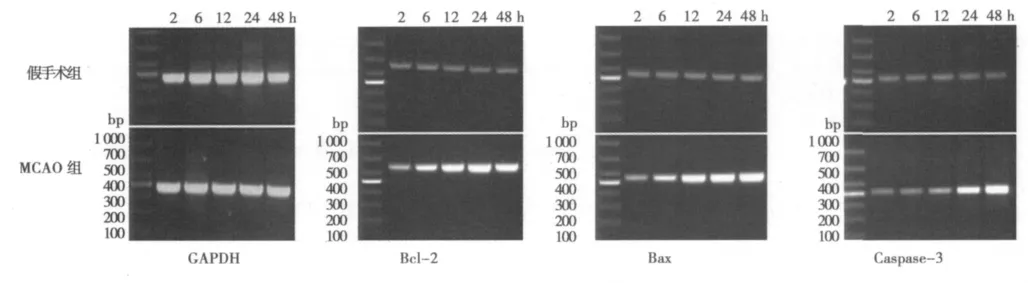
图2 Bcl-2、Bax、Caspase-3 mRNA 在缺血再灌注后 2、6、12、24、48 h 的表达
3 讨论
细胞凋亡过程受多种基因的调控,Bcl-2、Bax、Caspase-3被认为是凋亡信号传导途径中重要的调控基因〔1,2〕。研究证实细胞凋亡有两条途径,分别为细胞表面死亡受体途径和线粒体途径〔3〕。Bcl-2家族参与线粒体途径介导的细胞凋亡。Bcl-2和Bax是一对相互拮抗的凋亡相关因子,他们通过在线粒体外膜形成离子通道,阻止或促进细胞色素C的释放〔4,5〕。细胞色素C自线粒体释放入胞质,与凋亡蛋白活化因子-1(Apaf-1)、Caspase-9酶原结合,形成凋亡体。Caspase-9继而激活下游的Caspase-3。Caspase-3主要参与细胞表面死亡受体途径介导的细胞凋亡,是细胞凋亡下游通路的关键执行者,Caspase-3的激活可以引起一系列酶促级联反应,通过破坏核骨架及细胞骨架导致细胞凋亡〔2~7〕。由此可见,Bcl-2家族蛋白是联系细胞外凋亡途径和细胞内凋亡途径的重要纽带。Bcl-2和Bax通过表达的变化调控细胞色素C的释放,进而影响Caspase-3的激活。本研究发现大鼠局灶脑缺血再灌注损伤的12 h内,Bcl-2和Bax表达有逐渐增高的趋势,且Bcl-2的表达早于Bax,证明了Bcl-2蛋白的表达对神经细胞具有保护作用,并且这种保护通路在损伤早期就已经启动。本实验结果表明,在脑缺血再灌注的不同时间,Bax生成的上调与Caspase-3的激活密切相关。
总之,本研究结果提示在脑缺血再灌注的早期调控并促进Bcl-2的表达,减少Bax和Caspase-3的表达将会对降低缺血再灌注过程中神经细胞的损伤发挥重要作用。
1 Markus HS.Cerebral perfusion and stroke〔J〕.J Neurol Neurosurg Psychiatry,2004;5:353-61.
2 Memezawa H,Smith ML,Siesjo BK,et al.Penumbral tissues salvaged by reprefusion following middle cerebral artery occlusion in rats〔J〕.Stroke,1992;23(4):552-9.
3 Gross A,McDonnell JM,Korsmeyer SJ.BCL-2 family members and the mitochondria in apoptosis〔J〕.Genes Dev,1999;13(15):1899-911.
4 Rupniewska Z,Bojarska-Junak A.Apoptosis:mitochondrial membrane permeabilization and the role played by Bcl-2 family proteins〔J〕.Postepy Hig Med Dosw,2004;58:538-47.
5 Hu XL,Olsson T,Johansson IM,et al.Dynamic changes of the anti-and pro-apoptotic proteins Bcl-w,Bcl-2,and Bax with Smac/Diablo mitochondrial release after photothrombotic ring stroke in rats〔J〕.Eur J Neurosci,2004;20(5):1177-88.
6 Cheng EH,Kirsch DG,Clem RJ,et al.Conversion of Bcl-2 to a Bax-like death effector by caspases〔J〕.Science,1997;278(5345):1966-8.
7 Kirsch DG,Doseff A,Chau BN,et al.Caspase-3-dependent cleavage of Bcl-2 promotes release of cytochrome C〔J〕.J Biol Chem,1999;274(30):21155-61.
猜你喜欢
粉末冶金技术(2021年3期)2021-07-28
昆明医科大学学报(2020年11期)2020-12-28
中国病理生理杂志(2020年3期)2020-04-03
野生动物学报(2020年1期)2020-02-21
模具制造(2019年7期)2019-09-25
中成药(2018年4期)2018-04-26
中国高原医学与生物学杂志(2017年4期)2017-03-08
中国康复理论与实践(2015年10期)2015-12-24
中国体外循环杂志(2015年3期)2015-12-08
