
不同植酸酶的热稳定性和生物学活性比较
2011-08-09 02:38唐德富娄瑞颍张兴会
饲料工业 2011年20期
唐德富 娄瑞颍 张兴会
植物性饲料中的磷多以植酸磷形式存在,植酸磷必须降解为无机磷后才能被动物利用。玉米和豆粕中植酸磷占总磷的比例分别为71%和58%,而该两种原料基本不含植酸酶。目前,肉鸡饲养中多以玉米-豆粕型日粮为主,肉鸡消化道内虽有部分饲料、消化腺、微生物来源的植酸酶,但酶活微弱,其对植酸磷的利用率不到10%。目前主要通过添加外源植酸酶降低植酸的抗营养作用。外源植酸酶发挥功效的前提是到达胃肠道时具有生物学活性,而饲料制粒过程中,由于高温高压蒸汽的加入和压模机的强力摩擦,使得植酸酶的活性受到损害。因此,提高植酸酶热稳定性对于植酸酶的推广应用具有重要意义。
本试验对3种植酸酶产品的热稳定性及其不同添加水平对肉仔鸡生长性能的影响进行了研究,旨在为植酸酶产品的选择及应用提供试验参考。
1 材料与方法
1.1 植酸酶
分别购买3种市售植酸酶产品A、B和C,经测定其实际酶活分别为10 859、9 002和5 126 U/g。1个植酸酶活性单位(U)定义为:在37℃、pH值5.5条件下每分钟从0.005 mol/l的植酸钠溶液中释放出1 μmol的无机磷所需的酶量。
1.2 植酸酶热稳定性测试
本试验在丹麦Kolding技术研究所完成。3种植酸酶的添加量均为5 000 U/kg饲料(推荐添加量的10倍),采用玉米-豆粕型饲料配方(制粒试验的饲料配方见表1),生产颗粒饲料一个批次120 kg。制粒条件为:初始温度18~22℃,调制蒸汽温度122℃,调制时间30 s、蒸汽压2 bar,制粒孔径3 mm,制粒温度90℃和95℃。取样检测制粒前后饲料中的植酸酶活性,计算饲料中酶活存留率。

表1 制粒试验的饲料配方(%)
1.3 试验动物和日粮处理
选取健康、体重相近的1日龄AA肉仔鸡770只,随机分为11个处理组,每个处理7个重复,每个重复10只鸡(公母各半),以粉料形式饲喂。
试验设正对照和负对照。正对照采用正常营养水平日粮,负对照日粮则在正对照基础上,将P和Ca分别降低0.23%和0.16%,其他营养水平保持不变。在负对照基础上添加A、B、C 3种植酸酶,添加量分别为500、1 000和1 500 U/kg饲料。对照组日粮配方及营养水平见表2。
1.4 饲养管理
全部鸡只采用层叠式笼养,肉仔鸡自由采食和饮水。第一周鸡舍温度保持在32℃,采用24 h光照,之后分别按每周5℃和每周1 h的速率降低室温及光照时间至25℃和18 h,鸡舍内采用纵向通风。1日龄、7日龄和14日龄分别对肉仔鸡进行马立克、传支、法氏囊病免疫,28日龄和35日龄分别对肉仔鸡进行传支和法氏囊病二次免疫。

表2 试验正负对照组日粮配方及营养水平
1.5 生长性能测定
试验期共42 d,分为前期(1~21 d)和后期(22~42 d),按周以重复(笼)为单位记录采食量,1日龄、21日龄和42日龄以重复为单位称试鸡重。试验期间准确记录淘汰鸡只或死亡鸡只体重,用于计算死亡率、校正采食量和料重比。
1.6 统计分析
采用SPSS16.0统计软件ANOVA程序进行单因子方差分析,差异显著则采用Duncan's法进行多重比较。显著水平设为0.05。
2 结果与分析
2.1 制粒后酶活存留率(见图1)
由图1可知,饲料在90℃和95℃制粒后,植酸酶产品A的酶活存留率为96.3%和90.7%,产品B和C的酶活存留率分别为 26.4%、24.2%和 12.6%、14.2%。植酸酶产品A的热稳定性明显优于产品B和产品C。
2.2 肉仔鸡生长性能(见表3、表4、表5)
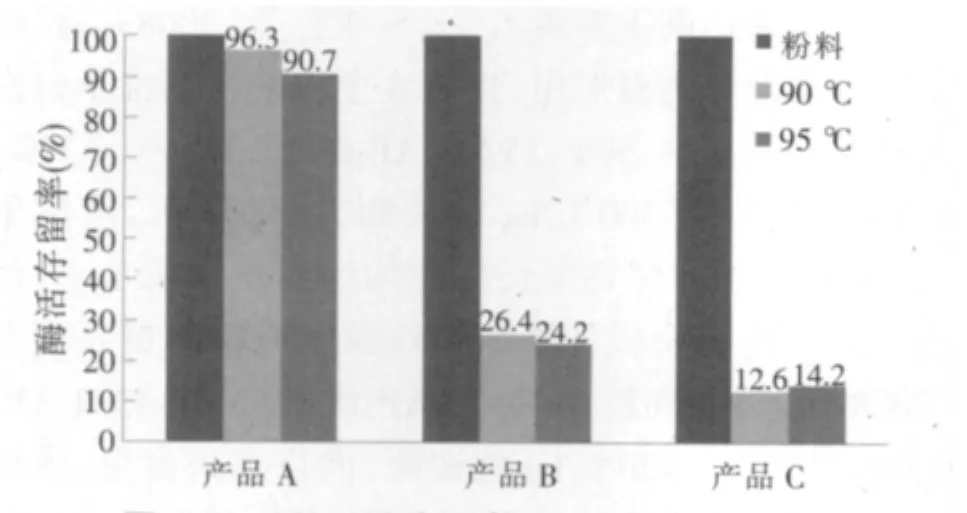
图1 3种植酸酶产品高温制粒酶活存留率
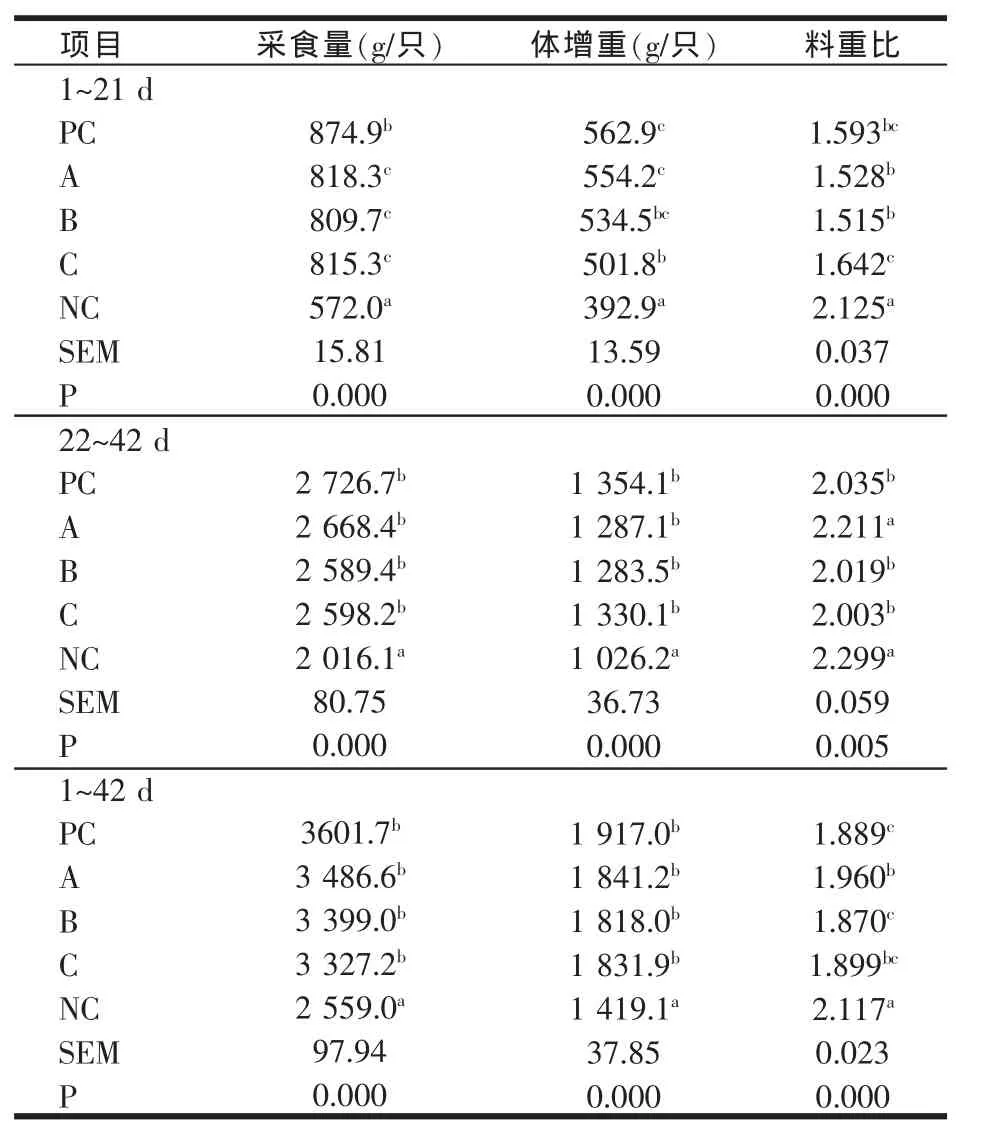
表3 500 U/kg植酸酶添加组肉仔鸡生长性能
本试验中,与正对照组相比,负对照组肉仔鸡生长性能显著降低(P<0.05),其中 1~21 日龄采食量、体增重分别降低了34.6%、30.2%,料重比提高了33.4%,22~42日龄采食量、体增重分别降低了26.1%、24.21%,料重比提高了13.0%,表明负对照组肉仔鸡钙和磷严重缺乏。
500 U/kg加酶组,与负对照组相比,产品A、C和B组肉仔鸡1~21日龄采食量分别增加246.3 g、243.3 g和237.7 g,体增重分别提高161.3 g、108.9 g和141.6 g,料重比分别降低了28.09%、22.73%、28.7%,A和B植酸酶添加组肉仔鸡生长性能与正对照组无显著差异(P>0.05),而优于C植酸酶添加组。22~42日龄肉仔鸡采食量和体增重在3种植酸酶添加组间无显著差异(P>0.05)。1~42日龄肉仔鸡全期采食量和体增重在3种植酸酶添加组间无显著差异(P>0.05)。
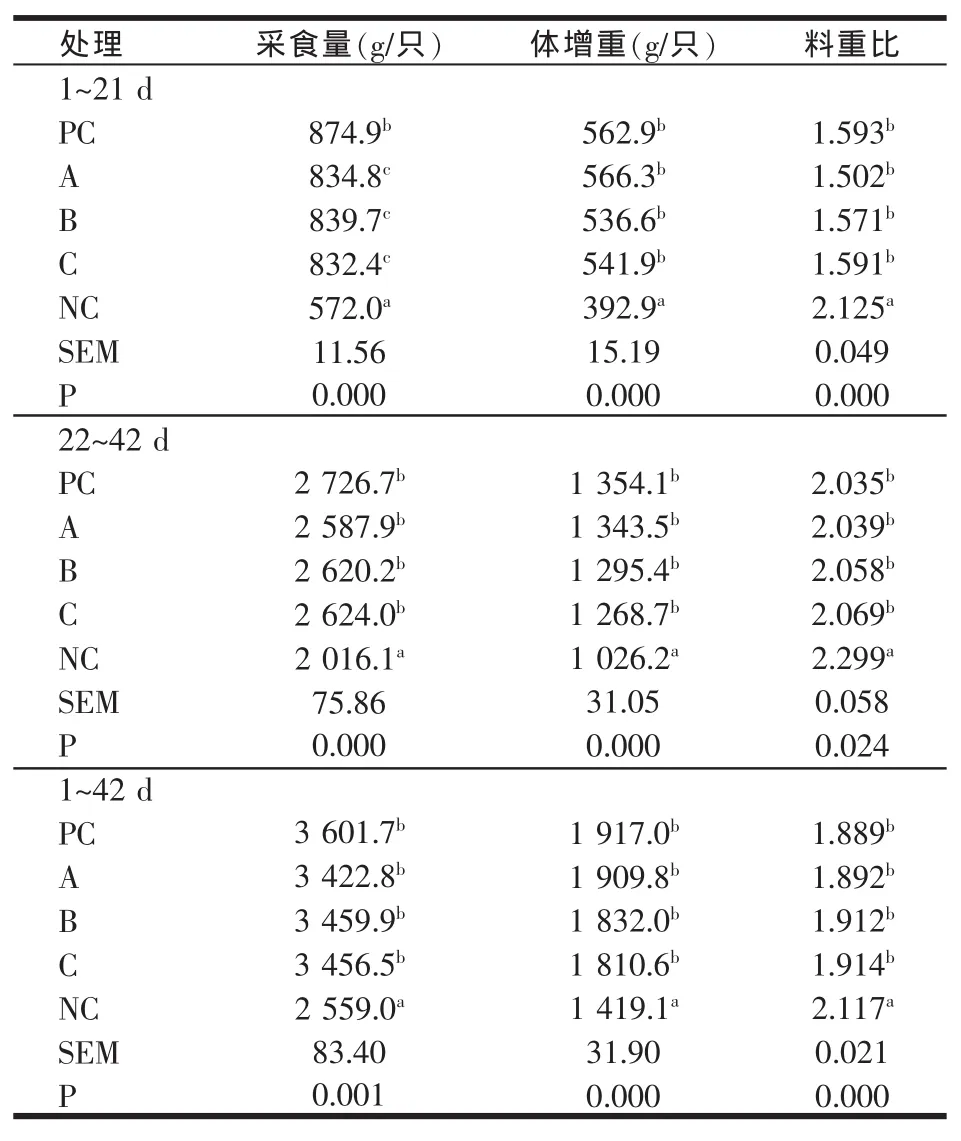
表4 1 000 U/kg植酸酶添加组肉仔鸡生长性能
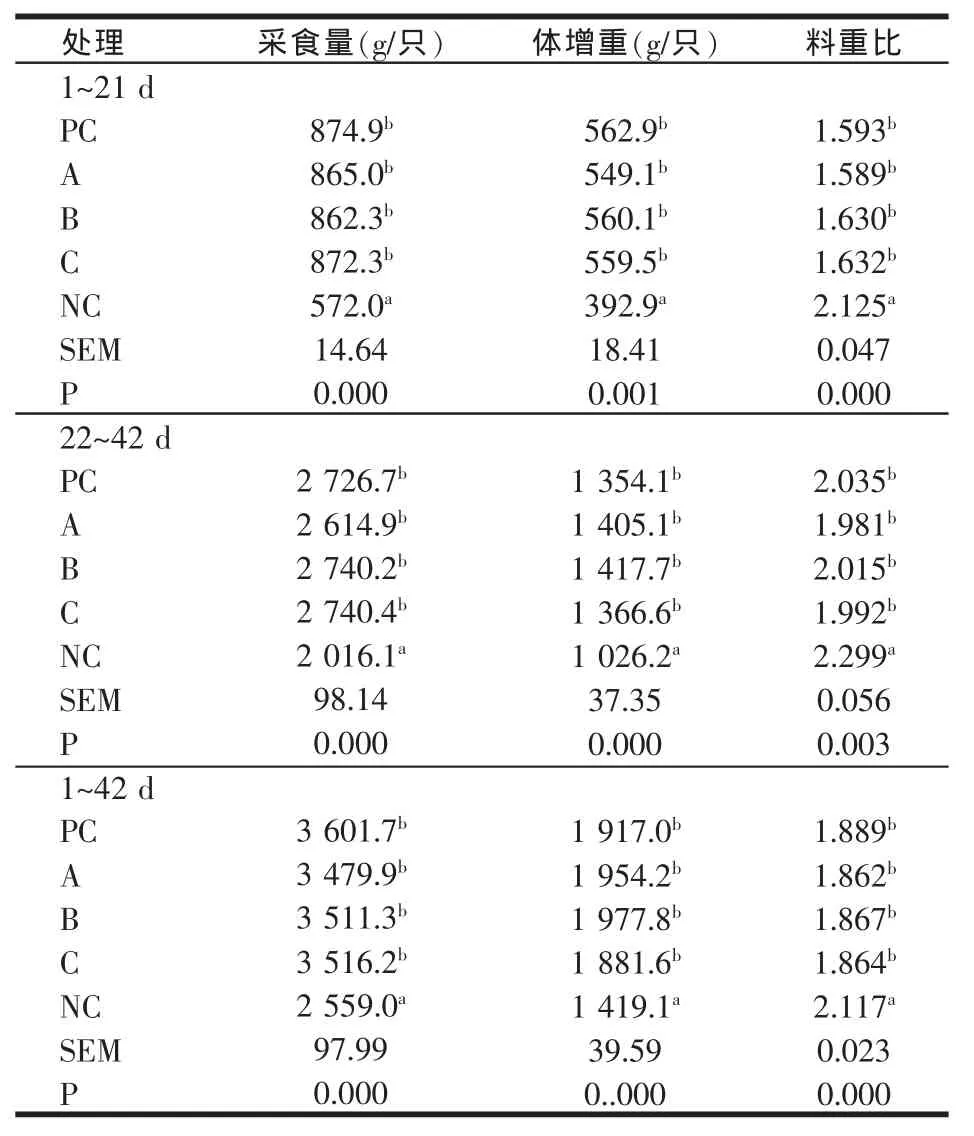
表5 2 000 U/kg植酸酶添加组肉仔鸡生长性能
添加剂量增至1 000 U/kg时,植酸酶添加组肉仔鸡除1~21日龄采食量显著低于正对照组外(P<0.05),生长后期和全期各组间肉仔鸡生长性能与正对照组相比无显著差异(P>0.05)。添加剂量增至2 000 U/kg时,各阶段肉仔鸡生长性能与正对照组相比均无显著差异(P>0.05),表明添加高剂量植酸酶释放的无机磷能够完全满足肉仔鸡对磷的需要。
随着植酸酶添加量的增加,肉鸡的体增重有提高的趋势,料肉比有下降的趋势。植酸酶产品A趋势较明显,植酸添加量由500 U/kg提高到1 000 U/kg和2 000 U/kg,肉仔鸡体增重分别提高3.7%和6.1%,料重比降低3.5%和5.0%。产品B和C添加量增至2 000 U/kg时,肉鸡体增重方表现出明显升高,与500 U/kg添加量相比,产品B和C组肉鸡体增重分别提高8.8%和2.7%。产品B和C组肉鸡料重比植酸酶各添加剂量间变化不明显。
3 讨论
3.1 制粒对植酸酶活性的影响
Slominski等(2007)将颗粒和粉末植酸酶在70℃保温30 s后,酶活分别损失80.6%和53.9%,将两种植酸酶在67℃制粒后,酶活损失分别为55.9%和50.6%,说明饲料制粒时的温度是影响外源植酸酶生物学活性的主要原因。许多学者从生产植酸酶的菌株着手,通过分离纯化或基因改组得到耐高温植酸酶。王放银(1995)将一种耐高温植酸酶置于90℃温度下30 min,其酶活损失不超过16%。赵春等(2007)在低磷日粮中(前期有效磷为0.356%,后期为0.289%)分别添加500 U/kg的粉状、液体状和包被颗粒状植酸酶,生长试验结果发现,包被植酸酶对肉仔鸡的促生长作用最大。这主要是由于对植酸酶进行包被处理,增强了酶在饲料制粒过程中的热稳定性,同时也降低了在胃液中的降解速度。因此,制粒前可通过包被或添加稳定剂的方法提高植酸酶的热稳定性。
本试验中,饲料在90℃和95℃制粒后,产品A的酶活存留率在90%以上,而产品B和产品C的酶活存留率不到30%(12%~26%),表现出了极大的差异。由于饲料植酸酶制造工艺和颗粒饲料制粒条件十分复杂,许多因素都会影响植酸酶的制粒稳定性,饲料厂迫切需要更多的此类研究来指导生产。
3.2 植酸酶对肉鸡生长性能的影响
研究表明,在低磷日粮中添加植酸酶,能显著提高肉仔鸡的采食量、体增重和饲料转化率(Yi,1996;谢新东,2002)。Ribeiro等(2003)在低磷日粮(AP:0.16%)中添加290 U/kg植酸酶,肉仔鸡采食量、体增重和饲料转化效率分别提高了58.29%、37.05%和13.3%,但未达到正常磷水平。杨维仁等(2008)以磷酸氢钙为磷源作为对照组,设定4个磷酸氢钙水平分别为对照组的75%、50%、25%、0(AP:0.42%、0.38%、0.32%、0.26%),并添加500 U/kg植酸酶,结果显示,磷酸氢钙+植酸酶组肉仔鸡生长性能均达到正常磷水平,但植酸酶不能完全代替添加的外源无机磷。黄兴国等(2008)在低磷日粮(0~21 d AP:0.36%;22~42 d AP:0.30%)中添加500 U/kg植酸酶,肉仔鸡采食量、体增重和饲料转化效率均达到正常磷水平,且存活率显著高于正常磷水平。Dilger等(2004)研究发现,在低磷日粮中(AP:0.24%)添加750 U/kg植酸酶组肉鸡生长性能优于1 000 U/kg植酸酶添加组。李桂明等(2008)在低磷日粮(0~21 d AP:0.23%;22~42 d AP:0.14%)中添加 0、250、500、750、1 000 U/kg 5 个梯度的植酸酶,结果显示以500 U/kg水平添加植酸酶,肉仔鸡获得最佳生长性能。由以上研究结果可知,植酸酶添加效果与日粮中无机磷水平和植酸酶添加剂量有关。
本试验与前人报道结果一致。在低磷日粮中添加植酸酶显著提高了肉仔鸡生长性能;肉仔鸡的生产性能随植酸酶的添加量增加而提高,但植酸酶产品A比产品B和C趋势明显。植酸酶的添加量主要取决于饲料中底物——植酸磷的含量,在纯玉米-豆粕型肉鸡日粮中由于植酸磷含量低,植酸酶的最佳添加量一般推荐500 U/kg饲料,在玉米-豆粕-杂粕(DDGS,农副产品)型肉鸡日粮中由于植酸磷含量很高,植酸酶的添加量可以提高到750或1 000 U/kg饲料,甚至更高,可以替代更多磷酸氢钙,节省更多饲料成本。
4 小结
从本试验结果可知,植酸酶产品A的热稳定性明显优于植酸酶B和植酸酶C。在低磷日粮中添加植酸酶显著提高了肉仔鸡生长性能,添加量500 U/kg时,植酸酶C在1~21 d的肉鸡体增重稍低。肉仔鸡的生产性能随植酸酶的添加量增加而提高,植酸酶产品A趋势较明显,添加量由500 U/kg提高到1 000 U/kg和2 000 U/kg,肉仔鸡体增重分别提高3.7%和6.1%,料重比分别降低3.5%和5.0%。植酸酶A在添加量1 000 U/kg时肉鸡体增重和正对照相当,而植酸酶B和C的相当添加量似乎需要2 000 U/kg。由于采食量的原因,产品B和C在料重比上没能反映出来,需要更多试验来验证;根据饲料组成和配方中植酸磷的含量,植酸酶的添加量可由传统的500 U/kg饲料提高到1 000 U/kg饲料及以上。
14篇,刊略,需者可函索)
猜你喜欢
中南大学学报(自然科学版)(2022年8期)2022-09-21
国外畜牧学(猪与禽)(2021年6期)2022-01-11
国外畜牧学(猪与禽)(2021年4期)2021-08-31
中国饲料(2018年22期)2018-12-13
中成药(2018年4期)2018-04-26
中国畜禽种业(2018年1期)2018-01-18
畜牧兽医科技信息(2015年5期)2015-12-27
兽医导刊(2015年8期)2015-03-25
食品科学(2013年13期)2013-03-11
浙江农业科学(2012年10期)2012-12-24
