
紫蛇尾的生化成分及作为饲料原料对刺参幼参生长和存活的影响
2012-02-09 07:18:36姜森颢梁峻孙欣
饲料工业 2012年16期
姜森颢 梁峻 孙欣
(大连獐子岛渔业集团股份有限公司海洋生物技术研发中心,辽宁大连116503)
紫蛇尾(Ophiopholis mirabilis Duncan,1879)属于棘皮动物门(Echinodermata),蛇尾纲(Ophiuroidea)。全世界现生蛇尾纲动物约2 000种,分布于从潮间带到6 000 m海底的各大洋中,营底栖集群生活,在砂质、石质和珊瑚礁环境中生物量最为丰富,在寒带海洋和泥质底质环境中数量很少。紫蛇尾在我国主要分布于黄海中、北部,以大连、烟台和青岛外海分布量最大;刺参(Apostichopus japonicus)属于棘皮动物门(Echinodermata)、海参纲(Holothuroidea),主要分布于西太平洋北部,包括中国黄渤海海域、俄罗斯东部沿岸、日本和韩国沿岸。在日本,刺参按体色可分为3个品系:红刺参、青刺参和黑刺参,红刺参栖息在沙砾底质的近海,青刺参和黑刺参栖息在沙泥底质的近岸。2010年中国刺参产量10万吨,产值达200亿元,刺参已经成为我国最重要的水产养殖品种之一。
国内外学者对蛇尾和海参的研究主要包括资源分布、生理生态学、细胞生物学与再生以及生化成分和医药应用等方面。关于紫蛇尾生化成分的研究较少,刺参营养与饲料的研究较多,但以紫蛇尾为饲料原料进行刺参养殖的科学研究报道较少。本试验分析了紫蛇尾的生化成分,探寻了将紫蛇尾作为饲料原料进行刺参养殖的可行性,为蛇尾资源的开发利用和海参饲料产业的发展提供参考。
1 材料与方法
1.1 试验材料的来源与规格
试验用紫蛇尾于2011年8月采集于辽宁长山群岛獐子岛附近海域,平均体质量(0.64±0.19)g;刺参为大连獐子岛渔业集团广鹿基地海上生态繁育的一龄幼参,初始体质量(2.77±0.20)g。
1.2 饲养试验用饲料
试验饲料以鼠尾藻、海泥、紫蛇尾和牡蛎壳为主要原料。鼠尾藻和牡蛎壳采自獐子岛沿岸,海泥和蛇尾取自獐子岛附近海域。各原料经烘干,超微粉碎(200目以上)后,再混匀制成粉料。饲料原料配比与营养成分见表1。
1.3 饲养试验设计
饲养试验开始前,先将刺参幼参置于玻璃钢水槽(3 m3)中进行10 d的驯化。驯化期间每天投喂一次对照组饲料,过量投喂。驯养结束后,将试验刺参随机置于18个规格为70 cm×65 cm×70 cm的玻璃钢水槽中,每个水槽30头刺参。试验共设6个投喂处理(A、B、C、D、E和F),分别添加0%(对照组)、1%、2.5%、5%、10%和15%的紫蛇尾(干粉)(饲料配方见表1),每个处理3个重复,饲养试验为期4周。试验期间,每天换水、投饵一次,日投喂量按幼参体重的8%计算。每两周测定一次刺参体重,并统计其存活率,调整投喂量。试验用水为经过沉淀、砂滤的天然海水;试验过程中持续充气;光照周期12L∶12D。
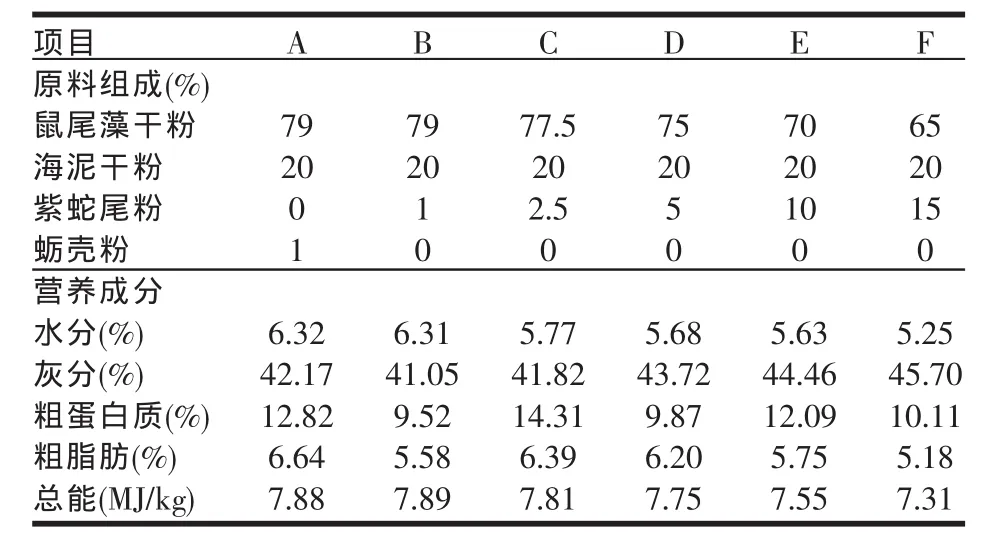
表1 饲料原料的组成与营养成分
1.4 紫蛇尾及饲料样品的生化成分分析
紫蛇尾及饲料样品的水分采用直接干燥法(GB/T 5009.3—2010)测定;灰分采用高温灼烧法(GB/T 5009.4—2010)测定;粗蛋白采用凯氏定氮法(GB/T 5009.5—2010)测定;粗脂肪采用索氏抽提法(GB/T 5009.6—2003)测定;总糖采用苯酚-硫酸法测定;饲料样品总能用PARR 1281氧弹热量计测定。
紫蛇尾样品氨基酸的测定:精确称取冷冻干燥样品20 mg,加入6 mol/l盐酸进行水解等处理后,用Biochrom 30氨基酸自动分析仪测定(GB/T 5009.124—2003)。色氨酸的测定:分光光度法(GB/T 15400—1994),对样品进行脱脂处理后,风干、混匀,用Spectrum SD-754紫外分光光度计测定。脂肪酸的测定:称取冷冻干燥样品0.2 g于10 ml带塞刻度试管中,经皂化(1 mol/l KOH/甲醇,2 ml)、甲酯化(2 mol/l HCl/甲醇,4 ml)等处理后,用Agilent 6890+气相色谱仪测定。无机元素的测定:火焰原子吸收分光光度法(GB 17378—2007),取鲜冻蛇尾样品0.5 g左右于微波消解罐中,加入3 ml硝酸和2 ml过氧化氢,加热板预处理,于微波消解仪内消解至溶液澄清,定容至10 ml后,用SOLAAR M6火焰-石墨炉原子吸收光谱仪测定。
1.5 计算公式与统计学分析
鲜样中某种营养成分含量(%)=干样中该营养成分含量×(1-水分含量/100);
氨基酸分(%)(AAS)=待评蛋白质中氨基酸含量(mg/g pro)/参考蛋白质中同种氨基酸含量(mg/g pro)×100;
化学分(%)(CS)=待评蛋白质中氨基酸含量(mg/g pro)/鸡蛋蛋白中同种氨基酸含量(mg/g pro)×100;
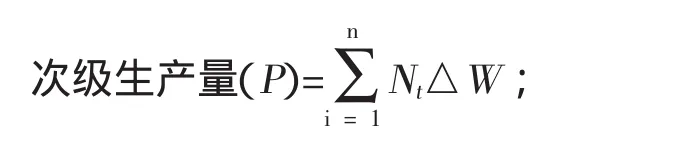
特定生长率(SGR)(%/d)=(lnWt-lnWt-1)/△t×100。式中:P——试验刺参的次级生产量(g/月);
Nt——调查时间t时的存活数量(头);
△W——为调查间隔期间的个体平均增重(g);
Nt△W——各阶段的生物量增长(g)。
SGR——特定生长率(%/d);
Wt——调查时间t时的体质量(g);
Wt-1——调查t-1时的体质量(g);
△t——两次调查时间间隔(d)。
采用统计软件IBM SPSS Statistics 19.0对不同饲料组数据(平均值±标准差)进行方差分析,Duncan's法多重比较检验差异显著性,以P<0.05作为各处理间显著差异的标志。
2 结果
试验期间水温变化为18.8~22.9℃,盐度31.38±0.11,pH值7.93±0.12,溶氧(6.17±0.32)mg/l,总氨氮<0.4 mg/l。
2.1 紫蛇尾的生化成分(见表2~表6)

表2 紫蛇尾一般营养成分含量(%)

表3 紫蛇尾的氨基酸含量(%)
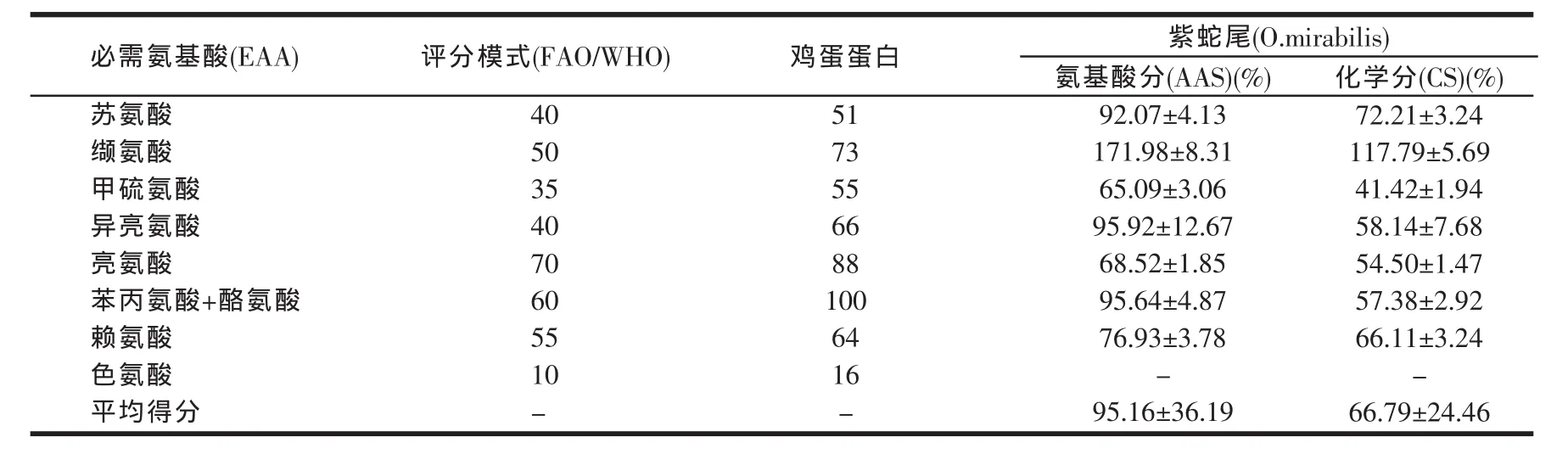
表4 紫蛇尾必需氨基酸组成的评价
由表3可知,紫蛇尾中共检出16种氨基酸,总含量(14.28±0.49)%。其中,谷氨酸含量较高[(1.96±0.09)%]、缬氨酸次之[(1.67±0.08)%],组氨酸含量较低[(0.32±0.03)%]。必需氨基酸、呈味氨基酸和药效氨基酸的含量分别占氨基酸总含量的(41.00±0.31)%、(45.75±0.38)%和(43.50±0.85)%。由表4可知,紫蛇尾所含必需氨基酸的氨基酸分和化学分除缬氨酸外均小于100。在FAO/WHO模式(AAS)与化学评分模式(CS)下,紫蛇尾第一限制氨基酸均为甲硫氨酸,第二限制氨基酸均为亮氨酸。
由表5可知,试验检出并标定紫蛇尾主要脂肪酸8种。其中,饱和脂肪酸(SFA)3种,单不饱和脂肪酸(MUFA)2种,多不饱和脂肪酸(PUFA)3种。SFA、MUFA和PUFA分别占脂肪酸总量的(42.71±2.14)%、(16.38±1.75)%和(40.91±1.31)%;在标定的脂肪酸中,EPA相对含量最高,C18∶2相对含量最低。
由表6可知,在测定的12种无机元素中,紫蛇尾Ca含量较高[(121 590.1±2 528.4)mg/kg]、Mg[(17 020.0±2 088.6)mg/kg]、Fe[(1 613.1±10.1)mg/kg]、Al[(756.67±89.07)mg/kg]次之,Co[(0.33±0.06)mg/kg]、As[(0.28±0.06)mg/kg]含量较低。
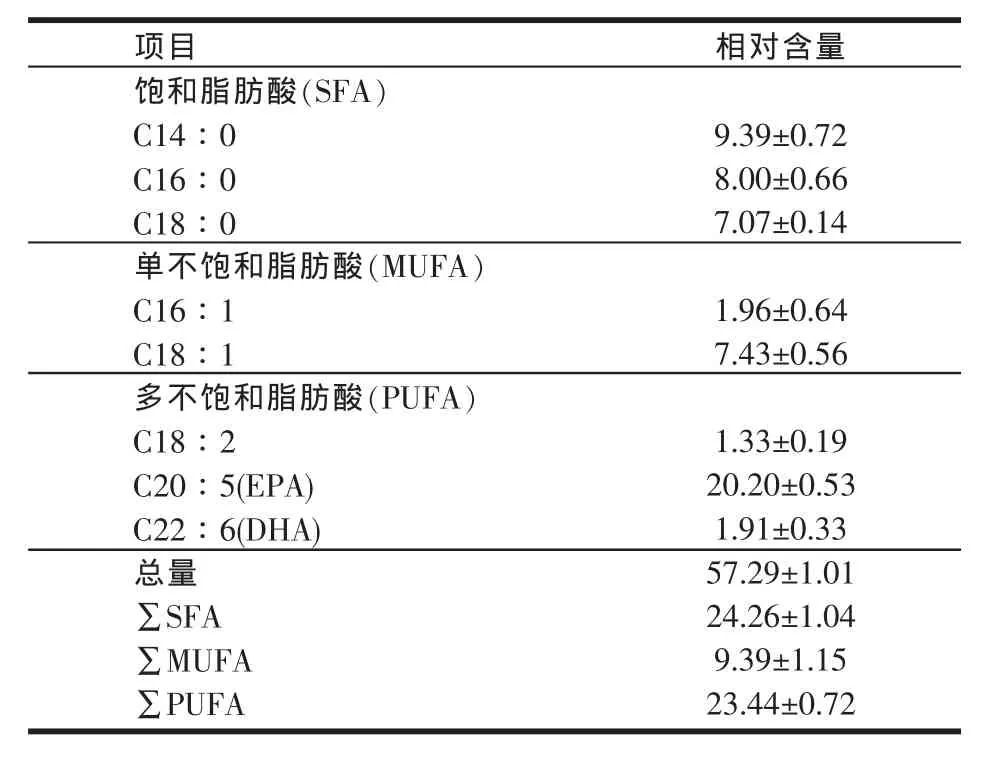
表5 紫蛇尾脂肪酸的组成与含量(%)
2.2 刺参的生长与存活
表7是试验期间在添加不同比例蛇尾饲料原料条件下,刺参幼参体重(Wt)、特定生长率(SGR)、次级生产量(P)和存活率(SR)的变化情况。结果表明:A组(对照组)试验刺参Wt显著小于D、E和F组(P<0.05);D、E、F组的SGR显著大于B组(P<0.05);A、B组次级生产量(P)显著小于D、E和F组(P<0.05);上述参数在D、E和F组之间的差异并不显著(P>0.05);试验刺参存活率在各处理组之间亦无明显差异(P>0.05)。
3 讨论
3.1 紫蛇尾的生化成分
紫蛇尾一般营养成分中除水分外,灰分含量最高,粗蛋白、总糖次之,粗脂肪含量最低。这一结果与其它的研究结果相一致。紫蛇尾体内的水分和粗脂肪含量低于刺参、马氏珠母贝(Pteria martensii)、海湾扇贝(Argopectens irradias)和大黄鱼(Larimichthys crocea)等水产经济动物以及猪肉、鸡蛋等主要畜禽产品。粗蛋白和总糖含量除低于刺参、马氏珠母贝和海湾扇贝外,也低于金氏真蛇尾(Ophiura kinbergi)和大西洋鲑(Salmo Salar)等;而灰分含量却远高于上述所有水产动物,这主要是由紫蛇尾特殊的网状骨骼结构所致。
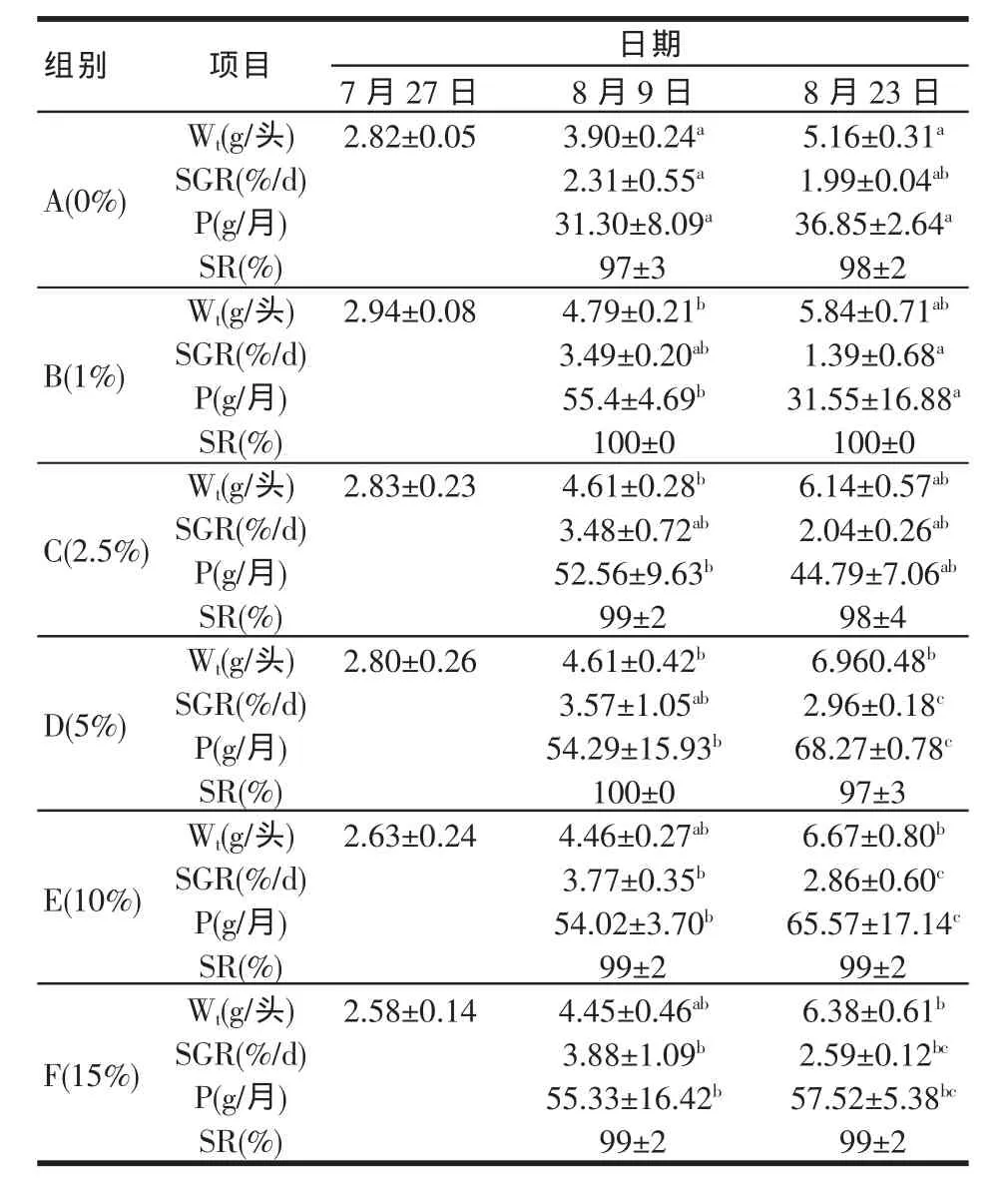
表7 添加不同比例紫蛇尾饲料对刺参的生长性能的影响
紫蛇尾中共检出16种氨基酸。其中,必需氨基酸的含量低于刺参体壁、梅花参(Thelenota ananas)和罗氏海盘车(Asterias rollestoni)高于金氏真蛇尾等棘皮动物;呈味氨基酸含量占氨基酸总量近46%,这保证了其作为饲料原料的诱食性和适口性;紫蛇尾中谷氨酸含量最高,而谷氨酸具有促进人体红细胞生成、改善与维持大脑功能的生理作用,是治疗神经衰弱和记忆力减退的有效成分。按照FAO/WHO的评价模式(AAS),紫蛇尾的必需氨基酸分在65~172之间(见表4),这一得分高于刺参。
试验共检测并标定8种主要脂肪酸,其中,SFA 3种、MUFA 2种、PUFA 3种(见表5),各种脂肪酸相对含量与其它作者基本一致。紫蛇尾体内SFA、MUFA和PUFA的相对含量均低于大黄鱼、鲐鱼(Pneumatophorus japonicus)和栉孔扇贝(Chlamys Farreri)等,但EPA的含量却远高于上述水产经济动物,同时也远高于刺参和金氏真蛇尾(Ophiura kinbergi)等棘皮动物。动物试验表明,EPA具有重要的生理功能和营养医疗价值,而紫蛇尾可作为EPA的稳定来源,应用于人类保健品开发和水产动物的饲料原料。
紫蛇尾含有的12种无机元素(Ca、Mg、Fe、Al、Mn、Zn、Ni、Cr、Cu、Se、Co和As)中,其中常量元素Ca、Mg、微量元素Fe的含量远高于金氏真蛇尾(Ophiura kinbergi)、沙刺参(Arenicola Simper)和黑乳参(Holothuria nobilis Selenka)等棘皮动物;微量元素Mn和Cu含量高于刺参,Zn含量低于上述动物,Cr和As含量(干品)均低于相关国家标准(GB 4810—94;GB 18406.4—2001)的限量。紫蛇尾体内含有的必需微量元素具有重要的生理功能,是人和海洋生物生命活动不可缺少的重要物质,其中某些元素已作为饲料添加剂在水产养殖中使用,并取得良好的效果。紫蛇尾常量与微量元素含量丰富、自然资源量巨大,可作为刺参很好的无机元素补充源应用于养殖生产。
3.2 紫蛇尾饲料原料对刺参生长和存活的影响及产业应用价值
刺参在水温超过25.5℃时夏眠,而体质量小于25 g的刺参并不夏眠。本试验中刺参体质量远低于25 g,且试验期间水温均低于23℃,刺参幼参在试验过程中保持正常生长状态,无夏眠现象。试验结果表明:较对照组(0%)和低比例添加组(1%和2.5%),试验幼参在紫蛇尾相对高比例添加组(5%、10%和15%)生长优势明显(P<0.05)(各组间存活率无差异);在相对高比例添加组之间,生长差异并不显著,这说明在一定范围内,更高的紫蛇尾添加比例,并没有带来更好的生长效果,但亦无负面影响。
从世界范围看,在某些海区特别是经济贝类养殖区,蛇尾、海星和海燕等棘皮动物生物量巨大,已经成为主要的敌害物种。对于这些棘皮动物的综合利用,国内外学者已经做了一些有益的探索,并在食品、保健、医药以及养殖业中取得了一定的成果。紫蛇尾作为一种新的低成本饲料原料来源能够很好地促进刺参的生长,其在水产养殖领域特别是海参养殖产业中的应用前景广阔。在海参保苗与养殖生产实践中,主要以市售和自配配合饲料为主。配合饲料中主要使用鱼粉作为动物性蛋白来源,然而鱼粉的品质并不稳定,同时,以更高等动物作为饲料来源饲养相对低等动物的做法并不是产业健康发展的方式,也无法得到水产养殖界的认可。配合饲料中大量使用鼠尾藻和马尾藻等大型藻类,而这些藻类在长期的掠夺性利用后资源量已所剩无几,且市场价格昂贵,因此,寻找合适的替代性饲料原料,突破产业瓶颈,就成了亟待解决的问题。紫蛇尾与刺参同属棘皮动物门,它们在营养组成上具有一定相似性,将紫蛇尾作为刺参的饲料原料可以很好地促进后者的生长。同时,紫蛇尾自然资源量极其巨大(特别是在一些经济贝类养殖区),因此,将其作为替代性饲料原料应用于刺参养殖生产是完全可行的。关于紫蛇尾作为饲料原料对刺参体成分影响等进一步研究工作已经展开。
4 结论
紫蛇尾营养成分丰富,对刺参的生长有很好的促进效果,可作为饲料原料应用于刺参养殖。
猜你喜欢
红蜻蜓·低年级(2021年12期)2022-01-19 05:18:32
红蜻蜓·低年级(2021年12期)2021-12-19 15:06:23
中国生殖健康(2020年5期)2021-01-18 03:00:06
大连海洋大学学报(2020年2期)2020-05-06 02:26:10
中国生殖健康(2018年5期)2018-11-06 07:15:56
中国洗涤用品工业(2015年8期)2015-02-28 19:02:49
养殖与饲料(2014年10期)2014-02-28 22:14:51
食品科学(2013年19期)2013-03-11 18:27:44
食品科学(2013年14期)2013-03-11 18:25:09
养禽与禽病防治(2012年3期)2012-08-01 06:15:50
