
鳝藤等4种藤本植物耐盐性试验
2011-05-08 09:35:08杜月青刘建强陈立潮金水虎
浙江林业科技 2011年4期
杜月青,刘建强,陈立潮,金水虎
(浙江天泰园林建设有限公司,浙江 绍兴 312000)
鳝藤等4种藤本植物耐盐性试验
杜月青,刘建强,陈立潮,金水虎*
(浙江天泰园林建设有限公司,浙江 绍兴 312000)
摘要:以鳝藤(Anodendron affine)、山蒟(Piper hancei)、风藤(Piper kadsura)和厚藤(Ipomoea pes-caprae)为试材,研究在盐胁迫下4种藤本植物形态特征和生理生化指标的变化。结果表明:4种藤本植物的细胞膜透性均随盐胁迫时间的延长而升高,同时盐浓度越大的处理其细胞膜透性越高;山蒟、风藤和厚藤的POD活性随盐浓度的升高而升高,鳝藤的POD活性基本随盐浓度的升高和处理时间的延长而逐渐下降;风藤、鳝藤和厚藤的游离脯氨酸随盐胁迫时间的延长而升高,山蒟的游离脯氨酸随盐胁迫时间的延长呈先升后降的趋势。4种藤本植物的可溶性蛋白含量基本随盐胁迫浓度的升高而呈先升高然后迅速下降并趋于稳定的趋势,并且盐胁迫时间越长其可溶性蛋白含量越低;4种植物的MDA含量随盐浓度的上升而增加,处理时间上以处理20 d的MDA含量最高。
关键词:盐胁迫;生理生化指标;鳝藤;山蒟;风藤;厚藤
藤本植物垂直绿化已经成为我国园林生态景观建设的新热点。特别在上海市,垂直绿化己经获得成功。同时,垂直绿化在北京、深圳、南京等城市也有较快的发展,许多藤本植物在立体绿化中得到了较好的利用[1~2]。
我国东部大部分发达城市处在沿海地区,这些地区分布有大面积的滨海盐渍土。淡水资源缺乏、土壤盐碱化、植被覆盖率底、环境质量需要改善是这些城市发展的重要问题。另外,在我国的一些内陆地区,尤其是东北、西北地区,由于对水土、草原、森林等资源的过度开发利用,致使某些地区沙漠化、盐碱化问题特别严重,引种耐盐碱植物,恢复地被成为这些地区的当务之急[3~5]。
1 材料与方法
1.1 材料与处理
试验材料为鳝藤(Anodendron affine)、山蒟(Piper hancei)、风藤(Piper kadsura)和厚藤(Ipomoea pes -caprae)的一年生扦插苗和实生苗。用20 cm×20 cm底部有孔的营养钵,下面设托盘,装入湿度和重量基本一致的基质(基质为珍珠岩、泥炭和蛭石按2:3:1的比例配制),选择生长良好、长势基本一致的植株进行移栽;每种藤本植物选择15盆,置于智能温室内缓苗一个月后开始进行盐胁迫处理,处理盐(NaCl)浓度设定为:0、0.3%、0.6%、0.9%和1.2%,每隔4 d定时定量浇灌一定浓度的盐溶液;于第5天、第10天、第15天和第20天分别测定相关生理生化指标。试验重复3次。按时均匀取样,叶片剪后置于冰袋中,样品均当天处理分析完毕。
1.2 方法
电导仪法测定细胞膜透性、愈疮木酚比色法测定过氧化物酶(POD)活性、考马斯亮蓝G-250染色法测定可溶性蛋白含量、茚三酮染色法测定游离脯氨酸含量、硫代巴比妥酸法测定丙二醛含量[6]。
试验数据用Excel2003和SAS9.0进行分析,并进行差异性比较,采用模糊数学的隶属函数法综合评价4种藤本植物的耐盐能力。
2 结果与分析
2.1 盐胁迫对4种藤本植物细胞膜透性的影响
细胞受盐害后,细胞膜遭受伤害,透性变大。大量研究表明,植物在逆境胁迫或衰老过程中,细胞内活性氧代谢的平衡被破坏而有利于自由基的超量积累,过剩自由基的毒害之一就是引发或加剧膜脂过氧化作用,造成细胞膜系统和代谢过程的损伤。在盐分胁迫下,相对电导率增幅小,表明植物细胞膜系统结构受损程度低,从而其耐盐性就高[7~9]。
由图1、图2、图3、图4表明,4种藤本植物的相对电导率均随胁迫时间的延长而升高,同时盐浓度越大的处理其相对电导率也越高,到第20天时,山蒟、风藤、鳝藤和厚藤在不同盐浓度处理的相对电导率平均分别是ck的2.55倍、2.75倍、3.11倍、2.30倍。
采用多重比较分析可知:山蒟、风藤和厚藤之间差异不显著,鳝藤和其它植物之间差异极显著;4种植物的相对电导率均随盐浓度的提高和处理时间的延长而增加。
根据上述分析可知,4种藤本植物的耐盐能力强弱依次为:厚藤 > 山蒟 > 风藤 > 鳝藤。
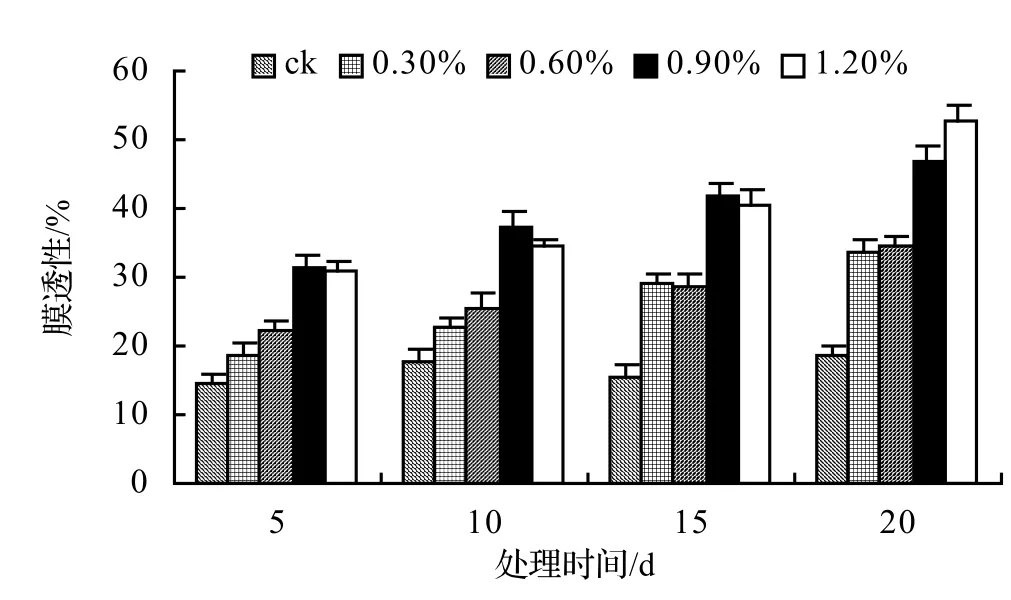
图1 盐胁迫对山蒟相对电导率的影响Figure 1 Effect of salt stress on relative conductivity of P. hancei

图2 盐胁迫对风藤相对电导率的影响Figure 2 Effect of salt stress on relative conductivity of P. kadsura
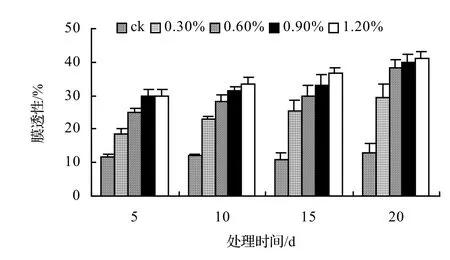
图3 盐胁迫对鳝藤相对电导率的影响Figure 3 Effect of salt stress on relative conductivity of A. affine

图4 盐胁迫对厚藤相对电导率的影响Figure 4 Effect of salt stress on relative conductivity of I. pes-caprae
2.2 盐胁迫对4种藤本植物POD活性的影响
在盐胁迫下POD活性变化目前尚无定论,盐胁迫后POD活性表现出既有升高也有降低,并且耐盐品种比盐敏感品种有较高的POD活性[10],还有一些植物体内POD活性随盐浓度的增大而增强[11~12];而有些植物在不同浓度盐胁迫下POD活性随盐浓度增加呈波动性变化[13]。
由图5、图6、图7、图8表明,山蒟、风藤和厚藤的POD活性随盐浓度的升高而升高,并且在胁迫处理第10天时达到最大值,然后迅速下降;鳝藤的POD活性基本随盐浓度的升高和处理时间的延长而逐渐下降。处理第10天时,山蒟在0.3%、0.6%和1.2%盐胁迫浓度下的POD活性达到最大值,其中1.2%盐浓度下其POD活性是ck的4.90倍;风藤在盐胁迫处理第5天时的POD活性迅速上升,0.3%、0.6%和1.2%盐浓度处理在第10天时达到最大值,其中1.2%盐浓度下其POD活性是ck的4.89倍;鳝藤在0.3%盐浓度处理第5天时达最大值,随后迅速下降,到第20天时下降了72.44%,1.2%盐浓度处理在第5天时达最大值,随后迅速下降,到第20天时下降了66.66%;厚藤4个盐浓度处理在第5天时同时达到最大值,盐浓度越高其POD活性也越高,随后迅速下降。
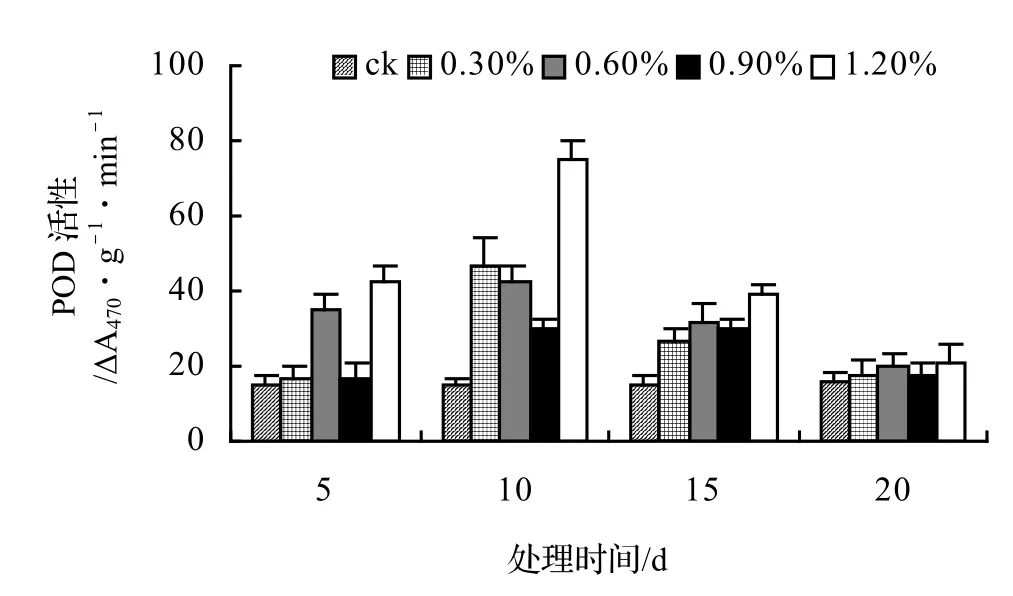
图5 盐胁迫对山蒟POD活性的影响Figure 5 Effect of salt stress on POD of P. hancei
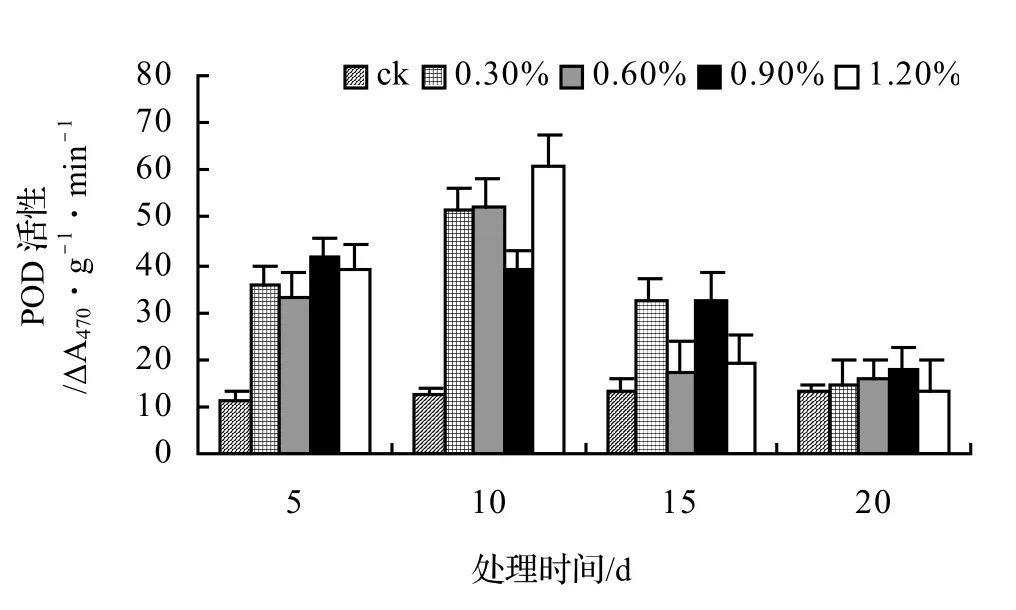
图6 盐胁迫对风藤POD活性的影响Figure 6 Effect of salt stress on POD of P. kadsura

图7 盐胁迫对鳝藤POD活性的影响Figure 7 Effect of salt stress on POD of A. affine

图8 盐胁迫对厚藤POD活性的影响Figure 8 Effect of salt stress on POD of I. pes-caprae
采用多重比较分析可知:山蒟和风藤之间差异不显著,厚藤和鳝藤之间差异显著,其中厚藤具有较高的POD活性,风藤最低;POD活性随盐浓度的升高而升高,0.3%和0.6%盐浓度之间差异不显著,0.9%和1.2%盐浓度之间差异不显著。
根据上述分析可知,4种藤本植物的耐盐能力强弱依次为:厚藤 > 鳝藤 > 山蒟 > 风藤。
2.3 盐胁迫对4种藤本植物游离脯氨酸含量的影响
目前把游离脯氨酸积累的作用大致归结为以下几个方面:可以作为细胞的有效调节物质[14];具有保护酶和膜的作用[15];可以直接利用的无毒形式的氮源,作为能量和呼吸底物,参与叶绿素的合成等,从脯氨酸在逆境条件下积累的途径来看,它既可能有适应性的意义,又可能是细胞结构和功能受损伤的表现,是一种伤害反应[16~17]。
由图9、图10、图11、图12表明,风藤、鳝藤和厚藤的游离脯氨酸随盐胁迫时间的延长而升高,山蒟的游离脯氨酸随盐胁迫时间的延长而呈现出先升高后降低的趋势。山蒟在各盐浓度处理至第15天时其游离脯氨酸含量达到最大值,其中0.3%为最高值,是ck的3.15倍,风藤游离脯氨酸在1.2%盐浓度处理至第20天时其游离脯氨酸含量为各处理中的最大值,是ck的3.07倍,鳝藤在盐胁迫初期各盐浓度之间差异不显著,到第20天时,其游离脯氨酸含量迅速增加,其中1.2%盐浓度处理为最高值,是ck的3.61倍,厚藤游离脯氨酸含量随盐胁迫的时间延长而升高,到第20天时,0.6%盐浓度处理达到最高值,是ck的3.76倍。
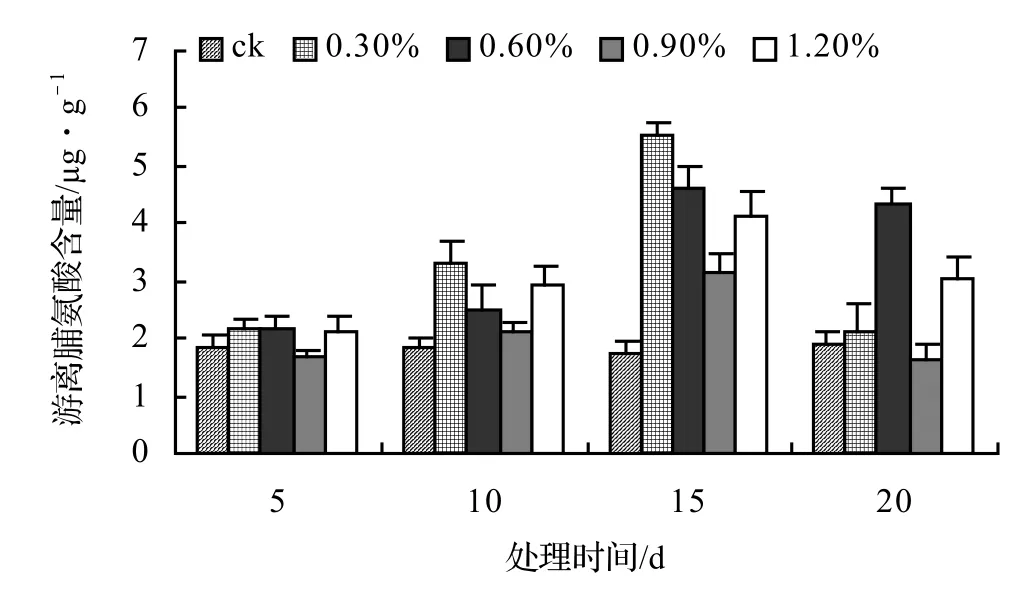
图9 盐胁迫对山蒟游离脯氨酸含量的影响Figure 9 Effect of salt stress on free proline of P. hancei
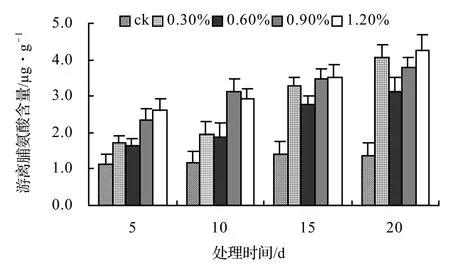
图10 盐胁迫对风藤游离脯氨酸含量的影响Figure 10 Effect of salt stress on free proline of P. kadsura
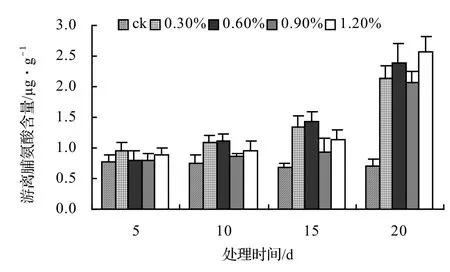
图11 盐胁迫对鳝藤游离脯氨酸含量的影响Figure 11 Effect of salt stress on free proline of A. affine

图12 盐胁迫对厚藤游离脯氨酸含量的影响Figure 12 Effect of salt stress on free proline of I. pes-caprae
采用多重比较分析可知:0.3%和0.6%盐浓度处理之间差异不显著,0.9%处理和其它处理之间差异极显著,各盐浓度处理之间以1.2%盐浓度处理最高,0.9%最低。
根据上述分析可知,4种藤本植物的耐盐能力强弱依次为:厚藤 > 鳝藤 > 山蒟 > 风藤。
2.4盐胁迫对4种藤本植物可溶性蛋白含量的影响
逆境下植物可溶性蛋白的变化与植物的抗逆性有一定的关系。许多资料表明,在盐胁迫下,植物体内的可溶性蛋白含量随着盐浓度的增加呈下降趋势[18]。徐云岭却发现可溶性蛋白并不随盐处理浓度的增大而呈直线变化;可溶性蛋白在不同的树种、不同发育阶段起的作用是不同的[19]。
由图13、图14、图15、图16表明,4种藤本植物的可溶性蛋白含量基本上随盐胁迫浓度的升高而呈现出先升高后下降并趋于稳定的趋势,并且盐胁迫时间越长其可溶性蛋白含量也越低,在后期其不同盐浓度之间表现不显著。山蒟在第10天时达到最大值,随后开始下降,到第20天时各盐浓度处理平均比ck下降43.53%;风藤在第10天时为最大值,随后便迅速下降,到第20天时各盐浓度处理平均比ck下降43.59%;鳝藤在第10天时为最大值,随后便迅速下降,到第20天时各盐浓度处理平均比ck下降39.29%;厚藤在第10天时为最大值,随后便迅速下降,到第20天时各盐浓度处理平均比ck下降37.06%。
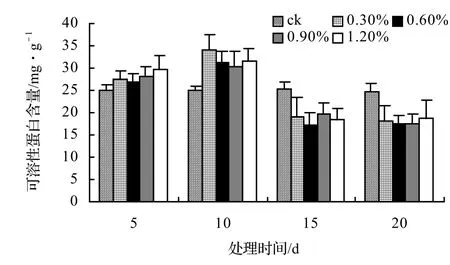
图13 盐胁迫对山蒟可溶性蛋白含量的影响Figure 13 Effect of salt stress on soluble protein conten of P. hancei
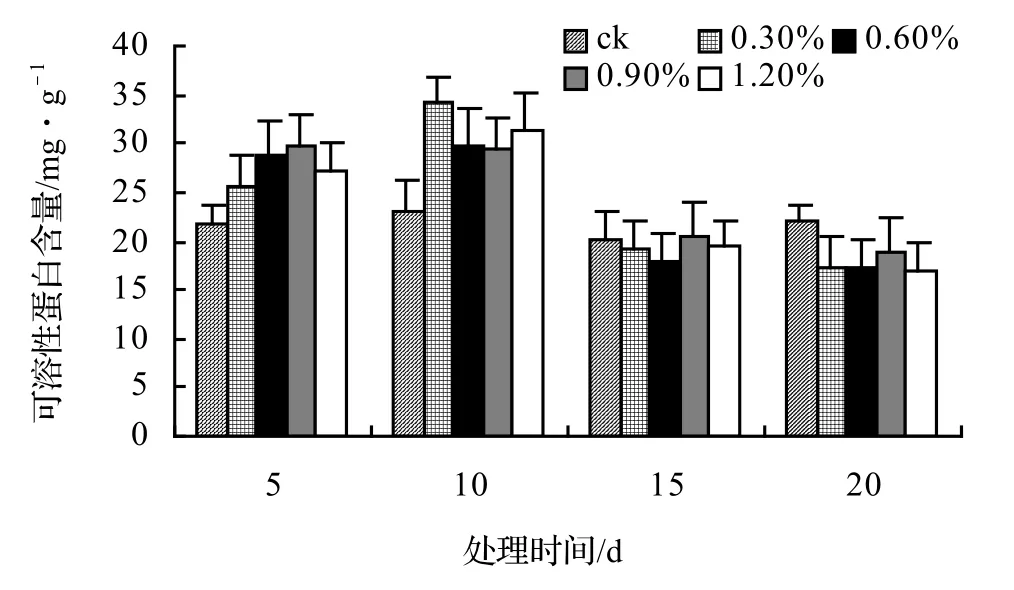
图14 盐胁迫对风藤可溶性蛋白含量的影响Figure 14 Effect of salt stress on soluble protein conten of P. kadsura

图15 盐胁迫对鳝藤可溶性蛋白含量的影响Figure 15 Effect of salt stress on soluble protein conten of A. affine

图16 盐胁迫对厚藤可溶性蛋白含量的影响Figure 16 Effect of salt stress on soluble protein conten of I. pes-caprae
采用多重比较分析可知:风藤、鳝藤和山蒟、风藤之间差异不显著,山蒟和厚藤之间差异极显著;各盐浓度之间差异不显著,可溶性蛋白随盐浓度的升高而降低;盐胁迫处理第10天达到最大值,第5天和第10天、15天之间差异极显著,第15天和第20天之间差异不显著。
根据上述分析可知,4种藤本植物的耐盐能力强弱依次为:厚藤>鳝藤>山蒟>风藤。
丙二醛(MDA)被认为是膜脂过氧化的产物,其含量与细胞膜系统的伤害程度密切相关[20~21]。随着盐胁迫强度的加大,MDA含量逐渐升高,MDA的含量大小可衡量膜质过氧化程度。在盐分胁迫下,MDA含量增幅小,表明植物抗氧化胁迫的能力强,从而其耐盐性就强。
由图17可知:山蒟的MDA含量在盐胁迫第5天时达到最大值,随后便开始下降,各盐胁迫处理的平均MDA含量约是ck的1.78倍。由图18可知:风藤的平均MDA含量约是ck的1.45倍。由图19可知:鳝藤各处理的平均MDA含量约是ck的1.19倍。由图20可知:厚藤各处理的平均MDA含量约是ck的1.17倍。
采用多重比较分析可知:不同植物之间差异极显著,其MDA总量大小为:风藤 > 鳝藤 > 山蒟 > 厚藤;其中0.6%和0.9%盐浓度处理之间差异不显著,0.3%和1.2%盐浓度处理之间差异极显著;处理时间以处理第20
2.5 盐胁迫对4种藤本植物丙二醛含量的影响天的MDA含量最高,第15天最小,第10天和第20天之间差异不显著,第5天和第15天差异不显著。
根据上述分析可知,4种藤本植物的耐盐能力强弱依次为:厚藤 > 鳝藤 > 风藤 > 山蒟。
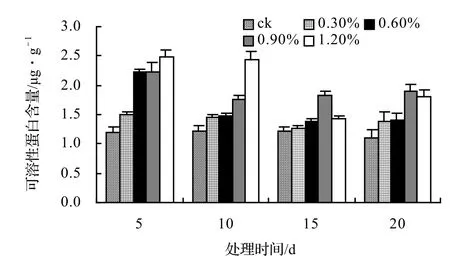
图17 盐胁迫对山蒟丙二醛含量的影响Figure 17 Effect of salt stress on MDA conten of P. hancei

图18 盐胁迫对风藤丙二醛含量的影响Figure 18 Effect of salt stress on MDA conten of P. kadsura

图19 盐胁迫对鳝藤丙二醛含量的影响Figure 19 Effect of salt stress on MDA conten of A. affine
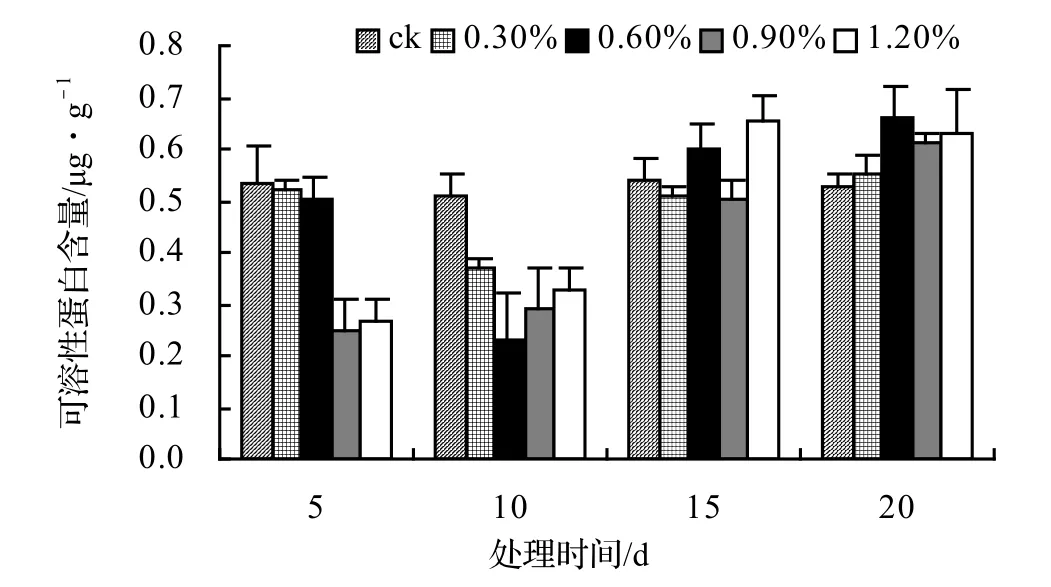
图20 盐胁迫对厚藤丙二醛含量的影响Figure 20 Effect of salt stress on MDA conten of I. pes-caprae
2.6 4种藤本植物耐盐性综合评价
植物的耐盐性是一个受多种因素影响的较为复杂的综合性状,多种因素的综合作用才促进耐盐性的形成,因此对于植物耐盐性的评价应该用尽可能多的指标来综合评价,从而弥补与缓和单个指标对于评定植物耐盐性所造成的片面性。本文选取细胞膜透性、POD活性、游离脯氨酸含量、丙二醛含量、可溶性蛋白含量5个指标,采用模糊数学的隶属函数法对4种藤本植物的耐盐性进行综合评价[22~25]。
由表1可知,4种藤本植物的耐盐能力大小依次为:厚藤 > 鳝藤 > 山蒟 > 风藤。
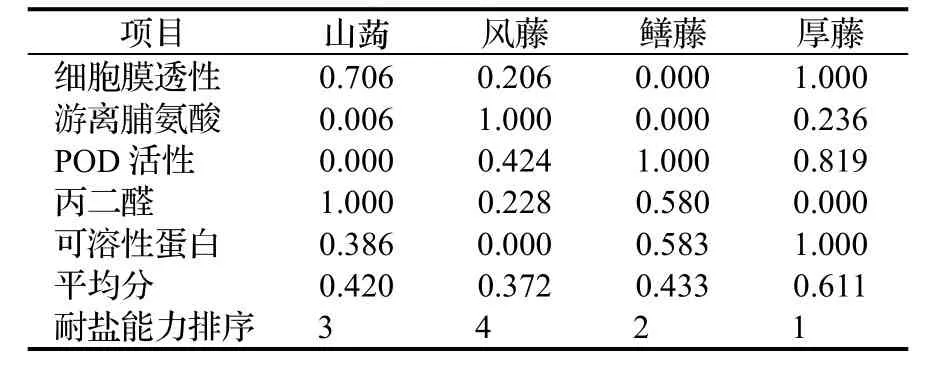
表1 4种藤本植物耐盐能力综合评价(隶属函数法)Table 1 Integrated evaluation of salt-resistance of 4 tested plants (by subordinate function)
3 结论与讨论
植物在受到逆境胁迫时,其植株的水分、激素含量等因子会发生一系列的变化,外观表现为叶片发黄、萎蔫、脱落,严重时将致其死亡等特征,具体表现为体内的一些酶发生改变,影响植物体内的同化作用。不同植物在不同的阶段有着不同的反应,植物对逆境胁迫下的生理调整会因胁迫方式、强度不同而有所变化,即存在生理调整的多样性。通过测定其植物体内的激素含量的变化来了解植物体内受迫害程度,从而得知植物体内受逆境胁迫的最大能力,在植物配置时做到适地适栽,为其园林推广应用提供理论依据。
厚藤在野外主要生长在沙滩上,耐盐性较强,植株根系极深,可作海滩固沙或覆盖植物,其四季常绿,叶形奇特,生长势强,花果期较长,叶花果具有较高的观赏价值。鳝藤在野外主要生长在岩石边上,具有一定的耐盐能力,园林上可借助强大的攀援能力充分覆盖裸露的山体及墙面等;山蒟和风藤也具有一定的耐盐性,其四季常绿,枝叶繁茂,叶有辣味不宜感染病虫害,具气生根,可攀援与岩石、墙面、树干、篱笆上生长,其优良的攀援能力可以形成别具特色的立题景观。
参考文献:
[1] 唐丽. 论藤本植物的造景功能[J]. 林业调查规划,2006,2(1):150-153.
[2] 董延龙. 攀缘植物在垂直绿化中的应用[J]. 黑龙江农业科学,2007(2):63-64.
[3] 马兴旺. 干旱区沙漠化土地治理与保护性耕作[J]. 新疆农业科学,2004,41(3):138-142.
[4] 山仑,邓西平,康绍忠. 我国半干旱地区农业用水现状及发展方向[J]. 水利学报,2002(9):27-31.
[5] 郭然,王效科,欧阳志云,等. 中国土地沙漠化,水土流失和盐渍化原因和驱动力总体分析[J]. 自然资源学报,2004,19(l):119-127.
[7] 白玉娥,易津,谷安琳,等. 盐胁迫对几种禾本科牧草根系细胞膜透性的影响[J]. 干早地区资源与环境,2001,15(5):51-54.
[8] 孙国荣,阎秀峰. 盐胁迫对星星草幼苗保护酶系统的影响[J]. 草地学报,2001,9(l):34-38.
[9] 杨传平,焦喜才,刘文祥,等. 树木的细胞膜透性与抗盐性[J]. 东北林业大学学报,1997,25(l):1-3.
[10] Sreenivasulu N,Ramanjulu S,Ramachandra K K, et al. Total peroxidase activity and peroxidase isoforms as modified by salt stress in two
cultivars of fox-tail millet with differential salt tolerance[J]. Plant Sci,1999,141(1):1-9.
[11] 金赞敏,王长海,刘兆普. 盐胁迫对芦荟几项生理生化指标的影响[J]. 食品与发酵工业,2004,30(10):1-4.
[12]张云起,刘世琦,杨凤娟,等. 耐盐西瓜砧木筛选及其耐盐机理的研究[J]. 西北农业学报,2003,12(4):105-108.
[13] 卢静君,多立安,刘祥君. 盐胁迫下两草种SOD和POD及脯氨酸动态研究[J]. 植物研究,2004,24(1):115-119.
[14] Watad A A,Reinhold L,Lerner H R. Comparision between as table NaCl selected nicotianacell lieand wild type[J]. Plant Physiol,1983(73):624-632.
[15] 汤章城. 逆境条件下植物脯氨酸累积及其可能的意义[J]. 植物生理学通讯,1984(l):15-21.
[16]Hanson A D,Nelsen C E,Everson E H. Evalution of free proline ac-cumulation as an index of drought resistance using two contrasting barley cultivars[J]. Crop Sci,1977(17):720-734.
[17] Stewar C R,Lee J A. Therole of proline accumulation ihalophytes[J]. Planta,1974,120(3):279-289.
[18]郭房庆,周建明,汤章城. NaCl胁迫下小麦突变体和野生型叶片中一些有机溶质积累和基因表达差异[J]. 植物生理学报,1999,25(3):263-268.
[19] 徐云岭,余叔文. 植物盐胁迫蛋白[J]. 植物生理学通讯,1989(2):12-16.
[20]王爱国,邵从本. 丙二醛作为植物膜质过氧化指标的探讨[J]. 植物生理学通讯,1986(2):55-57.
[21] 贾娜尔·阿汗,杨春武,等. 盐生植物碱地肤对盐碱胁迫的生理响应特点[J]. 西北植物学报,2007,27(1):79-84.
[22] 高建社,刘玉媛,符毓秦,等. 不同种源青杨幼树的生长特性[J]. 浙江林学院学报,2004,21(l):11-118.
[23] 蒲光兰,周兰英,胡学华,等. 土坡干旱胁迫对杏树渗透调节物质的影响[J]. 北方园艺,2005(2):50-51.
[24] 杨敏生,裴保华,朱之梯. 水分胁迫下白杨无性系生理和生长的数量遗传分析[J]. 北京林业大学学报,1997,19(2):50-56.
[25] 郑书星,樊军锋,苏小华. 欧洲黑杨无性系抗旱性综合鉴定研究[J]. 西北林学院学报,2005,20(1):57-64.
中图分类号:S731.2
文献标识码:A
文章编号:1001-3776(2011)04-0051-07
收稿日期:2011-02-27;修回日期:2011-04-05
作者简介:杜月青(1968-),女,浙江绍兴人,工程师,从事园林绿化工作;*通讯作者。
Salt Stress on Four Species of Climbing Plants
DU Yue-qing,LIU Jian-Qiang,CHEN Li-Chao,JIN Shui-hu*
(Zhejiang Tiantai Landscaping Co.,Ltd, Shaoxing 312000, China)
Abstract:Experiments were conducted on salt stress on morphology, physiology and biochemistry changes ofAnodendron affine,Piper hancei,P. kadsuraandIpomoea pes-caprae. The results showed that cell membrane permeability of four tested species increased with stress duration and salt concentration. The POD ofP. hancei,P. kadsuraandI. pes-capraeincreased with salt concentration. POD ofA. affinedecreased with stress duration and salt concentration. Free proline content ofP. kadsura,A. affineandI. pes-capraeincreased with stress duration, that ofP. hanceiincreased first and then decreased with stress duration. The soluble protein content of four tested species increased first and then decreased. The MDA content of tested four species increased with stress duration, MDA content topped the highest after twenty days of salt stress.
Key words:salt stress; physiological and biochemical indexs;Anodendron affine;Piper hancei;P. kadsura;Ipomoea pes-caprae
猜你喜欢
江苏农业科学(2016年12期)2017-04-05 14:08:05
江苏农业科学(2016年8期)2017-02-15 20:17:01
江苏农业科学(2016年8期)2017-02-15 19:41:25
江苏农业科学(2016年8期)2017-02-15 19:41:06
山东农业科学(2016年12期)2017-01-21 14:20:16
中国科技博览(2016年1期)2016-04-25 07:47:28
江苏农业科学(2015年8期)2015-09-10 15:56:05
湖北农业科学(2015年15期)2015-09-09 22:21:54
安徽农学通报(2015年12期)2015-07-09 11:06:39
山东体育学院学报(2014年4期)2014-09-22 11:11:40
