广西鸭源H6N6亚型禽流感病毒HA和NA基因的序列分析*
2011-05-31 06:55:28莫胜兰陈义祥屈素洁粟艳琼陆文俊陈玉华
动物医学进展 2011年12期
莫胜兰,郑 敏,陈义祥,胡 杰,屈素洁,梁 媛,粟艳琼,2,陆文俊,苏 凯,陈玉华,李 军,刘 棋,2*
(1.广西动物疫病预防控制中心,广西南宁 530001;2.广西大学动物科技学院,广西南宁 530001)
水禽可以自然感染A型禽流感病毒的所有亚型,并通过多种途径直接或间接地将病毒传染给其他禽类和哺乳动物,被认为是禽流感病毒基因储存库和新毒株的重要来源,因此,在禽流感病毒的遗传进化和流行病学中具有重要意义[1-2]。根据表面糖蛋白血凝素(hemagglutinin,HA)和神经氨酸酶(neuraminidase,NA)抗原性不同,A型禽流感病毒(Influenza A virus)可分成16个不同的HA亚型和9个NA亚型[3]。在众多A型禽流感病毒亚型中,只有H5和H7亚型中的部分病毒才能引起高致病性禽流感,但越来越多的研究表明,禽类中的低致病性禽流感病毒(Low pathogenic avian influenza virus,LPAIV)往往通过基因重排方式产生新的病毒,一些新的病毒毒力增强或具有感染人的潜力。Wang C W 等[4-5]研究发现,对水禽无症状的 H6亚型禽流感病毒株在感染家鸡后会引起蛋鸡产蛋率下降和上呼吸道病变,甚至引起感染鸡死亡,造成较大的经济损失,其原因可能是H6亚型禽流感病毒在由水禽传染至家鸡的过程中毒力发生了变异而导致后者发病。另外,Chin P S等[6]报道,一株从野鸭体内分离到的 H6N2亚型禽流感病毒 A/teal/HongKong/W312/97(H6N2)与 能 致 人 死 亡 的H5N1亚型禽流感病毒株(A/Chick/HongKong/156/97(H5N1)的7个基因高度同源,推测前者很可能为后者提供了基因,使后者成为具有感染人能力的新病毒。因此,加强对H6等LPAIV的监测具有重要的公共卫生学意义。广西自治区动物疫病预防控制中心在对广西活禽市场进行禽流感病毒带毒状况的监测过程中,从健康家鸭中分离鉴定出一株H6N6亚型禽流感病毒,将其命名为A/duck/GX/038/2009(H6N6)(Dk/GX/038/09),并对其 HA和NA基因进行了克隆和序列分析,现将结果报告如下。
1 材料与方法
1.1 材料
1.1.1 病毒和试剂 禽流感病毒 Dk/GX/038/09由广西区动物疫病预防控制中心分离鉴定和保存;Trizol试剂为Invitrogen公司产品;AMV逆转录酶及相关试剂、Taq DNA聚合酶、Agarose Gel DNA Purification Kit、pMD-18T载体等为宝生物工程(大连)有限公司产品。
1.1.2 引物 依据GenBank上发表的H6N6亚型禽流感病毒HA和NA基因片段两端核苷酸保守序列采用Primer 6.0软件分别设计出2对引物,HA 基 因 引 物 6HA-R: 5′-ACAAGGGTGTTTTTCTTAAT-3′,6HA-F:5′-AGCAAAAGCAGGGGAAAA-3,预期扩增 HA基因片段为1 737bp;NA 基 因 引 物 6NA-F:5′-AGCAAAAGCAGGAGTGAAGA-3,6NA-R: 5′-TTTTCTAAAATTGCGAAAGC-3,预期扩增NA基因片段为1 449bp。引物由上海生工生物工程技术服务有限公司合成。
1.2 方法
1.2.1 病毒RNA的提取 取250μL病毒尿囊液,加入750μL Trizol试剂,混匀后室温静置5 min,加入200μL氯仿,轻微振荡混匀,4℃、12 000 r/min离心10min,取上清约500μL,加入0.5mL异丙醇,混匀后室温静置10min,12 000r/min离心10min,沉淀干燥后悬于适量的DEPC处理水中备用。
1.2.2 RT-PCR 取病毒RNA悬液15μL,与流感病毒随机引物1μL(20pmol/μL)混合,加入 AMV逆转录酶5×buffer 5μL、dNTP(10mmol/L)2 μL、RNasin 1μL(20U)、AMV 逆转录酶1μL(5 U),逆转录反应体系为25μL。混合物置于42℃作用60min。PCR反应体系:10×Taq聚合酶缓冲液5μL,dNTP(10mmol/L)2μL,上、下游引物(20 pmol/μL)各1μL,cDNA 2.5μL,Taq DNA聚合酶1μL(5U),用无菌水补足至50μL。PCR反应条件:95℃2min;94℃35s,53℃35s,72℃2min,35个循环;72℃10min。取PCR产物5μL,在10 g/L琼脂凝胶上电泳检查结果。
1.2.3 PCR产物的回收、克隆和序列测定 PCR产物用Agarose Gel DNA Purification Kit回收后,与pMD-18T载体连接,转化,蓝白斑筛选重组质粒并用PCR鉴定。每个样品挑取阳性克隆,由宝生物工程(大连)有限公司进行基因序列测定。
1.2.4 同源性分析和进化树构建 应用DNA Star软件对测序基因片段进行拼接和氨基酸序列推导,利用GenBank中的BLAST软件查找核苷酸和氨基酸的同源序列,并用MEGA 4.0软件绘制遗传进化树。
2 结果
2.1 HA和NA基因扩增
经10g/L琼脂糖凝胶电泳鉴定,扩增到的HA基因片段长约1.7kb,扩增到的NA基因片段长约1.5kb,两个基因的扩增结果与预期扩增长度一致(图1)。
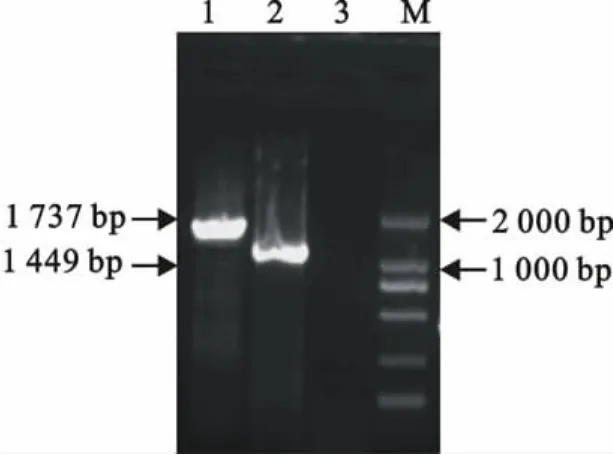
图1 HA和NA基因PCR扩增结果Fig.1 PCR amplification product of HA and NA gene
2.2 核苷酸同源性分析
测序结果显示Dk/GX/038/09的 HA基因全长为1 737bp。利用GenBank中的BLAST工具进行同源性搜索比对显示,Dk/GX/038/09的 HA基因与广东 A/duck/Shantou/17887/2005(H6N6)、A/duck/Shantou/8379/2005(H6N2)等 H6亚型毒株HA基因核苷酸同源性最高,达97.0%;与江苏A/duck/Jiangsu/022/2009(H6N6)和福建 A/duck/Fujian/12035/2005(H6N6)等 H6N6亚型毒株相应基因同源性则相对较低,分别为95%和93%。值得注意的是,Dk/GX/038/09的 HA基因核苷酸与广西2005年鸭源 H6N2毒株 A/duck/Guangxi/141/2005的HA基因同源性更低,只有86.5%。
对 NA 基因的测序结果显示,Dk/GX/038/09的NA基因片段长度为1 449bp,与GenBank中收录的参考毒株进行了同源性比较。结果表明,Dk/GX/038/09的 NA 基 因与广东 鸭源毒 株 如 A/duck/Shantou/20313/2005(H6N6)和福建鸭源毒株 A/duck/Fujian/11661/2005(H6N6)等的同源性最高,达98.0%;而与江苏鸭源毒株 A/duck/Jiangsu/022/2009(H6N6)的NA基因核苷酸同源性则只有88.6%,差异较大。
2.3 基因推导氨基酸分析
对Dk/GX/038/09的 HA基因推导氨基酸分析显示,其HA基因的编码区全长为1 701bp,编码566个氨基酸,HA1含有344个氨基酸,HA2含有221个氨基酸,不存在核苷酸缺失。HA1和HA2裂解位点的序列为 P-Q-I-E-T-R-D,仅含一个碱性氨基酸,具有典型低致病性禽流感病毒的特征序列。同时,该毒株HA受体结合位点氨基酸序列为K226-S227-P228-E229,与人类受体位点氨基酸序列不同,暂时不存在感染人的风险。
对Dk/GX/038/09的NA基因推导氨基酸分析显示该基因编码470个氨基酸,中间不存在任何氨基酸缺失。氨基酸序列比对发现,Dk/GX/038/09的NA基因氨基酸与广东鸭源毒株A/duck/Shantou/8365/2005(H6N6)的相应氨基酸同源性最高,为 99%;与 江 苏 鸭 源 毒 株 A/duck/Jiangsu/022/2009(H6N6)的相应氨基酸同源性较低,为93.0%。
2.4 HA和NA基因遗传进化树分析

图2 HA基因进化图Fig.2 Phylogenetic trees of HA gene

图3 NA基因进化树Fig.3 Phylogenetic trees of NA gene
HA基因遗传进化树(图2)显示,Dk/GX/038/09与广东分离的多个鸭源H6N6毒株如A/duck/Shantou/8365/2005(H6N6)和 H6N2 毒株如 A/duck/Shantou/17887/2005(H6N2)等位于同一分支上,遗传距离很近;而与江苏、福建、香港以及台湾等其他地区分离的H6亚型流感毒株的遗传距离较远。
在 NA基因进化树上(图3)显示,Dk/GX/038/09与广东、福建分离的H6N6亚型流感毒株位于同一个进化分支内;与江苏、香港等地的H6亚型禽流感病毒分离株则位于不同的分支上,亲缘关系较远。
3 讨论
本实验室首次在广西自治区分离到H6N6亚型禽流感病毒并对其HA和NA基因进行了序列分析和绘制基因遗传进化树,初步明确了该病毒在广西的流行态势。由于该病毒的NA和HA基因序列与广东相应毒株最为接近,因而推测Dk/GX/038/09与广东分离的H6N6亚型流感毒株可能来自同一祖先病毒,这也许与广东和广西密切的家禽产品交易相关。江苏、香港、台湾等地区分离的H6毒株亲缘关系较远,则表明目前在中国流行的H6亚型禽流感病毒存在一定的地域差异性和基因遗传多样性。同时,顾敏等[7]报道,江苏分离的鸭源H6N6毒株 A/duck/Jiangsu/022/2009为基因重排病毒,8个基因比较复杂,但均属于欧亚系。而万春和等[8]也报道我国福建分离的另一株鸭源 H6N6毒株 A/Muscovy/duck/Fujian/FZ01/2008也为多亚型禽流感病毒基因重排产物。因此,Dk/GX/038/09也存在基因重排的可能性,但仍需要进一步证实其内部基因。
表面糖蛋白基因HA和NA之间的协调与病毒的增殖和复制密切相关,HA蛋白是禽流感病毒最主要的表面糖蛋白,而NA蛋白则在病毒的成熟和释放中起重要作用。这两个蛋白也是宿主免疫系统最主要的靶抗原[9]。HA基因的突变有可能引起抗原变异,使病毒能够逃避宿主免疫压力而获得选择优势[10]。本次研究发现,两广地区流行的H6N6病毒从2005年至2009年间其HA和NA基因核苷酸变异均较小,原因可能是H6N6亚型禽流感病毒相对于人禽流感病毒而言,所受的免疫压力小。因为该病毒主要感染鸭,而鸭的饲养模式则决定了鸭的存活时间较短,对H6N6亚型禽流感病毒有免疫力的鸭较少,使得病毒不需过多地改变自身来适应宿主。所以近几年来,流行于两广地区的H6N6亚型禽流感病毒基因进化没有明显加快的迹象,但新的基因重排H6N6亚型禽流感病毒增多的趋势可能存在。
广西处于中国温暖潮湿的华南地区,是多种侯鸟迁徙路线的必经之地,同时该地区水网密布,四季放养或混养,这一特殊的生态环境加上该地区独特的地理气候条件和人们的生活习惯等,使得候鸟、水禽、猪、人频繁接触并交叉循环感染,具备禽流感病毒生存与传播的天然条件,国外众多学者把中国南方称为“流感起源中心”[11]。因此,准确地了解和把握广西水禽群体中的禽流感病毒的多样性,对我国预防禽流感的发生是非常重要的。H6亚型禽流感病毒是我国南方地区家鸭中比较常见的亚型,因此,对H6亚型病毒进行跟踪监测可为预防和控制禽流感提供重要的流行病学信息。
[1]Hoffmann E,Stech J,Guan Y,et al.Universal primer set for the full-length amplification of all influenza A viruses[J].Arch Virol,2001,146:2275-2289.
[2]Fouchier R A M,Munster V,Wallensten A,et al.Characterization of a novel influenza a virus hemagglutinin subtype(H16)obtained from black-head gulls[J].J Virol,2005,79:2814-2822.
[3]Takada A,Kuboki N,Okazaki K,et al.Avirulent avian influenza virus as a vaccine strain against a potential human pandemic[J].J Virol,1999,73(10):8303-8307.
[4]Wang C W,Wang C H.Experimental selection of virus derivatives with variations in virulence from a single low-pathogenicity H6N1avian infiuenza virus field isolate[J].Avian Dis,2003,47:1416-1422.
[5]Richard J W,Peter R W,Scott L K,et al.Reassortment an interspecies transmission of North American H6N2influenza viruses[J].Virology,2002,295:44-53.
[6]Chin P S,Hoffmann E,Webby R ,et al.Molecular evolution of H6influenza viruses from poultry in Southeastern China prevalence of H6N1influenza viruses possessing seven AP Hong Kong P156P97(H5N1)-like genes in poultry[J].J Virol,2002,76(2):507-516.
[7]顾 敏,宋庆庆,李彦芳,等.1株H6N6亚型禽流感病毒A/duck/Jiangsu/022/2009的全基因测序及遗传进化分析[J].中国人兽共患病学报,2010,26(7):642-647.
[8]万春和,傅光华,程龙飞,等.番鸭源H6N6亚型禽流感病毒全基因组的分子特征[J].微生物学报,2010,50(9):1147-1154.
[9]Hatta M,Gao P,Halfmann P,et al.Molecular basis for high virulence of HongKong H5N1influenza A viruses[J].Science,2001,293(5536):1840-1842.
[10]Steinhauer D A.Structure and function of the hemagglution[A].In:Nicholson K G,Webster R G,Hay A J.Textbook of influenza[C].London:Black Well Science Ltd,1998:54-63.
[11]Webster R G,Bean W J,Gorman O T,et al.Evolution and ecology of influenza A viruses[J].Microbiol Rev,1992,56(1):152-179.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
科学大观园(2022年2期)2022-01-23 11:05:15
天津市教科院学报(2021年5期)2021-11-10 07:32:40
生物学通报(2021年9期)2021-07-01 03:24:44
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
江苏农业科学(2016年8期)2017-02-15 19:54:11
动物医学进展(2015年10期)2015-12-07 05:46:18
淮阴工学院学报(2014年5期)2014-09-10 09:15:56
特产研究(2014年4期)2014-04-10 12:54:12