
丛枝菌根真菌N代谢与C代谢研究进展
2011-01-11 12:36刘洁刘静金海如
微生物学杂志 2011年6期
刘洁,刘静,金海如
(浙江师范大学化学与生命科学学院,浙江金华321004)
丛枝菌根(arbuscular mycorrhizal,AM)真菌是一类内生菌根真菌,分布范围相当广泛,它可以与自然界普遍存在的植物根系建立共生体系从而促进宿主植物养分的吸收,并且可以增强植物的抗逆性,因此对植物生长有重大意义。AM真菌在促进宿主植物生长的同时,也完全依赖宿主植物供给自身生长所必需的碳水化合物[1]。AM真菌共生系统通常会增加植物的生物量,提高光合作用,并且对光合作用的部分产物有分配的作用[2]。AM真菌之所以能够促进植物生长主要是因为它能够促进宿主植物对矿质元素的吸收,包括磷、氮、硫、铜、锌等[3]。对AM真菌吸收、利用、转运P的机制已经研究的较透彻,尤其对在AM真菌吸收土壤中P的分子基础和基因调控方面取得了重要成果[4]。利用各种分子生物学、离体AM真菌培养、显微镜和核磁共振等各种技术,对AM真菌的C代谢和转运的机理研究有一定的进展[5]。但是对人们十分关注的AM 真菌对氮元素吸收、利用和转运机制研究的不十分明了。近期的研究主要针对氮素吸收的形态、共生体中氮转运和传递的形态、代谢途径做了大量的工作,并取得了一定的成果,进而从分子方面着手研究AM真菌的N代谢。本文主要是对AM真菌N代谢的研究进展,C和N代谢之间的关系做一个系统阐述。
1 AM真菌N元素吸收与植物氮素营养
AM真菌与植物建立共生体系,根外菌丝(extraradical mycelium,ERM)的生长扩大了根系的吸收面积,植物不仅由根吸收N源,也可以通过ERM从周围环境中吸收不同形态的N[6-7]。
一些研究表明,AM真菌转运无机N给植物[8]。土壤中主要无机N源的种类有-N,-N。是ERM吸收N的最主要形式,菌丝吸收后,以主动运输的形式输送给氮代谢途径,进行同化作用,但是要进入同化途径,必须消耗一定的能量,还原为才可以进入氮代谢途径[4]。在高等植物中,吸收时需要的能量来自于质膜两侧的电化学梯度,也就是通过2H+同向转移进行吸收
菌根真菌利用有机N源,例如外生菌根真菌利用氨基酸[10]和多肽类物质[11],欧石楠类菌根可以利用不易降解的有机物作为N源[12]。
土壤中的主要的氮形态还包括有机N源:氨基酸态氮、氨基糖态氮、腐殖质氨基酸[13]。谷氨酸、谷氨酰胺、天冬氨酸和丙氨酸都是土壤中最普遍的氨基酸,这些有机氮大部分是以复杂大分子的形式存在,不能被植物直接利用,但是AM真菌可以吸收利用这些有机氮源。利用15N标记技术发现,接种了Glomus fasciculatum的麦草可以吸收利用天冬氨酸和丝氨酸[14]。Heidi-Jayne Hawkins等[6]证明了不同的AM菌种对有机氮源的吸收速率也有所不同,不同的AM真菌与不同植物建立的不同共生体对各种N源的吸收能力也不相同。在Angela Hodge等[1]的试验中利用3室培养和15N、13C标记技术的方法,证明了AM真菌的菌丝优先生长到有有机材料的小室中,而不是先生长到未接种AM真菌的宿主植物室,且加快了有机材料的降解,证明了AM真菌可以促进有机物的降解,并且可以吸收转运有机物降解得到的N源,所以说AM真菌的共生体系具有腐生性营养的能力。利用AM真菌无菌2室培养系统,ERM培养基中分别加多肽Tri-Ala和牛血清蛋白BSA的研究证明了AM真菌还可以分解利用多肽和可溶性蛋白质[15]。
2 AM真菌与宿主植物间的N转运
通过同位素标记实验证明AM真菌吸收N源,并且可以将N转运给宿主植物[1,14,16]。
菌根真菌吸收N源后一般是将N整合入有机N载体—氨基酸,然后再进行运输。AM真菌与宿主植物建立共生体系中的N循环包括ERM吸收外源N,并且合成Arg[17],转运至根内菌丝(intraradical mycelium,IRM)[18],在IRM中Arg降解,分解出来的N以形式释放,可以作为N源整合入菌根内的其他氨基酸,也可以传递给宿主植物[19]。金海如等[4]报道了AM真菌氮代谢模型,见图1。
2 .1N源整合入精氨酸
AM真菌的ERM中富含氨基酸,其中主要的氨基酸包括天冬酰胺、精氨酸、谷氨酸、谷氨酰胺[20]。在众多氨基酸中,Manjula[19]和Cristina[21]通过向菌丝室供应15的方法培养共生体系发现了ERM中的大部分Arg被15N所标记,被15N标记Arg可占全部15N标记的游离氨基酸的90%,菌丝吸收的N大部分转化为Arg。
AM真菌的ERM吸收的外源氮要首先合成精氨酸。外生菌根真菌中ERM从土壤中吸收无机外源N,都转化为形式,通过AMT(NH+转运蛋白ammonium transpoters),以4的形式通过NADP依赖的谷氨酸脱氢酶(Glutmate dehydrogenase,GDH)途径或者谷氨酰胺合成酶-谷氨酸合酶(Glutamine synthetase-glutamate synthase,GS-GOGAT)途径进行N同化[22],ERM和IRM中N吸收关键酶的mRNA转录水平证明AM真菌中也存在GS-GOGAT的途径[19]。
在1985年,Smith等[22]就用酶法确认了谷氨酰胺合成酶的存在,并认为谷氨酰胺合成酶可能是AM真菌中N同化的关键酶。
研究报道G.intraradics的ERM中分离出谷氨酰胺合成酶,并对其部分cDNA进行了编码,谷氨酰胺合成酶活性受到不同N源的调节,其在AM真菌中也不总是表达,添加不同的N源,表达水平不同[23]。同样还对Glomus mosseae中谷氨酰胺合成酶的cDNA进行了全程编码[23]。Chunjie Tian[24]研究证明谷氨酰胺合成酶有2个亚型,他们具有不同的底物亲和力和表达模式,在N同化中起不同的作用。琥珀酸合成酶、精氨琥珀裂解酶和氨甲酰基磷酸合成酶参与了精氨酸的合成,并且也获得了这3种酶的序列。琥珀酸合成酶在ATP作用下催化瓜氨酸和天冬氨酸形成了精氨琥珀酸,琥珀酸合成酶有2个高度保守序列。精氨琥珀酸在琥珀酸裂解酶的作用下形成了精氨酸和延胡索酸。推测出的琥珀酸裂解酶849 bp长的部分序列已经扩增。
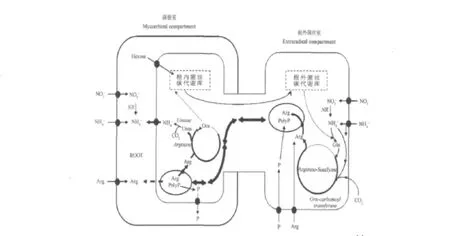
图1 AM真菌对外源氮的吸收、转运、分解代谢及传递的途径模型[4]Fig.1 A model on the pathway of external nitrogen uptake,translation,catabolism and transfer in arbuscular mycorrhizal symbiosis[4]
2.2 N素在菌丝内的转移
向ERM中提供13C标记的醋酸盐后,发现菌根组织中的标记游离氨基酸是在IRM中的[19]。精氨酸水平在侵染根中也比未侵染根中的比率高。与研究报道的精氨酸在ERM中快速转运一样,表明精氨酸可能从ERM中转运到IRM中[19]。
为了检测Arg是否从ERM中转运到IRM中,全部C分子都被13C标记的Arg添加到ERM中,培养6周以后ERM中34%的游离Arg和菌根中33%的游离Arg都有标记,通过质谱表示菌根中游离Arg要么未标记,要么所有的C分子都被标记。因此证明,Arg是从ERM中转运到IRM中的[19]。
Cristina测得N(以Arg为主的形式)能以3 nmol/(1h·mg fresh weight)的速度从ERM向IRM转移,其在菌丝间的转移速度接近于磷[21]。而目前还没有检测到菌丝间存在高效的Arg转运体,存储于液泡中的Arg如果在菌丝间不断跨膜运输必然消耗大量的能量及C,推测Arg可能和多聚磷酸盐(PolyP)协同运输,即P、N在菌丝内是同时转运的。另外,AM真菌具有特殊的无隔多细胞核结构,其液泡可能在菌丝间变形为管状,或通过原生质变形而形成管状液泡脉冲,这样Arg便能在此管状液泡中顺浓度梯度从ERM向IRM移动[25]。
精氨酸不仅仅可以由ERM向IRM中转运。Jin[26]利用13C同位素标记Arg证明了当ERM处于N匮乏阶段,并且菌根组织能从培养基质吸取大量的氮时,菌根组织以Arg的形式将氮运输给处于氮匮乏的ERM。由此证明AM真菌中的Arg可以在菌根和AM真菌中双向运转。
2.3 IRM中N素的转变
有研究报道缺乏外源N和添加L-戊氨酸(精氨酸合成的抑制因子)都会阻止ERM中精氨酸的合成,并且导致菌根中精氨酸酶和尿素酶的活性降低。证明了菌根中的尿素循环的分解代谢活动依靠ERM中转运来的精氨酸[21]。
N以Arg的形式转运到IRM中,设想如果以Arg的形式在宿主植物和AM真菌中传递,那么C也会跟随一起转移,但是实验表明,菌丝中的13C并没有随着Arg一起转移到宿主植物中,说明了N是以无机形态在真菌和宿主间转运的[19,27]。输送到IRM中的Arg通过精氨酸酶(arginase)和尿素酶(urease)的作用,通过鸟氨酸循环分解释放出NH4+和鸟氨酸,NH4+再通过AMT迅速转移给植物根细胞,而参与转运的C则留在IRM中,继续参加代谢[25]。Manjula发现,IRM中与精氨酸降解相关酶基因大量表达,明显比ERM高,AMT的基因也高效表达[19]。
精氨酸酶的活性受Arg诱导以及NH4+抑制,IRM中精氨酸酶的活性高,必然精氨酸的浓度高,NH4+的浓度低[21]。高浓度的精氨酸在精氨酸酶的催化下生成尿素和鸟氨酸。G.intraradics中的精氨酸酶与外生菌根真菌中分离得到的精氨酸酶具有60%的同源性。鸟氨酸转氨酶(GiOTA1)已经推论出有442个氨基酸序列与已经报道的GiOTA2的部分序列有所不同[28],利用互补实验研究GiOTA1,证明该基因功能是分解鸟氨酸[8],鸟氨酸脱羧酶是多聚胺生物合成途径的关键调节酶,并且可以催化鸟氨酸脱羧形成1,4-腐胺。缺乏尿素酶的酵母突变体,检测GiURE的功能是催化尿素水解为CO2和NH4+。在真菌、植物和细菌中的尿素酶都显示出很高的相似性。
3 AM真菌的C代谢与N代谢
C元素对AM真菌的生长具有相当重要的作用,C源可以促进孢子萌发,也可以促进菌丝生长[29],但是AM真菌自身不能进行光合作用,AM真菌与宿主植物互利共生,宿主植物可以将其光合作用的产物向下传递给AM真菌[30],宿主植物有4%~20%的光合作用产生的碳水化合物直接供给了菌根[31-32],然后碳水化合物分解释放出C素为AM真菌提供C源。
利用同位素标记和核磁共振光谱的方法研究菌根,发现ERM不能吸收葡萄糖、果糖。宿主植物光合作用合成的碳水化合物己糖,运送给IRM,共生体的IRM就可以吸收和利用己糖。己糖可以在IRM中转化为海藻糖和糖原,也可以通过戊糖磷酸途径进行代谢[33]。Saito等[34]之前就检测到戊糖磷酸途径中葡萄糖磷酸脱氢酶的活性。在IRM中也检测到几个糖酵解和三羧酸循环的酶的活性[34-35]。在番茄与Glomus mosse共生体的菌根中分析获得了磷酸甘油酸激酶的基因[36]。由己糖转化得到的糖原,也可以运送到ERM中去[2]。
宿主植物向AM真菌提供的C源是以脂质体三酰基甘油的形式储存在泡囊中,当植物的光合作用在14CO2中进行,或者提供14C标记的蔗糖、醋酸盐或者甘油,检测洋葱的菌根的脂质是有14C标记的[37]。三酰甘油是AM真菌中最大量的脂质[38-40],可以沿着菌丝由IRM向ERM运输[41]。在ERM中,通过乙醛酸循环,糖异生环途径将脂质转化为海藻糖或己糖[42],脂质也是ERM中三羧酸循环的主要呼吸底物[34]。
同样的研究发现ERM不能吸收外源碳(甘油、葡萄糖、果糖)为氨基酸提供碳骨架[33]。ERM中合成精氨酸及其他氨基酸的C骨架主要来源于内部C的存储及其降解[19,26]。有研究表明,提高空气中的CO2分压,促进AM真菌对N元素的吸收利用,提高宿主植物的N营养[43],添加C源甘油对ERM中精氨酸含量没有明显影响[26],但是在孢子萌发过程中添加葡萄糖,促进外源氮的吸收,产生的游离氨基酸是无葡萄糖添加的5倍[42],菌丝室施加葡萄糖和根浸出液提高了ERM干重和孢子数量,但是降低了ERM和菌根中Arg含量以及植株茎叶中N含量[44]。这一切都表明,AM真菌的C代谢和N代谢有着密切的联系,C、N代谢之间是有一定的调控关系。
4 展望
目前许多研究都已经证明AM真菌对植物的N营养作用,对C素的吸收的促进作用也有报道[33],AM真菌的N代谢和C代谢之间是有联系的,但是C、N代谢的调控关系还有待于进一步的研究。参与N代谢的关键酶及基因虽然获得[23-24],但各种酶之间的调控关系也有待进一步的了解。这将有助于更加完善AM真菌N代谢,以及C、N代谢之间的关系。
[1] Hodge A,Campbell C,Fitter A H.An arbuscular myeorrhizal fungus accelerates decomposition and acquisition nitrogen directly from organic materia1[J].Nature,2001,413:297-299.
[2] Bago B,Pfeffer P E,Shachar-Hill Y.Carbon metabolism and transport in arbuscular mycorrhizas[J].Plant Physiol,2000,124(3):949-958.
[3] Marschner H,Dell B.Nutrient uptake in mycorrhizal symbiosis[J].Plant Soil,1994,159(1):89-102.
[4] 金海如,蒋湘艳.AM真菌氮代谢与运转研究新进展[J].菌物学报,2009,28(3):466-471.
[5] Bago B,Shachar-Hill Y,Pfeffer P E.Dissecting carbon pathways in arbuscular mycorrhizas with NMR spectroscopy[C].GK Podila,DD Douds Jr.Current Advances in Mycorrhizae Research.APS Press St.Paul,2000:111-126.
[6] Hawkins H J,Johansen A,George E.Uptake and transport of organic and inorganic nitrogen by arbuscular mycorrhizal fungi[J].Plant and Soil,2000,226(2):275-285.
[7] Azcón R,Ruiz-Lozano J M,Rodriguez R.Differential contribution of arbuscular mycorrhizal fungi to plant nitrate uptake of15N under increasing N supply to the soil[J].Canadian Journal of Botany,2001,79(10):1175-1180.
[8] Hawkins H J,George E.Effect of plant nitrogen status on the contribution of arbuscular mycorrhizal hyphae to plant nitrogen uptake[J].Physiologia Plantarum,1999,105(4):694-700.
[9] McClure P R,Kochian L V,Spanswick R M,et al.Evidence for cotransport of nitrate and protons in maize roots.II.Measurement of NO3-and H+fuxes with lon-Selective Microelectrodes[J].Plant Physiol,1990,93(1):290-294.
[10] Lipson D A,Schadt C W,Schmidt S K,et al.Ectomycorrhizal transfer of amino acid-nitrogen to the alpine sedge Kobresia myosuroides[J].New Phytol,1999,142(1):163-167.
[11] Abuzinadah R A,Read D J.The role of proteins in the nitrogen nutrition of ectomycorrhizal plants.IV.The utilization of peptides by birch(Betula pendula L.)infected with different mycorrhizal fungi[J].New Phytol,1989,112(1):55-60.
[12] Read D J.The structure and function of the ericoid mycorrhizal root[J].Annals of Botany,1996,77(4):365-374.
[13] Azcón R,Rodríguez R,Amora-Lazcano E,et al.Uptake and metabolism of nitrate in mycorrhizal plants as affected by water availability and N concentration in soil[J].European Journal of Soil Science,2008,59(2):131-138.
[14] Cuenca G,Azc6n R.Effecfs of ammonium and nitrate on the growth of vesicular-arbuscular myeorrhizal Erythrina poeppigiana O.I.Cook seedlings[J].Biol Fertit Soil,1994,18(3):249-254.
[15] 金海如,张萍华,蒋冬花.同位素示踪研究丛枝菌根真菌吸收不同氮素并向寄主植物输运的机理[J].土壤学报,2011,48(4):888-892.
[16] Jean B C,Philip J M,Jean B.Effect of the arbuseular myeorrhizal fungus Glomus fascicalatum on the uptake of amino nitrogen by Lolium perenne[J].New Phytol,1997,137(2):345-349.
[17] Jin H R,Pfeffer P E,Douds D D,et al.The uptake,metabolism,transport and transfer of nitrogen in an arbuscular mycorrhizal symbiosis[J].New Phytologist,2005,168(3):687-696.
[18] Chen X M,Baumgartner K.Arbuscular mycorrhizal fungi-mediated nitrogen transfer from vineyard cover crops to grapevines[J].Biology and fertility of soil,2004,40(6):406-412.
[19] Govindarajulu M,Pfeffer P E,Jin H R,et al.Nitrogen transfer in the arbuscular mycorrhizal symbiosis[J].Nature,2005,(435):819-823.
[20] Bago B,Pfeffer P E,Douds D D,et al.Carbon metabolism in spores of the arbuscular mycorrhizal fungus Glomus intraradices as revealed by nuclear magnetic resonance spectroscopy[J].Plant Physiol,1999,121(1):263-271.
[21] Cristina C,Helge E,Carmen T,et al.Enzymatic evidence for the key role of arginine in nitrogen transloation by arbuseular myeorrhizal fungi[J].Plant Physiology,2007,144(2):782-792.
[22] Smith S E,John,B J St.,Smith F A,et al.Activity of glutamine synthetase and glutamate dehydrogenase in Trifolium subterraneum L.and Allium cepa L.:effects of mycorrhizal infection and phosphate nutrition[J].New Phytol,1985,99(2):211-227.
[23] Breuninger M,Trujillo C G,Serrano E,et al.Different nitrogen sources modulate activity but not expres sion of glutamine synthetase in arbuscular mycorrhizal fungi[J].Fungal Genet Biol,2004,41(5):542-552.
[24] Tian C J,Kasiborski B,Koul R,et al.Regulation of the Nitrogen Transfer Pathway in the Arbuscular Mycorrhizal Symbiosis:Gene Characterization and the Coordination of Expression with Nitrogen Flux[J].American Society of Plant Biologists,2010,153(3):1175-1187.
[25] 邓胤,申鸿,郭涛.丛枝菌根利用氮素研究进展[J].生态学报,2009,29(10):5627-5635.
[26] Jin H R.Arginine bi-directional translation and breakdown into ornithine along the arbuscular mycorrhizal mycelium[J].Science in China Series C:Life Sciences,2009,52(4):381-389.
[27]Fitter A H,Graves J D,Watkins N K,et al.Carbon transfer between plants and its control in networks of arbuseular mycorrhizas[J].Functional Ecology,1998,12(3):406-412.
[28] Gomez S K,Javot H,Deewatthanawong P,et al.Medicago truncatula and Glomus intraradices gene expression in cortical cells harboring arbuscules in the arbuscular mycorrhizal symbiosis[J].BMC Plant Biol,2009,(9):10.
[29] Bécard G,Piché Y.Fungal growth stimulation by CO2and root exudates in the vesicular-arbuscular mycorrhizal symbiosis[J].Appl Environ Microbiol,1989,55(9):2320-2325.
[30] 刘润进,李晓林.丛枝菌根及其应用[M].北京:科学出版社,2000.
[31] Douds DD,Pfeffer PE,Shachar-Hill Y.Carbon partitioning,cost and metabolism of arbuscular mycorrhizae in arbuscular mycorrhizas:physiology and function[C]//YKapulnick,DD Douds Jr.Arbuscular Mycorrhizas:Molecular Biology and Physiology.Kluwer Academic,Dordrecht,The Netherlands,2000:107-130.
[32] Graham JH.Assessing costs of arbuscular mycorrhizal symbiosis agroecosystems fungi[C].GK Podila,DD Douds Jr.Current Advances in Mycorrhizae Research.APS Press,St.Paul,2000:127-140.
[33] Pfeffer PE,Douds DD,Bécard G,et al.Carbon uptake and the metabolism and transport of lipids in and arbuscular mycorrhiza[J].Plant Physiol,1999 120(2):587-598.
[34] Saito M.Enzyme activities of the internal hyphae and germinated spores of an arbuscular mycorrhizal fungus,Gigaspora margarita Becker and Hall[J].New Phytol,1995,129(3):425-431.
[35] MacDonald R M,Lewis M.The occurrence of some acid phosphatases and dehydrogenases in the vesicular-arbuscular mycorrhizal fungus Glomus mosseae[J].New Phytol,1978,80(1):135-141.
[36] Harrier L A,Wright F,Hooker J E.Isolation of the 3-phosphoglycerate kinase gene of the arbuscular mycorrhizal fungus Glomus mosseae(Nicol.and Gerd.)Gerdemann and Trappe[J].Curr Genet,1998,34(5):386-392.
[37] Lösel D M,Cooper K M.Incorporation of14C-labeled substrates by uninfected and VA mycorrhizal roots of onions[J].New Phytol,1979,83(2):415-426.
[38] Cooper K M,Lösel D M.Lipid physiology of vesicular arbuscular mycorrhiza[J].New Phytol,1978,80(1):143-151.
[39] Beilby J P,Kidby D K.Biochemistry of ungerminated and germinated spores of the vesicular-arbuscular mycorrhizal fungus Glomus caledonium:changes in neutral and polar lipids[J].Journal of lipid research,1980,21(6):739-750.
[40] Gaspar L,Pollero R,Cabello M.Partial purification and characterization of a lipolytic enzyme from spores of the arbuscular mycorrhizal fungus Glomus versiforme[J].Mycologia,1997,89(4):610-614.
[41] Bago B,Pfeffer P E,Shachar-Hill Y.Could the urea cycle be translocating nitrogen in the arbuscular mycorrhizal symbiosis?[J].New Phytol,2001,149(1):4-8.
[42] 金海如,田萌萌.丛枝菌根真菌萌发孢子利用不同氮素及外源葡萄糖对其代谢的影响[J].中国科学:生命科学,2010,40(3):239-249.
[43] Gampere H,Hartwig U A,Leuchtrrmnn A.Mycorrhizas improve nitrogen nutrition of Trifolium repens after 8 yr of selection under elevated atmospheric CO2 partial pressure[J].New Phytol,2005,167(2):531-542.
[44] 田萌萌,吉春龙,刘洁,等.葡萄糖、根浸出液对丛枝菌根真菌吸收不同外源氮产生精氨酸的影响[J].微生物学通报,2011,38(1):14-20.
猜你喜欢
食品与生物技术学报(2021年4期)2021-01-17
园林科技(2020年2期)2020-01-18
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年1期)2016-11-12
中外医疗(2015年11期)2016-01-04
医学研究杂志(2015年12期)2015-06-10
中国当代医药(2015年30期)2015-03-01
中国果业信息(2014年2期)2014-01-23
植物营养与肥料学报(2011年6期)2011-10-24
植物营养与肥料学报(2011年6期)2011-10-24
