
封育对伊犁绢蒿种群营养元素分配格局及贮量的影响
2012-03-13 02:42:48郑伟
草业科学 2012年10期
郑 伟
(1.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐 830052;2.新疆维吾尔自治区草地资源与生态重点实验室,新疆 乌鲁木齐 830052)
植物资源分配(Resource Allocation)是生活史理论的核心之一[1-2],是指植物将同化的资源分配于不同功能器官的比例[2]。植物能在环境资源限制和干扰条件下将光合作用所固定的能量优化分配从而最大化其在多变环境中的适合度[3]。碳(C)是植物光合作用的产物,其贮存量及分配格局往往作为植物资源分配格局的重要依据[4]。因此,许多学者在考虑植物资源分配格局时,多以生物量进行测度[5],但营养元素有时是比生物量更重要的资源,植物体内关键营养元素分配也是物种生态策略的反映。高甲荣等[6]就认为营养元素积累规律和分配格局是生态系统的基本特征,也是维持生态系统结构和功能稳定不可缺少的重要方面。氮(N)、磷(P)、钾(K)、钙(Ca)、镁(Mg)是植物的主要营养元素,其在植物体内的变化将导致整个系统生物地球化学循环特征的显著变化[7]。如N在世界范围内影响着植物的生产力[8],也在一定程度上决定着草地生态系统的物种分布[9]、丰富度[10]、物种入侵[11]、植被的退化和恢复[12]等。草地生态系统常面临着干旱和过度放牧两种干扰,造成草地生态系统可利用N十分匮乏[13],进一步加剧草地生态系统退化。Carlen等[14]的研究则表明,是植物对N的利用能力而不是N水平决定了物种的竞争力和对逆境的适应能力。P不仅是植物生长的重要限制营养因子,并且是地质时代尺度上生物生产力的限制性营养元素[15]。上述营养元素均来源于土壤,靠植物的根系进行吸收利用,因而与光合作用所产生碳水化合物的量及生长器官的数量和活力密切相关[16]。从草地生态系统尺度来看,草、畜产品的输出造成营养元素从系统中的流失;再加上草地利用由游牧转变为半舍饲放牧(白天放牧夜间入圈管理),也减少了营养元素向草地的返还[17]。由此导致草地生态系统养分循环失衡,主要牧草种群的营养元素供应受到胁迫。因此,研究植物体内主要营养元素的积累规律和分配格局对揭示植物种群在不同环境压力下的适应对策变化有重要意义[16,18]。
绢蒿荒漠草地广泛分布于新疆北部沙漠边缘、山前洪积扇、低山及低山丘陵和山前平原,南部天山和昆仑山中山区也有分布,是北疆山地植被垂直分布的基础植被之一;绢蒿荒漠草地除了具有生产功能外,在防止水土流失、防风固沙、生物多样性保护、维护山地草地生态平衡和平原绿洲区生态安全等方面发挥着不可替代的作用[19]。近年来,由于极端气候频发,超载无序放牧得不到有效控制,以伊犁绢蒿(Seriphidiumtransiliense)为主体的草地退化严重,其生产功能以及生态功能严重退化[20-21]。围栏封育可以使草地环境进一步得到改善,能科学地恢复草地植被,保护草地生态环境,且投资少、见效快,已成为当前新疆部分退化草地恢复的重要措施之一。目前对伊犁绢蒿种群的生理生态、再生性能、生物量与贮藏营养物质分配、幼苗分布格局等均有报道[19,22-26],但对伊犁绢蒿营养元素分配方面的报道甚少。本研究以绢蒿荒漠草地上的伊犁绢蒿为研究对象,从不同构件的生物量、C与营养元素含量、贮量及分配格局入手,揭示围栏封育对其资源分配格局和生态策略的影响,以期为绢蒿荒漠草地资源持续合理利用及植被恢复等方面提供科学依据。
1 材料与方法
1.1研究区概况 研究区位于天山北坡中段的山前倾斜洪积扇平原,地理位置43.858°~43.870° N, 87.136°~87.142° E,距乌鲁木齐市60 km,海拔825~897 m,地形起伏平缓。属典型的温带干旱大陆性气候,年降水量较少,为180~190 mm,年均温6.5 ℃。土壤为灰漠土,成土母质为黄土状物质,土层较厚,土壤较贫瘠,表层(0~10 cm)土壤有机质在10~15 g·kg-1[26]。围栏外,土壤表层结构被严重破坏,加上植被盖度小,土壤水分蒸腾快;围栏内,土壤结构的完整性,土壤透气透水性能强以及水分条件充足。该地区主要植被为伊犁绢蒿,春季有短生、类短生早春植物发育,主要包括天山郁金香(Tuliptianschanica)、腋球顶冰花(Gageabulbifera)、弯果胡卢巴(Trigonellaarcuata)等,夏季一年生植物可形成层片,主要包括叉毛蓬(Petrosimoniasibirica)、角果藜(Ceratocarpusarenarius)、紫翅猪毛菜(Salsolaaffinis)等。该类型草地为当地重要的春秋放牧场。每年春秋各放牧1次,春季在3月下旬-5月下旬,秋季在9月中下旬-11月,冬夏两季利用强度不大[19]。
1.2研究方法
1.2.1样地设置 2007年4月在研究区内设置样地,其长期处于过度放牧的状态。在研究区由北向南设置3条样线,统计包括伊犁绢蒿在内的植物种类、密度、生物量、盖度等指标,以上述指标作为划分退化梯度的参量,将整个地段划分为中度退化(Moderate Degraded Plot,MDp)、重度退化(Heavy Degraded Plot,HDp)和极度退化(Over Degraded Plot,ODp)3个梯度,具体划分方法见参考文献[26]。划分完后,在3个退化梯度的中心区域设置研究样地,然后对样地进行围栏,样地面积为40 m×40 m。2009、2010年8-9月每月中下旬在中度退化围栏内及各退化梯度围栏外各设6个1 m×1 m样方,两个月共48个样方,分别对其草地植被特征进行调查。其群落特征见表1。
1.2.2样品采集与处理 采样时间在伊犁绢蒿生长发育基本完全时,具体时间为2009和2010年8、9月中下旬,每年取样两次。在中度退化围栏内及各退化梯度围栏外的每个样地间隔1.0~2.0 m随机选定伊犁绢蒿1株,每样地分别采样40株。用收获法齐地面刈割采集地上部分;采用挖掘法采集地下部分,就地洗净。全部样品用聚乙烯密封袋分别盛装,带回实验立即称鲜质量,然后迅速分离成不同构件:根(Root,R)、根颈(Root Crown,RC)、营养枝(Vegetative Shoot,VS)、生殖枝(着生花或种子的枝条,不包括叶,Reproductive Shoot,RS)。先将样品置于105 ℃的烘箱中烘15 min进行杀青处理,然后调至65 ℃烘干至质量恒定。烘干后对不同构件分别称量,然后粉碎,过0.25 mm的尼龙筛。
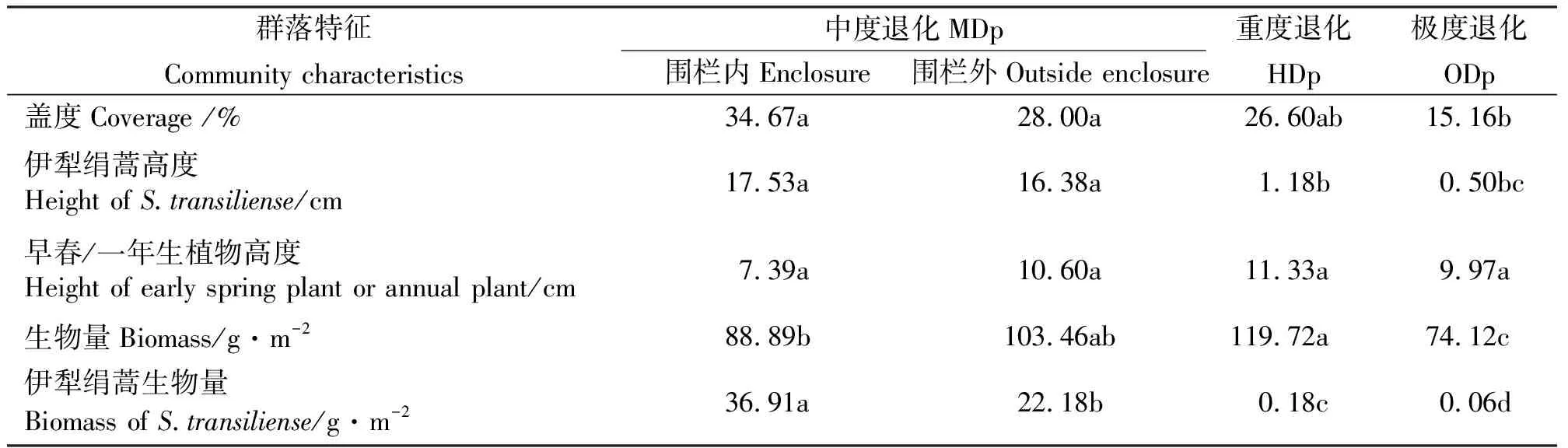
表1 围栏内外样地基本特征Table 1 Community characteristics in enclosure of moderate degraded plots and outside plots
1.2.3样品分析 参照鲍士旦的方法[27],对植物不同部位样品采用H2SO4-H2O2消煮,制备成待测液,以备同时进行N、P、K元素的测定。每个样品均设置平行样,以其算术平均值为结果,平行测定结果允许相差≤0.02 g·kg-1。N的测定采用凯氏定氮法;P的测定采用钼蓝比色法,使用日本岛津UV-2450可见光紫外分光光度计测定;K的测定采用火焰光度计法,使用上海欣益FP6410火焰光度计测定。原子吸收光度计法测定Ca和Mg,使用北京普析通用仪器有限责任公司TAS-990原子吸收光谱仪进行测定;采用重铬酸钾容量法测定C。
1.3数据分析 所有数据均为两年两次采样的均值。用各不同构件的生物量乘以各种元素的含量,即元素的贮量,再根据不同构件元素的贮量计算出分配情况。采用Excel 2007进行构建图表,采用SPSS for Windows 13.0软件包的one-way ANOVA进行方差分析,并利用LSD对中度退化围栏内及各退化梯度围栏外元素含量及比例进行比较,差异显著性水平P=0.05。
2 结果与分析
2.1伊犁绢蒿种群各构件营养元素含量 从不同退化梯度来看,极度退化样地伊犁绢蒿根、营养枝的N含量分别大于其他样地,根颈、生殖枝中N含量随着退化梯度的减轻而增加;围封后根颈、生殖枝中N含量显著增加(P<0.05),而根、营养枝则变化不大。未围封样地根颈、生殖枝中P含量随着退化梯度的减轻而显著增加(P<0.05),重度退化营养枝的大于其他退化梯度样地;围封后根中P含量显著增加(P<0.05)。根、营养枝中K含量以极度退化最高,根颈、生殖枝的K含量在不同退化梯度间无显著差异(P>0.05);围封后各构件K含量变化不大。极度退化样地根、生殖枝的Ca含量显著大于其他退化梯度样地(P<0.05),随着退化梯度的增加,根颈、营养枝的Ca含量呈下降趋势;围封后根颈、营养枝的Ca含量增加,且营养枝的Ca含量显著大于极度退化样地(P<0.05)。根的Mg含量以极度退化最高,根颈中以中度退化围栏外最高,营养枝中以重度退化最高,生殖枝中以极度退化最高;围封后各构件Mg含量变化不大。根和营养枝的C含量以重度退化最高,生殖枝中以中度退化围栏外最高;围封后各构件C含量未增加。从构件的元素含量来看,C含量为根>营养枝>根颈>生殖枝,N含量为生殖枝>根>根颈>营养枝,P含量为根>根颈>营养枝>生殖枝,K含量为营养枝>生殖枝>根>根颈,Ca含量为生殖枝>根>根颈>营养枝,Mg含量为营养枝>生殖枝>根颈>根(表2)。
2.2伊犁绢蒿种群各构件营养元素贮量的分布特征 生态系统营养元素积累为生物产量与各器官中营养元素之积,不仅取决于营养元素含量的高低,而且取决于生物量的大小。在8-9月生长的伊犁绢蒿种群,营养枝生物量大于其他构件生物量。随着退化梯度的减轻,生殖枝生物量呈增加趋势;围封后其进一步增加。营养枝中N的贮量最高,围封后根中P的贮量较高。营养枝中K的贮量最高,随着退化梯度减轻,其贮量呈增加趋势。营养枝中Ca的贮量较高,随着退化梯度减轻,其贮量呈增加趋势;极度退化样地则是根中Ca的贮量最高。营养枝中Mg的贮量较高,且围封后其贮量增加;极度退化样地则是根颈中Mg的贮量最高。营养枝中C的贮量最高,且围封后其贮量增加;营养枝、生殖枝营养元素总贮量在围栏封育后增加(表3)。
2.3伊犁绢蒿种群各构件营养元素分配格局 伊犁绢蒿营养枝生物量分配比例在各退化梯度均高于其他构件;随着退化梯度的减轻,生殖枝生物量分配比例呈升高趋势,根、根颈的呈下降趋势。随着退化梯度的增加,根的N分配比例呈增加趋势,根颈中N分配比例以重度退化样地最高,营养枝以极度退化样地最高,生殖枝中N分配比例随着退化梯度的增加而呈下降趋势;围封后生殖枝中N分配比例增加,且显著大于极度退化样地(P<0.05)。中度退化围栏外根中P的分配比例显著小于其他退化梯度样地(P<0.05);中度退化围栏外根颈、生殖枝中P的分配比例显著大于其他退化梯度样地(P<0.05);围封后根中P的分配比例显著增加(P<0.05),营养枝中P的分配比例也增加。根中K的分配比例随着退化梯度的增加而增加,且极度退化显著大于中度退化围栏外样地(P<0.05),根颈中K的分配比例以重度退化最高,显著大于其他退化梯度样地(P<0.05),中度退化围栏外营养枝中K的比例大于其他退化梯度样地,随着退化梯度的增加,生殖枝中K的分配比例呈下降趋势;围封后生殖枝中K的比例增加。随着退化梯度的增加,根中Ca的分配比例显著增加(P<0.05),根颈中Ca的分配比例重度退化大于其他退化梯度样地,随着退化梯度的增加,营养枝、生殖枝中Ca的分配比例显著降低(P<0.05);围封后各构件中Ca的分配比例变化不明显。随着退化梯度的增加,根中Mg的分配比例呈增加趋势,根颈中Mg的分配比例以极度退化样地最高,显著大于重度退化样地(P<0.05),极度退化样地营养枝中Mg的分配比例显著小于其他退化梯度样地(P<0.05),极度退化样地生殖枝中Mg的分配比例显著大于其他退化梯度样地(P<0.05);围封后营养枝中Mg的分配比例增加。营养枝中C的分配比例重度退化显著小于其他退化梯度样地(P<0.05),生殖枝中C的分配比例随着退化梯度的减轻呈增加趋势;围封后营养枝中C的分配比例增加(表4)。
根、营养枝中N/C在各退化梯度样地间无显著差异(P>0.05),根颈中N/C随退化梯度的增加而减小,生殖枝中N/C极度退化显著小于其他退化梯度样地(P<0.05);围封后根颈、生殖枝中N/C增加。根中P/C重度退化显著大于其他退化梯度样地(P<0.05),根颈、生殖枝中P/C随着退化梯度的增加呈下降趋势,营养枝中P/C重度退化显著大于极度退化样地(P<0.05);围封后根中P/C增加明显,而生殖枝中显著下降(P<0.05)。根颈、营养枝中K/C样地间均无显著差异(P>0.05),根中K/C极度退化显著大于其他退化梯度样地(P<0.05),生殖枝中K/C重度退化显著大于中度退化围栏外样地(P>0.05);围封后各构件中K/C变化不明显。根中Ca/C随退化梯度的增加而显著增加(P<0.05),根颈、营养枝中Ca/C随退化梯度的减轻而增加,生殖枝中Ca/C在各退化梯度样地间无显著差异(P>0.05);围封后根、营养枝中Ca/C增加。根中Mg/C样地间无显著差异(P>0.05),根颈中Mg/C重度退化显著小于其他退化梯度样地(P<0.05),营养枝中Mg/C极度退化显著小于其他退化梯度样地(P<0.05),生殖枝中Mg/C极度退化显著大于其他退化梯度样地(P<0.05);围封后根、营养枝中Mg/C增加。
3 讨论与结论
植物体内营养元素含量的变化反映了土壤供应速效养分能力的差异和营养元素的丰富程度[7],因此,植物体内养分含量的变化可以指示土壤功能的变化和土壤的退化状况。本研究表明,围封后,伊犁绢蒿根中P的含量和根颈、生殖枝中N的含量以及根颈、营养枝中Ca的含量均显著增加,这说明围封后土壤N、P及Ca供应能力有所改善。围封对伊犁绢蒿K、C、Mg含量影响则不明显。不同的植物其构件营养元素含量的变化一方面受植物种群本身的特性影响,如冷蒿(Artemisiafrigida)和星毛委陵菜(Potentillaacaulis)在同一放牧强度处理下,其构件N、P含量的变化趋势有差异:随着放牧强度的增加,冷蒿叶中N含量下降,而星毛委陵菜地上部分N含量增加;冷蒿根中P含量增加,而星毛委陵菜地下部分变化不明显[7];而另一方面则受到不同生态对策及生活型(单子叶、双子叶)影响[18]。因此,伊犁绢蒿在围封后生态对策的变化可能是其构件营养元素含量变化的主要因素之一。
草地植物营养元素积累的差异不仅表现在草地植物各营养元素贮量的不同,还表现在各营养元素积累的构件类型上存在较大的差异[28-30],另外还可能受到放牧、干旱等外界干扰因素的影响[7,31]。本研究表明,伊犁绢蒿营养枝中C、N、K、Ca、Mg的贮量较高,根中P的贮量较高。营养枝、生殖枝所有营养元素总贮量在围栏封育后大都增加。随着退化梯度的减轻,生殖枝、营养枝生物量呈增加趋势,围封使其进一步增加。C是植物光合同化资源的产物,与生物量密切相关,因此,其积累规律与生物量相同。伊犁绢蒿退化严重的样地主要受到放牧干扰的影响,而放牧不仅影响植物的生产力,而且可以引起营养元素在植物不同构件中的分配发生变化,进而影响元素的周转[32-33]。本研究表明,伊犁绢蒿营养枝生物量分配比例均高于其他构件;而随着退化梯度的减轻,生殖枝生物量分配比例呈升高趋势,根、根颈生物量分配比例呈下降趋势;围封后生殖枝中的N、K分配比例与营养枝中Ca、Mg、C的分配比例增加。这说明在8、9月生长的伊犁绢蒿主要将资源投入到地上生长,特别是营养生长。由于伊犁绢蒿是多年生植物,生殖生长的分配比例较低,再加上放牧的干扰,生殖生长更受到限制。围封后放牧干扰消失,生殖生长限制因素减少,其投入的资源增加,为伊犁绢蒿种群的更新发展提供更多机会。伊犁绢蒿不同构件的生物量(包括C贮量)、营养元素贮量及其分配对围封表现出不同的响应规律,与其他草地植物在环境干扰(如放牧、N添加、雪被厚度等)下的响应规律相比[7,17,34],不同构件对围封具有“特异性”,根、根颈属于“贮藏者”,对环境变化不敏感;生殖枝、营养枝属于“指示者”,对环境变化响应比较迅速。

表2 围栏内外伊犁绢蒿各构件营养元素及C含量Table 2 Elements contents of S. transiliense modules in enclosure of moderate degraded plots and outside plots g·kg-1
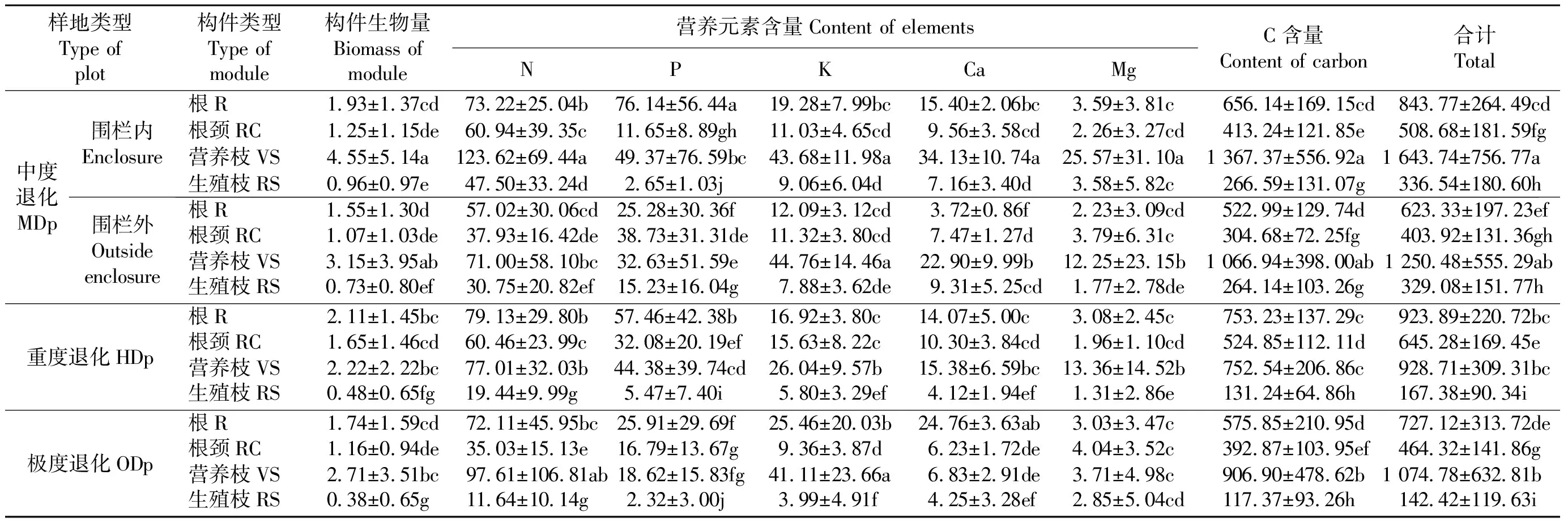
表3 围栏内外伊犁绢蒿各构件生物量、营养元素及C的贮量Table 3 Biomass and elements storage of S. transiliense modules in enclosure of moderate degraded plots and outside plots g·株-1

表4 围栏内外伊犁绢蒿各构件生物量、营养元素及C的分配比例Table 4 Proportion of elements and biomass of S. transiliense modules in enclosure of moderate degraded plots and outside plots %

表5 围栏内外伊犁绢蒿各构件营养元素贮量与C贮量的比值Table 5 Ratios of nutrient elements and carbon of S. transiliense modules in enclosure of moderate degraded plots and outside plots %
有研究表明,放牧可以增加植物的N/C,改善牧草的饲用品质[35],还可加速枯落物的分解,促进N的矿化[36]。对于草本牧草而言,地上部分N/C减少,意味着蛋白质含量降低、牧草品质的降低,其一方面降低了草食动物的生产力,另一方面还抑制了C、N的生物地球化学循环过程;地下部分N/C增加,改变了微生物的C、N环境,有利于微生物的分解活动,使N矿化作用加快,使草地放牧生态系统中N的损失加快,长此以往将造成N匮乏,使草地土壤更贫瘠,植物生产力更低[7]。随着退化梯度的减轻,伊犁绢蒿根颈中N/C呈增加趋势,生殖枝中N/C在围封后显著增加,而根和营养枝中N/C则变化不显著。伊犁绢蒿的根颈是介于地上与地下的连接部,是重要的贮藏器官,起到了缓冲与调节的作用,因此,伊犁绢蒿的营养元素分配格局有别于草本植物。当围封消除了放牧干扰后,伊犁绢蒿草地N损失受到抑制,伊犁绢蒿从土壤中吸收的N主要通过根颈来调节和分配;8、9月是伊犁绢蒿开花结实期,N分配于生殖生长的量得到增加。
经过3~4年的围封,伊犁绢蒿草地土壤的营养元素供给能力、伊犁绢蒿种群的生产力、营养元素分配及光合产物分配对策均发生了改变。其中,伊犁绢蒿草地土壤的营养元素供给能力、伊犁绢蒿种群的生产力得到显著提高,这是伊犁绢蒿草地恢复的重要内容。在8、9月生长的伊犁绢蒿种群主要将资源投入到地上生长,特别是营养生长,而围封使其生殖生长得到加强,有利于伊犁绢蒿种群的更新发展。因此,围封使伊犁绢蒿草地从土壤供给能力、伊犁绢蒿种群生产力和更新能力等方面得到恢复。
[1] Bazzaz F A,Grace J.Plant resource allocation[M].San Diego London:Academic Press,1997.
[2] 张大勇.理论生态学研究[M].北京:高等教育出版社,2000.
[3] Tilman D.Plant Strategies the Dynamics and Structure of Plant Communities[M].Priinceton,New Jersey,USA:Princeton University Press,1988.
[4] Watson M A,Geber M A,Jones C S.Ontogenetic contingency and the expression of plant plasticity[J].Trends in Ecology & Evolution,1995,10:474-475.
[5] 赵彬彬,牛克昌,杜国祯.放牧对青藏高原东缘高寒草甸群落27种植物地上生物量分配的影响[J].生态学报,2009,29(3):1596-1606.
[6] 高甲荣,张东升,肖斌,等.黄土区油松人工林生态系统营养元素分配格局和积累的研究[J].北京林业大学学报,2002,24(1):26-30.
[7] 李金花,李镇清,王刚.不同放牧强度对冷蒿和星毛委陵菜养分含量的影响[J].草业学报,2003,12(6):30-35.
[8] An Y,Wang S,Zhou X,etal.Plant nitrogen concentration,use efficiency,and contents in a tallgrass prairie ecosystem under experimental warming[J].Global Change Biology,2005,11(10):1733-1744.
[9] 白永飞,许志信,李德心.内蒙古高原针茅草原群落土壤水分和碳氮分布的小尺度空间异质性[J].生态学报,2002,22(8):1215-1223.
[10] Stevens C J,Dise N B,Mountford J O,etal.Impact of nitrogen deposition on the species richness of grasslands[J].Science,2004,303(5665):1876-1878.
[11] Sperry L J,Belnap J,Evans R D.Bromustectoruminvasion alters nitrogen dynamics in an undisturbed arid grassland ecosystem[J].Ecology,2006,87(3):603-615.
[12] 贾晓红,李新荣,李元寿.干旱沙区植被恢复中土壤碳氮变化规律[J].植物生态学报,2007,31(1):66-74.
[13] 潘庆民,白永飞,韩兴国,等.内蒙古典型草原羊草群落氮素去向的示踪研究[J].植物生态学报,2004,28(5):665-671.
[14] Carlen C,Kölliker R,Nösberger J.Dry mater allocation and nitrogen productivity explain growth responses to photoperiod and temperature in forage grasses[J].Oecologia,1999,121(4):441-446.
[15] Compton J S,Mallinson D J,Glenn C R,etal.Variations in the global phosphorus cycle[A].In:Glenn C R,Prevot-Lucas L,Lucas J.Marine Authigenesis:From Global to Microbia1[C].Tulsa,UK:Society of Sedimentary Geology,Special Publication Number 66,2000:21-33.
[16] 李金花,李镇清,刘振国.不同刈牧强度对冷蒿生长与资源分配的影响[J].应用生态学报,2004,15(3):408-412.
[17] 董晓玉,傅华,李旭东,等.放牧与围封对黄土高原典型草原植物生物量及其碳氮磷贮量的影响[J].草业学报,2010,19(2):175-182.
[18] 段飞舟,陈玲,阿里穆斯,等.草原植物种群营养元素生殖分配规律研究(Ⅱ)[J].内蒙古大学学报(自然科学版),2000,31(2):193-197.
[19] 鲁为华,朱进,王东江,等.天山北坡围栏封育条件下伊犁绢蒿幼苗分布格局及数量动态变化规律研究[J].草业学报,2009,18(4):17-26.
[20] Zhao W Y,Li J L,Qi J G.Changes in vegetation diversity and structure in respons Tianshan Mountains,China[J].Journal of Arid Environments,2006(4):21-33.
[21] 赵万羽.新疆草地资源的劣化、原因及治理对策[J].草业科学,2002,19(2):19-22.
[22] 宋智芳,安沙舟,孙宗玖.刈割和放牧条件下伊犁绢蒿生物量分配特点[J].草业科学,2009,26(12):118-123.
[23] 陈乐宁,安沙舟,孙宗玖,等.刈割对伊犁绢蒿根系贮藏营养物质的影响[J].干旱区研究,2009,26(6):859-864.
[24] 侯钰荣,安沙舟,张永娟.刈割强度对伊犁绢蒿叶功能性状及构件的影响[J].草地学报,2010,18(3):333-338.
[25] 靳瑰丽,朱进忠,刘洪来,等.退化伊犁绢蒿荒漠草地主要植物生理生态适应性研究[J].草地学报,2011,19(1):26-31.
[26] 刘洪来,朱进忠,靳瑰丽,等.伊犁绢蒿荒漠草地退化演替序列的划分[J].新疆农业科学,2007,44(2):137-141.
[27] 鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000.
[28] 钟华平,杜占池,樊江文.红三叶群落微量营养元素和有益元素积累量的分配与动态特征[J].草业学报,2003,12(5):81-87.
[29] 仲延凯,孙维,孙卫国,等.割草对典型草原植物营养元素贮量及分配的影响[J].干旱区资源与环境,2000,14(1):55-63.
[30] Witowski E T F,Lamont B B.Disproportionate allocation of mineral nutrients and carbon between vegetative and reproductive structures inBanksiahookeriana[J].Oecologia,1996,105(1):38-42.
[31] 许振柱,周广胜,王玉辉.干旱和复水对羊草碳氮分配的影响[J].气象与环境学报,2007,23(3):65-71.
[32] 李金花,李镇清,任继周.放牧对草原植物的影响[J].草业学报,2002,11(1):4-11.
[33] Ritchie M E,Tilman D,Knops J M H.Herbivore effects on plant and nitrogen dynamics in oak savanna[J].Ecology,1998,79(1):165-177.
[34] 刘琳,吴彦,孙庚,等.季节性雪被对青藏高原东缘高寒草甸 2 种优势植物碳、氮积累和分配的影响[J].植物研究,2011,31(4):451-460.
[35] McNaughton S J.Grazing as an optimization process:Grass-ungulate relationships in the Serengeti[J].The American Naturalist,1979,113:691-703.
[36] Frank D A,Inouye R S,Huntly N,etal.The biogeochemistry of a north-temperate grassland with native ungulates:nitrogen dynamics in Yellowstone National Park[J].Biogeochemistry,1994,26:163-188.
猜你喜欢
西北农林科技大学学报(自然科学版)(2023年1期)2023-02-20 03:57:48
草地学报(2022年9期)2022-09-29 11:22:48
草地学报(2021年10期)2021-11-09 02:00:46
山东农业科学(2018年7期)2018-10-22 07:01:10
草地学报(2018年1期)2018-07-30 03:29:08
热带农业科学(2018年2期)2018-06-21 11:27:12
速读·中旬(2017年11期)2017-11-11 12:41:26
理论导刊(2017年8期)2017-08-30 06:21:38
草原与草坪(2016年5期)2016-11-28 07:57:43
江苏农业科学(2014年9期)2014-11-15 04:01:30
