
湘西南两江流域支流分流山地区维管束植物区系研究
2011-11-24 02:06彭珍宝旷柏根旷建军夏江林袁正科张灿明陆奇勇
浙江林业科技 2011年4期
彭珍宝,旷柏根*,旷建军,夏江林,袁正科,张灿明,陆奇勇
(1.湖南省南岳树木园,湖南 衡阳 421900;2.湖南省林业科学院,湖南 长沙 410004;3.湖南省通道县林业局,湖南 通道 418500)
长江流域沅江的渠水支流和珠江流域融江浔水支流以湖南省西南边陲通道侗族自治县境内的八斗坡为分水岭,在南、北两侧山地形成分流区[1]。浔水支流径流从北往南方向注入珠江,而渠水径流从南往北方向经沅江入洞庭湖后汇入长江。寒冷气流经沅江渠水由北向南、暖湿气流经珠江浔水由南向北输入该区,又由于分流山地处于云贵高原向南岭山地渡地带,地质构造部位特殊,对该区的植被产生重大影响。该区在森林分区上属中亚热带雪峰山南部低山丘陵森林亚区与三江流域北部中低山森林亚区的交接地带[2]。在中国植被分区上属中亚热带常绿阔叶林南部亚地带南岭山地,栲类、蕈树林区与三江流域山地,栲类、木荷林、石灰岩植被区的交接地带[3],显然该区植被处于一种复杂而特殊的地位。为了探讨该区植物区系的特殊性,我们在2003年对“龙底沟谷森林生态系统自然保护区自然资源研究”[4]、2008年“万佛山生物多样性”[5]和“通道县生物多样性”[6]考察研究的同时,调查研究了湘西南长江、珠江流域(下称两江流域)支流分流山地区维管束植物区系的特征,现总结如下。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于湖南省通道县境内,是湖南也是湘桂黔交接山地森林植被保存较好的地区,森林覆盖率达78%。地理位置 25° 52′ ~ 26° 29′ N,109° 26′ ~ 110° 1′ E,东邻城步八十里大南山,南连广西九万大山北部山地,西接云贵高原东缘延伸地带,北接雪峰山南部的青定山。调查包括珠江流域浔水支流北部甘溪、坪阳2乡和长江流域沅江渠水支流的木脚、临口、播阳3乡山地原生森林植被保护较好的地段。调查区涉及乡面积约600 km2,实际调查涉及的天然林区面积约100 km2,为中低山地貌。区内海拔1 607.7 m的牛皮界经海拔936 m的八斗坡至海拔1 230.0 m的传素山,以东西走向的山脊为分水岭,分水岭有海拔1 000 m以上山峰82座。分水岭以北(称渠水区),地势南高北低;分水岭以南(称浔水区)地表切割强烈,自北向南地势急剧下降,山高谷深,珠江流域浔水支流向北伸展至此,最低海拔(张里村洞雷)203 m,与牛皮界相差1 404.7 m。区内地层主要为震旦系、寒武系和白垩系地层。岩石以板岩、页岩和红色砂砾岩石为主。地带性土壤为红壤(亚类)、黄红壤(亚类)、黄壤和山地黄棕壤。气候属亚热带湿润季风气候。分水岭南浔水区(最南端洞雷)比岭北渠水区(县城海拔379.5 m)年均气温高1.8℃,为17.1℃;1月气温高出2.0℃,为7.2℃;日均温 < 0℃的天数少16 ~ 17 d,极端低温高4.2℃,为-3.1℃;≥10℃年活动积温高出100 ~ 400℃,为5 000 ~ 5 300℃;年均降水量低于渠水区100 ~ 300 mm,为1 300 mm[6]。
2.2 研究方法
在浔水区甘溪乡的恩科、张里和坪阳乡的新江、东江,渠水区木脚乡的溪上、凉江、临口乡的太平岩、江寨和独坡的三省坡7处原生性天然林(或天然次生林)植被保存较好的地段进行植被调查。分溪沟、沟谷、山脊、山坡和开阔地设置样线,进行实地调查与标本采集[7]。在线路调查的基础上,选择有代表性的典型地段设立标准地,结合群落测定作样方调查[8~9]。先后共调查线路25条,长度约72 500 m,设立样地147块,每个样方面积为1 000 m2(对于山脊和丹霞地貌小面积群落,视其面积大小设400 ~ 600 m2)。样地记录调查时间、地点、母岩、土层厚度、坡度、坡位、坡向,乔灌层种类、数量、树高、胸径(灌木为地径)、冠径及频度,草木层测多度、高度、投影盖度与频度。采集植物标本,室内进行标本分类与鉴定[9~11],编制植物名录,进行调查数据的统计与分析。
2 植物区系分析
2.1 植物种类组成
根据2 000多号野生植物标本结合登记的常见物种统计结果见表1。
从表1中可看出,①除去栽培植物,蕨类植物(按秦仁昌系统)分别占全国科、属、种的76.19%、48.26%和12.10%[12],裸子植物(按郑万钧系统)分别占全国的72.73%、41.67%和13.99%[6];被子植物(按哈钦松系统)分别占全国[13]的53.49%、29.93%和7.81%~ 7.72%。蕨类植物科、属、种(含变种)数大于蕨类植物丰富的 24° 59′ ~ 25°19′ N的贵州红水河谷季雨林区的30科67属178种[14],大巴山的38科92属339种[15],与峨眉山的46科 110属 425种相近[12]。种子植物大于南京的126科543属1 204种(含变种)[16],浙江乌岩岭保护区134科578属1 197种(含变种)[17]和临近的贵州雷公山的146科421属869种(含变种)[18]。可见,该区域是中国南方植物物种集中、丰富的地区之一。②蕨类植物以真蕨类占优势,区系中同时含有古老残遗的石松亚门、水韭亚门和楔叶蕨亚门等蕨类植物类群。种子植物以被子植物占优势,而被子植物又以双子叶植占优势。上述优势植物构成该区植物区系的主体。
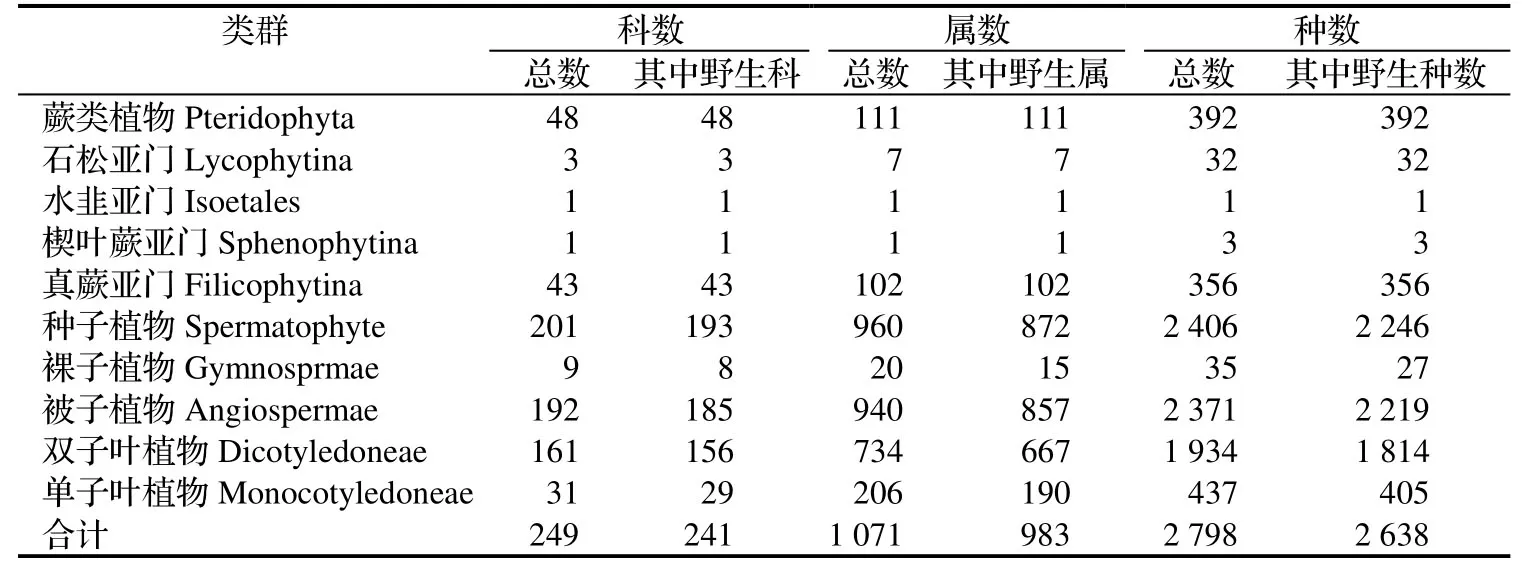
表1 湘西南两江流域支流分流山地区维管束植物类群统计Table1 Vascular plants in the southwest of Hunan
2.2 科、属大小统计分析
2.2.1 科的大小统计分析 根据研究区维管束植物科所含属、种的数量,将所含属的科分为5级,所含种的科分为6级,其不同类群在属、种等级中的分布列入表2和表3。
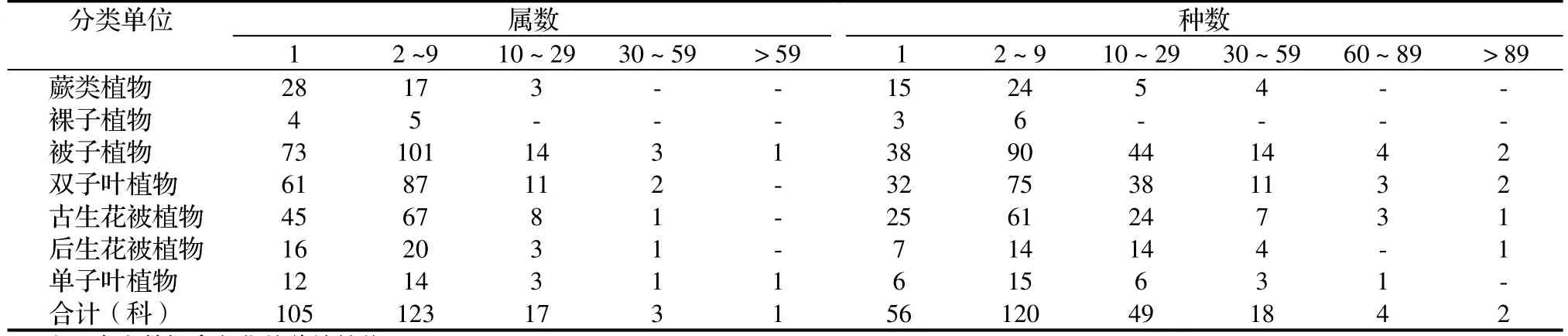
表2 湘西南两江流域支流分流山地区维管束植物科大小统计Table2 Vascular plants ordered by families

表3 湘西南两江流域支流分流山地区维管束植物属大小统计Table3 Vascular plants ordered by genera
由表2表明:①蕨类植物含10属以上的科有3科,含30种以上的科有4科,分别为蹄盖蕨科Athyriaceae(10属/39种,下同)、金星蕨科 Thelypteridaceae(13/36)、水龙骨科 Polypodiaceae(13/53)和鳞毛蕨科Dryopteridaceae(8/56)。这4个科占研究区科总数的8.33%,属总数的34.23%,占种总数的46.94%。裸子植物科均在9属9种以内,其中比较大的种为松科Pinaceae(3/9)、杉科Taxodiaceae(4/15)、柏科Cupressaceae(4/7)和红豆杉科Taxaceae(3/4),占研究区裸子植物科总数的44.44%,属总数的60.86%,种总数71.43%的种。②被子植物大于10属的有18个科,大于30种的有20个科,比较大的科有毛茛科Ranunculaceae(12/27)、葫芦科Cucurbitaceae(13/29)、山茶科Theaceae(11/53)、大戟科Euphorbiaceae(22/53)、蔷薇科Rosaceae(27/100),蝶形花科 Papilionaceae(40/89)、荨麻科 Urticaceae(12/32)、芸香科 Rutaceae(10/31)、五加科 Araliaceae(12/25)、茜草科 Rubiaceae(25/52)、菊科 Compositae(55/108)、玄参科 Scrophulariaceae(18/36)、唇形科Labiatae(25/41)、百合科Liliaceae(25/42)、天南星科Araceae(10/18)、兰科Orchidaceae(37/80)、莎草科 Cyperaceae(12/49)、禾本科 Gramineae(63/131)、樟科Lauraceae(9/61)、蓼科 Polygonaceae(5/35)、壳斗科Fagaceae(6/54)、桑科Moraceae(6/40)、冬青科Aquifoliaceae(1/41)和杜鹃花科Ericaceae(5/31),占有被子植物 13.47%的科,占有其 49.52%的属,53.66%的种,以上科是植物区系中占优势的科。③蕨类植物大于10种的属有卷柏属Selaginella(24种,下同)、凤尾蕨属Pteris(22)、铁角蕨属Asplenium(21)、鳞毛蕨属Dryopteris(24),蹄盖蕨属Athyrium(12)、复叶耳蕨属Arachniodes(13)6个属,5.41%的属占有29.59%的种;被子植物大于10种的属有含笑属Michelia(12)、山胡椒属Lindera(11)、新木姜子属Neolitsea(12)、铁线莲属Clematis(11)、堇菜属Viola(15)、蓼属Polygonum(29)、秋海棠属Begonia(10)、山茶属Camellia(16),柃木属Eurya(17)、猕猴桃属Actinidia(17)、大戟属Euphorbia(12)、野桐属Mallotus(10)、悬钩子属Rubus(36)、栲属Castanopsis(13)、青冈属Cycloblanopsis(15)、柯属Lithocarpus(12)、栎属Quercus(10)、榕属Ficus(27)、楼梯草属Elatostema(10)、冬青属Ilex(41)、卫矛属Euonymus(12)、蛇葡萄属Ampelopsis(10)、花椒属Zanthoxylum(10)、槭属Acer(20)、杜鹃花属Rhododendron(21)、紫金牛属Ardisia(15)、山矾属Symplocos(25)、荚蒾属Viburnum(17)、蒿属Artemisia(11)、珍珠菜属Lysimachia(12)、薯蓣属Dioscorea(11)、苔草属Carex(18)等32属,3.40%的属占有22.27%的种,以上属植物种类组成中占优势。⑷比较大的类群出现于蕨类植物和被子植物。在被子植物中双子叶植物的较大类群占优势,单子叶植物只有1科2属。双子叶植物中古生花被植物较大类群(19科24属)又多于后生花被植物(1科6属)。
2.3 属的分布区类型统计分析
根据吴征镒等2006年“种子植物分布区类型”划分原则[13],对研究区植物区系中872属野生种子植物属的分布区类型进行统计,结果见表4。
从表4中可看出:①野生种子植物属分布于14个分布区类型21个变型。如果以所占比例大小评价各分布类型与研究区联系的密切程度,则除世界广布型以外的 808属的各型的大小顺序为泛热带型(占总属数19.55%)、热带亚洲型(15.11%)、北温带型(占14.60%)、热带亚洲至热带非洲型(7.72%)、东亚分布型(12.62%)、热带亚洲至热带大洋洲(7.55%)、旧世界热带型(6.81%)、东亚及北美间断型(6.68%)、旧世界温带型(6.56%)、中国特有型(3.84%)、东亚(热带、亚热带)及热带南美间断型(2.73%)、地中海、西亚及中亚型(0.74%)、温带亚洲型(0.50%)。泛热带分布型以正型占优势,为该型总属数的82.32%;热带亚洲分布型以菲律宾、海南和台湾间断分布变型(7-5变型)占较大比例,为该型的56.59%,越南(或中南半岛)至华南或西南变型(7-4变型)次之,占15.56%;北温带分布型以北温带至南温带间断分布变型(8-4变型占50.82%)和正型(占42.41%)占优势;旧世界温带分布型以正型占较大比例(占58.45%),地中海、西亚(或中亚)和东南亚间断分布次之(10-1变型占22.68%);东亚型以正型占优势,为该型的 48.06%。②热带属440属,占除世界广布型以外属的54.52%,温带属337属,占41.76%,中国特有属31属,占3.84%,热带成分所占比例较大,R/T值为1.31, 与东面邻近的新宁崀 山风景名胜区(26° 15′ 06″ ~ 26° 24′ 21″ N,110° 42′ 53″ ~ 110° 49′ 34″ E)的 0.98[19]、西面邻近的雷公山自然保护区 0.86[18](26° 15′ ~ 26° 32′ N,108° 05′ ~ 108° 24′ E)、东南面的广东东八岭自然保护区1.18[20(]24° 2′ N,114° 0′ E)比较,分别高出33.67%、52.33%和11.02%,表现出较强的热带成分组成特性。③特有属31属,占除世界广布属以外属(下同)的3.84%,多于邻近新宁崀山的27属(占当地区系 3.59%)、舜皇山自然保护区[21](26° 15′ 06″ ~ 26° 55′ 22″ N,116° 28′ 53″ ~ 111° 18′ 34″ E)的25属(占3.65%)、广东东八岭的10属(占1.80%)、浙江清凉峰自然保护区(30° 10′ N,118° 56′ E)的19属(占 3.00%)[22]、贵州佛顶山自然保护区(27° 15′ ~ 27° 25′ N,107° 59′ ~ 108° 12′ E)的20属[23](占 4.10%)和贵州朱家山(26° 53′ 30″ ~ 27° 01′ 05″ N,107° 34′ ~ 107° 40′ E)的10属[24](占 2.55%),表现出特有属丰富而较集中的特性。研究区的中国特有属为:银杏属Ginkgo、观光木属Tsoongiodendron、血水草属Eomecon、泡果荠属Hilliella、堇叶芥属Neomartinella、石笔木属Tutcheria、藤山柳属Clematoclethra、棱果花属Barthea、虎颜花属Tigridiopalma、地构叶属Speranskia、半枫荷属Semiliqudambar、杜仲属Eucommia、青檀属Pteroceltis、枳壳属Poncirus、伞花木属Eurycorymbus、栾树属Koelreuleria、野鸭椿属Euscaphis、青钱柳属Cyclocarya、喜树属Camptotheca、通脱木属Tetrapanax、鸭头梨属Melliodendron、紫菊属Notoseris、虾须草属Sheareria、匙叶草属Latouchea、皿果草属Omphalotrigonotis、石山苣苔属Petrocodon、四轮香属Hanceola、白穗花属Speirantha、独花兰属Changnienia、井冈寒竹属Gelidocalamus和箬竹属Indocalamus31属,其中裸子植物1属,被子植物中的双子叶植物26属(古生花被亚纲19属,后生花被亚纲7属),单子叶植物4属。④种子植物不同类群与植物分布类型区和变型区的联系存在较大差异。野生裸子植物14属分布于6个分布型的3个正型与6个变型中,被子植物的双子叶植物出现于14个分布型21个变型中,单子叶植物出现于12个分布型15个变型中。

表4 湘西南两江流域支流分流山地区种子植物属分布区类型Table4 Distribution types of seed plant in the southwest Hunan province
2.4 种子植物区系中的单种属与寡种属统计分析
根据《中国种子植物分布区类型及其起源和分化》[13]中科、属的划分原则所确定的属中种数,统计出该区野生植物单种属59属,寡种属(2-6属)165属,分别占野生植物总属数的6.77%和18.92%。单种属中木本属26属,草本属33属。木本属中对应的乔木植物有福建柏Fokienia hodginsii、白豆杉Pseudotaxus chienii、观光木Tsoongiodendron odorum、山桐子Idesia polycarpa、任木Zenia insignis、杜仲Eucommia ulmoides、青檀Pteroceltis tatarinowii、伞花木Eurycorymbus cavaleriei、黄梨木Boniodendron minus、钟萼木Bretschneidera sinensis、银鹊树Tapiscia sinensis、喙核桃Annamocarya sinensis、青钱柳Cyclocarya paliurus、马尾树Rhoiptelea chiliantha、喜树Camptotheca acuminata、刺楸Kalopanax septemlobus、鸭头梨Melliodendron xylocarpum、鸡仔木Sinoadina racemosa18种;灌木有南天竹Nandina domestica、猫儿屎Decaisnea fargesii、舌柱麻Archiboehmeria atrata、野鸭椿Euscaphis japonica、通脱木Tetrapanax papyriferus5种;木质藤本有大血藤Sargentodoxa cuneata、藤山柳Clematoclethra lasioclada、飞龙掌血Toddalia asiatiea3种;以及草本属中的蕺菜Houttynia cordata、血水草Eomecon chionantha、虎颜花Tigridiopalma magnifica、葎草Humulopsis scandens、虾须草Sheareria nana、匙叶草Latouchea fokiensis、假婆婆纳Stimpsonia chamaedryoides、石山苣苔Petrocodon dealbatus、吉祥草Reineckea carnea、白穗花Speirantha gardenii等 33种,这些种是一些植物群落优势种、层外植物和林下植物的重要成分。单种属中大部分为古老残遗植物属,能反映出区系的古老残遗特性。寡种属中有木本属66属(乔灌木54属,藤本12属),草本属99属。木本属中有黄枝油杉Keteleeria calcarea、柳杉Cryptomeria fortunei、杉木Cunninghamia lanceolata、穗花杉Amentotaxus argotaeni、单性木兰Kmeria septentrionalis,乐东拟单性木兰Parakmeria lotungensis、粘木Ixonanthes chinensis、翅荚香槐Cleadrastis platycarpa、大果马蹄荷Exbucklandia tonkinensis、枫香Liquidumbar formosana、檵木Loropetalum chinensis、半枫荷Semiliquidambar cathayensis、糙叶树Aphananthe aspera、榉树Zelkova scheideriana、红椿Toona ciliata、黄杞Engelhardtia roxburghiana、青荚叶Helwingia japonica、复羽叶栾树Koelreuteria bipinnata、拟赤杨Alniphyllum fortunei、银钟花Halesia macgregorii、白辛树Pterostyrax psilophyllus等,是该区植物群落乔灌层的重要成分,有些种还为群落建群种。藤本属中细圆藤Pericampylus glaucus、紫藤Wisteria sinensis、粉背雷公藤Tripterygium hypoglaucum、定心藤Mappianthus iodoides、大花帘子藤Pottsia grandiflora、华萝藦Metaplexis hemsleyana、凌霄Campsis grandiflora、雷公莲Amydrium sinense等是森林中层外植物的重要成分,有些种还可在丹霞地貌崖壁下部构建藤本群落[25]。草本植物属中的裂叶星果草Asteropyrum caraleriei、三白草Saururus chinensis、草珊瑚Sarcandra glabra、假升麻Aruncus sivester、蛇莓Duchesnea indica、水蛇麻Fatora pilosa、微柱麻Chamabainia cuspidata、鸭儿芹Cryprotaemia japonicum、艾纳香Blumea balsamifera、石胡荽Centipeda minima、皿果草Omphalotrigonotis cuculifera、盾果草Thyrocarpus sampsonii、散斑竹根七Disporopsis aspera等为森林群落中易见成分。还有一些寡种属草本为山间沼泽和溪流湿地植物成分。寡种属也有许多古老残遗植物属。单种属和寡种属植物对于确定该区植物区系的古老残遗特性和森林植被的组成具有重要意义。
59个单种属的分布型中,15型有21属,占35.59%;14型18属,占30.51%,其中全型5属,14SH变型2属,14SJ变型11属;7型11属,占18.64%,其中7-4变型6属,7-5变型4属,7-2变型1属;10型3属,其中正型2属,10-1型1属;6型2属;5-1变型2属;1型3属。165个寡种属中,15型分布的有9属,占5.45%;14型36属,占21.82%,其中正型13属,14SH变型2属,14SJ变型21属;7型40属,占24.24%,其中正型2属,7-1型6属,7-2变型2属,7-3变型6属,7-4变型9属,7-5变型15属;9型20属,占12.12%,其中正型19属,9-1型1属;8型11属,占6.67%,其中正型7属,8-4变型3属,8-6变型1属;10型10属,占6.06%,其中正型5属,10-1变型4属,10-2变型1属;2型9属,占5.49%,其中正型6属,2-1变型3属;6型6属;5型5属;4型6属;3型1属;1型7属。统计中看出:①单种属中的中国特有属、东亚特有属和热带亚洲特有属占86.74%。表现出单种属的特有性,同时也说明,通过单种属的地理属性分析,可以反映区系成分的特有程度;②寡种属中的东亚分布、热带亚洲分布、东亚及北美间断分布、北温带分布和旧世界温带分布占70.91%。中国-日本(14SJ)、中国特有和东亚及北美间断分布成分是亚热带植物区系组成的特征成分[2]。寡种属的分布型属性能反映出该区植物区系的组成特点。
3 两江流域支流集水区植被特性的差异性分析
为分析两江流域支流分流区南北两支流发源地集水区植被特征的差异性,对1989年至2008年多次调查掌握的资料[4~5,29]对比分析后,将珠江浔水区和沅江渠水区的差异特征总结如表5。

表5 湘西南两流域支流两集水区植被特征差异Table5 Vegetation difference at tow watersheds in the southwest Hunan provnice
3.1 在区系种类组成上,两支流集水区山地存在较大的差异
这种差异主要表现在黔桂、华南成分对植物区系的影响程度上。在浔水支流集水区,黔桂、华南成分成为森林植物群落重要组成成分,有的还为群落优势种或建群种。典型的黔桂、华南成分小叶买麻藤Gnetum parvifolium、单性木兰Kmeria septentrionalis、黄果厚壳桂Cryptocarya concinna、臀形果Pygeum topengii、五列木Pentaphylax euryoides、海南树参Dendropanax hainanensis、多花山竹子Garcinia multiflora、观光木、东方古柯Erythroxylum kunthianum、红豆Ormosia hosiei、苍叶红豆Ormosia semicastrata、广东木瓜红Rehderodendron kwangtungense、天料木Homalium cochinchinense、亮叶猴耳环Pithecellobium lucidum、五层龙Salacia chinensis、子楝树Decaspermum gracilentum、密花树Rapanea neriifolia、石柑子Pothos cathcartii、黄梨木Boniodron minius、锦香草Phyllagathis caualeriei、翻白叶树Pterospermum heterophyllum、定心藤、追果藤Capparis uroplla、栗蕨Histiopteris incisa、红色新月蕨Pronephrium rubra、大羽鳞毛蕨Dryoptens wallichiana、中华双扇蕨Dipteris chinensis等,在渠水区甚至湖南其他地方少见或不见,为其分布北界。有海南五针松Pinus fenzeliana、白花含笑Michelia mediocris、乐东拟单性木兰、网脉山龙眼Helicia reticulata、广东琼楠Beilschmiedia fordii、黄果厚壳桂、大果铜锣桂Cryptocarya densiflora、华南桂Cinnamomum austro-sinense、华南木姜子Litsea greenmaniana、华南栲Castanopsis concinna、红钩栲C.lamontii、刺栲C.hystrix、黧蒴栲C.fissa、碟斗青冈Cyclobalanopsis disciformis、上思青冈Cy.delicatula、烟斗柯Lithocarpus corneus、愉柯L.amoenus、金毛柯L.chrysocomus、华南柯L.fenestratus、厚斗柯L.elizabethae、华南桦Betula austrosinensis、粗毛石笔木Tutcheria hirta、两粤黄檀Dalbergia benthami、华南云实Caesalpinia crista、半枫荷、白桂木Artocarpus hypargyreus、二色桂木A.styracifolus、九丁树Ficus nervosa,桂北木姜子Litsea subcoriacea、短柄山桂花Bennettiodendron brevipes、广西杜鹃Rhododendron kwangsiense、光枝杜鹃Rh.haoful、广西铜锤草Pratia wollastonii、黔稠Qutrcus stewadiana、西域青荚叶Helwingia himalaica、笔管榕F.wightiana、小果榕F.microcarpus、风车子Combretum alfredii、多花野牡丹Melastoma polyanthum、牛耳枫Daphniphyllum calycinum、九里香Murraya paniculata、飞龙掌血Toddalia asiatica、鸭脚罗伞Brassaiopsis glomerulata、拟罗伞树Ardisia depressa、当归藤Embelia parviflora、越南安息香Styrax tonkinensis、白花龙船花Ixora henryi。草本有海芋Alocasia macrorrhiza、野芭蕉Musa wilsonii、观音座莲Angiopteris fokiensis、金毛狗Cibotium barometz、华南紫萁Osmunda vachellii、鱼鳞蕨Acrophorus stipellatus、实蕨Bolbitis hainanensis、双盖蕨Dipiazium donianum、西南凤尾蕨Pteris wallichiana以及大量的黔桂、华南成分中的附生和石生草本植物等,成为浔水区森林植物群落中的重要组成,有些群落和优势种或建群种,这些种而在渠水区少见或不见,或零星分布,对森林植被结构影响很小。在渠水区,分布着一些典型的华中成分,像铁坚油杉Keteleeria davidiana、钟萼木、宜昌润楠Machilus ichangensis、假奓包叶Discoleidion rufescens、地瓜榕Ficus tikoua、红椿、柯楠Meliosma beaniana、巴东荚蒾Viburnum henryi、西川朴Celtis vandervoetiana、华榛Corylus chinensis、绢毛稠李Padus napaulensis、杜仲、猴樟Cinnamomum bodinieri、川桂C.wilsonii、爬藤榕Ficus sarmentosa、多花含笑Michelia floribunda、巴东栎Quercus engleriana、山羊角树Carrierea calycina、马桑Coriariasinica、荞麦叶大百合Cardiocrinum cathayanum、红柄木犀Osmanthus armatus等,是森林群落的重要成分,有的成为群落的优势种和建群种。这些种在浔水区少见或呈零星分布,对其森林植物群落的组成结构影响较小。
3.2 两集水区在植物群落类型组成上存在较大差异
浔水支流集水区由黔桂成分构建的竹柏群落Form.Nageia nagi、喙核桃群落、多花山竹子Garcinia multiflora群落、观光木群落、黧蒴栲群落、海南五针松群落,南岭山地成分构建的华南五针松群落、华南栲群落等乔林群落,以及沟谷中分布的金毛狗Cibotium barometz群落、福建观音座莲Angiopteris fokiensis群落、华南紫萁群落、海芋群落等小群落在渠水区不存在。而渠水区的亮叶水青冈Fagus lucido群落、水丝梨Sycopsis sinensis群落等浔水区不存在。
3.3 森林群落外貌存在很大差异
浔水区支流集水山地海拔 300 m左右的沟谷出现了与南亚热带季风常绿阔叶林外貌极为相似的沟谷雨林结构特征[4]:①乔木树干形成板根,在该区沟谷森林植物群落中极易见到,最大板根出现在甘溪恩戈沟谷的喙核桃下部,高达1.7 m,基部宽1.5 m,呈上窄下宽三角形状,一个树干有3块大型板根;②茎花、茎果在沟谷森林中易见,多见于桑科榕属的树干上;③大型叶和复叶植物增多,芭蕉Musa basjoo在该区能正常结果、成熟;④一些黔桂、华南成分,如白背鸭脚木Schefflera hypoleuca在湘南一带其他山地表现为矮小灌木,湘中湘北难见,在浔水支流集水区沟谷则表现为高大灌木或小乔木。上述特征在渠水支流集水区不见或难以见到。
根据上述分析,可以认为:①两江流域支流分流山地区具有错综复杂的地理成分,表现出区系组成成分较明显的热带属性及温、热带植物成分交汇特点;②浔水区和渠水区虽然以牛皮界经八斗坡至传素山的山脊相连,但在区系种类组成成分、植物群落类型组成和森林植物群落外貌结构上存在显著的差异,很难认定其是在同一植物区的共同特点。支持了祁承经等不宜将通道南部、黎平南部划归华中区范围的观点[26]。浔水区大量黔桂、华南成分的存在,可以归并为黔桂植物区。牛皮界经八斗坡至传素山可以作为两支流分流区的界线,这一界线也有可能向东西两侧延伸。
4 结论与讨论
(1)湘西南长江、珠江两流域渠水和浔水支流分流区有维管束植物249科1 071属2 798种,其中野生植物241科983属2 638种。野生植物中蕨类植物38科92属339种,裸子植物8科15属35种,被子植物185科857属2 219种。蕨类植物和被子植物科、属、种数明显大于邻近同纬度和近似纬度地区,在中国植物区系中占有较大的比例,该区是中国南方维管束植物集中丰富的地区之一。究其原因:①两流域支流分流区处于云贵高原向南岭山地、雪峰山脉西支向桂北山地过渡地带,环境因子变化很大,生境复杂多变,异质性增加。“环境异质性增加,环境复杂性增高,生物群落复杂,生物多样性就越高”[27];②处于华中、黔桂、华南植物区系的交汇地带,各区系植物种类的交互渗透,增加了该区物种多样性的丰富度;③对于流域生态系统而言,分流区处于珠江流域浔水支流生态系统和长江流域渠水支流生态系统两个系统边缘,边缘效应的多样性使物种丰富度增加。
(2)蕨类植物以真蕨类占优势,区系中含有一批古老残遗类群;被子植物以双子叶植物占优势。比较大的类群(10属或30种以上的科,10种以上的属)出现于蕨类植物和被子植物;被子植物中较大类群以双子叶植物占优势,较大类群的单子叶植物只有1科2属。双子叶植物中古生花被植物的较大类群(19科24属)又多于后生花被植物(1科6属)。
(3)872个野生种子植物属分布于14个分布型27个变型中。热带属占除世界广布种后总属数的54.52%,温带属占 41.78%,中国特有属占 3.84%。R(热带属)/T(温带属)值为 1.31,分别大于相似或较低于该区纬度东面相邻的新宁山风景名胜区、西面相邻的贵州雷公山自然保护区和东南面的车八岭自然保护区的33.67%、52.33%和11.02%。特有属及其所占总属数比例也明显高于相邻的相近似纬度地区,表现出区系较强的热带属性和特有种丰富而集中的特点。
(4)野生植物区系中有单种属59属,寡种属165属,分别占野生植物总属数的6.77%和18.92%。单种属中木本属26属(乔木18、灌木5、藤本3),草本属33属。寡种属中木本植物66属(乔灌木54,藤本12),草本99属。单种属和寡种属中的种类是该区一些植物群落的优势种、层外植物和林下植物的重要组成。单种属和寡种属能反映出区系的特有性、古老残遗性和亚热带地带性植被成分组成特性。
(5)渠水区和浔水区植物种类组成、植物群落类型和森林植物外貌结构上存在较大的差异。①黔桂、华南植物区的成分成为浔水区森林植物的重要组成成分,包括乔层、灌层、草层和层外植物,不少种类成为各层次的优势种。这些成分在渠水区不见和少见。在渠水区,分布着典型的华中成分,这些成分是该区森林植物成分的优势种或重要组成,但这些成分在浔水区少见或呈零星分布。②浔水区由黔桂、华南成分为优势种组成的一批群落在渠水区不见,而在渠水区高海拔区分布的有些群落在浔水区不见。③浔水区沟谷分布有相似于南亚热带或热带森林植物外貌相类似的沟谷雨林外貌:树干上板根常见,最大板根高达1.7 m、底宽1.5 m;茎花、茎果现象易见;大型叶和复叶种类增多,芭蕉能正常结果、成熟。这些外貌特征在渠水区不见,产生这些差异的原因:一是北部湾的暖湿气流沿珠江水系的融江直摇北上输入融江支流浔水北部的发源地,而沿洞庭湖水系沅江南下的寒冷气流经雪峰山阻挡削弱之后又被东西横亘的牛皮界至传素山拦截,削弱了对山脊南部浔水区的影响,使其成为湖南的“热窝子”[28],为黔桂成分和南岭山地、华南成分中喜热的种类在该区得到繁衍延续,使浔水区出现黔桂植物区的组成特性和植物群落分布的特点;二是特殊的地理位置成为多个植物区种类的汇集地区,也使两个不同支流发源地同时受到不同植物区系的影响,而出现植物区系种类的上述分布格局。
关于该区植物区的分区问题,祁承经曾经在研究“华中区植物区的特有种子植物”时,提出不宜将通道南部、黎平南部划归华中区范围的观点,但没有确定其划分界限。根据浔水区、渠水区植物区系组成、植物群落分布和森林植物群落外貌特征的差异性研究,可以以牛皮界经八斗坡至传素山向两侧地带作为通道南部华中植物区和黔桂区的分界线。
[1]邹锡求,刘应根.综合农业区域报告[A].湖南省通道县农业区划委员会.农业区域报告集[C].1984.1-56.
[2]蒋有绪,郭泉水,罗娟.中国森林群落分类及其群落学特征[M].北京:中国林业出版社,1998.5-9.
[3]吴征镒.中国植被[M].北京:科学出版社,1983.861-864.
[4]张海浪,刘国云,袁正科,等.湖南省通道县龙底沟谷森林生态系统自然保护区自然资源研究[M].长沙:湖南科学技术出版社,2003.9-20.
[5]袁正科,李星照,旷建军,等.万佛山丹霞地貌生物多样性调查研究报告[A].中南林业科技大学.万佛山生物多样性调查研究报告[C].2008.1-22.
[6]中国树木志编辑委员会.中国树木志(第一卷)[M].北京:中国林业出版社,1983.118.
[7]袁穗波,佘济云,付绍春,等.雪峰山西南支脉丘陵山地森林植物群落类型及结构特征研究[J].生态学杂志,2004,23(4):1-6.
[8]张金屯(译).生态学调查方法手册[M].北京:科学技术出版社,1999.127-140.
[9]祁承经.湖南树木志[M].长沙:湖南科学技术出版社,2002.1-719.
[10]祁承经,喻勋林.湖南种子植物总览[M].长沙:湖南科学出版社,2002.1-588.
[11]祁承经,孙希儒,林仕榕.湖南植物名录[M].长沙:湖南科学技术出版社,1986.1-32.
[12]谷海燕,李策宏.峨眉山蕨类植物区系的初步研究[J].西北植物学报,2008,28(2):381-387.
[13]吴征镒,周浙昆,孙航,等.种子植物的分布区类型及其起源和分布[M].昆明:云南科技出版社,2006.119.
[14]高焕晔,熊源新,苟光前.贵州红水河沟谷残存季雨林蕨类植物区系初步研究[J].浙江林学院学报,2008,25(1):48-54.
[15]许冬焱.大巴山自然保护区蕨类植物的区系分析[J].安微农业大学学报,2008,35(1):88-94.
[16]赵凯,张保卫,许远,等.南京种子植物区系的初步分析[J].安庆师范学院学报(自然科学版),2009,15(2):81-84.
[17]雷祖培,康华靖,张书润,等.乌岩岭国家级自然保护区种子植物区系的特征分析[J].武汉植物研究,2009,27(3):290-296.
[18]蓝开敏.雷公山自然保护区森林植物区系初步研究[A].周政贤.雷公山自然保护区科学考察集[C].贵阳:贵州人民出版社,1989.131-145.
[19]李星照,袁正科,彭珍宝,等.中国丹霞申报世界自然遗产提名地——湖南省新宁县崀山生物多样性考察与研究报告[R].湖南:中国林业科技大学,2008.
[20]陈锡沐,张常路,李秉滔.广东车八岭国家级自然保护区种子植物区系研究[J].广西植物,1994,14(4):321-333.
[21]罗仲春,刘小宁,伍石林,等.湖南舜皇山自然保护区植物区系考察报告[A].湖南舜皇山自然保护区综合科学考察报告集[C].北京:国家林业局中南林业规划设计院,2006.88-101.
[22]宋朝枢.浙江清凉峰自然保护区科学考察集[M].北京:中国林业出版社,1997.
[23]蓝开敏.佛顶山自然保护区种子植物区系研究[A].贵州佛顶山自然保护区科学考察集[C].北京:中国林业出版社,2000.42-72.
[24]蓝开敏.朱家山自然保护区种子植物区系研究[A].贵州省林业厅.贵州朱家山自然保护区科学考察集[C].北京:中国林业出版社,2000.
[25]彭珍宝,旷柏根,夏江林,等.崀山丹霞地貌区野生植物区系与生长特性研究[J].植物资源与环境学报,2009,18(4):58-65.
[26]祁承经,喻勋林,郑重,等.华中植物区的特有种子植物[J].中南林学院学报,1998,18(1):1-4.
[27]蔡晓明.生态系统生态学[M].北京:科学出版社,2000.122.
[28]祁承经,杨进鉴,梁绵业.通道县南部融江河谷盆地华南热带植物区系富集之乡[J].农业现代化研究,1983(2):127.
[29]袁正科,李星照,旷建军,等.通道侗族自治县生物多样性考察与研究综合报告[A].湖南省通道县侗族自治县生物多样性调查与研究报告[C].长沙:中南林业科技大学,1985.3-57.
猜你喜欢
数学大王·趣味逻辑(2022年8期)2022-07-10
医学概论(2022年8期)2022-06-19
世界热带农业信息(2019年11期)2019-01-05
天然产物研究与开发(2018年9期)2018-10-08
天然产物研究与开发(2018年9期)2018-10-08
小资CHIC!ELEGANCE(2018年24期)2018-08-13
网络空间安全(2016年3期)2016-06-15
天然产物研究与开发(2016年1期)2016-06-05
中成药(2016年4期)2016-05-17
湖北农业科学(2014年4期)2014-05-04
