
UV-B辐射增强对麻花艽叶片光合作用及相关生理参数的影响
2010-09-23 02:13吴建国
环境科学研究 2010年11期
吴建国
中国环境科学研究院,北京 100012
UV-B辐射增强对麻花艽叶片光合作用及相关生理参数的影响
吴建国
中国环境科学研究院,北京 100012
采用20 W和40 W的UV-B灯辐射增强处理,测定分析了UV-B辐射增强对麻花艽(Gentiana stram inea Maxim)叶片净光合速率及相关生理参数的影响.结果显示:2008年8月3日,研究区大气温度从09:00开始上升,到14:00达到最高点,叶温和大气饱和水气压变化与气温变化趋势相似;光合有效辐射强度从07:00开始上升,至13:00达到最高,之后下降;麻花艽叶片净光合作用速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(E)和水分利用效率在对照下最高,在40 W UVB辐射下最低;Pn在08:00—10:00最高,Gs在07:00—08:00最高,Ci在日出及日落时最高,E在09:00—13:00及14:00—18:00最高,水分利用效率在日出后最高;Pn与Gs,叶片温度,大气温度,光合有效辐射和大气饱和水气压亏缺呈正相关,与 Ci呈负相关,与光合有效辐射强度的相关系数较高.Pn在光合有效辐射强度为0~800μmol/(m2·s)时随光合有效辐射强度增加而增加,在2 200~3 000μmol/(m2·s)时变化不大;Gs和E随光合有效辐射强度的增加而增加;Ci在光合有效辐射强度为0~800μmol/(m2·s)时,随光合有效辐射强度的增加而呈下降趋势;水分利用效率在光合有效辐射强度为 0~800 μmol/(m2·s)时随光合有效辐射强度的增加而呈增加的趋势,在800~3 000μmol/(m2·s)时呈下降趋势.说明UV-B辐射增强将使高寒草甸植物叶片Pn,Gs,Ci,E和水分利用效率降低.
UV-B辐射;高寒草甸;光合作用;麻花艽;青藏高原
Abstract:The effects of enhanced UV-B radiation on the laminae net photosynthetic rate and related physiological parameters of strawyellow gentian(Gentiana straminea Maxim)were determined and analyzed by using 20 W and 40 W UV-B lamps to increase the UVB radiation.The results showed that air temperature in the studied area began to rise from 09:00 am and peaked at 14:00.The variation trend of leaf temperature was similar with that of the atmospheric saturated vapor pressure and air temperature on August 3rd,2008.The photosynthetic active radiation(PAR)intensity began increasing at07:00 am,peaked at 13:00,and then decreased.The laminae net photosynthetic rate(Pn),stomata conductance(Gs),intercellular CO2concentration(Ci),transpiration rate(E)and water use efficiency(WUE)of straw-yellow gentian were the highest under the control treatment,and lowest under 40 W UV-B lamp treatment.Pnwas the highest from 08:00-10:00;Gswas the highest from 07:00-08:00;Ciwas the highest at time of sunrise or sunset;E was the highest at09:00-13:00 or 14:00-18:00;and,WUE was the highest after sunrise.Pnwas positively related with Gs,leaf temperature,atmosphere temperature,PAR or atmospheric saturated vapor pressure deficit,and negatively related with Ci.The correlation coefficient between Pnand PAR was higher.Additionally,the results showed that Pnincreased with increasing PAR when the PAR ranged from 0-800μmol/(m2·s),but it varied little when the PAR ranged from 2200-3000μmol/(m2·s).Gsand E increased with increasing PAR.Cidecreased with increasing PAR when the PAR ranged from 0-800μmol/(m2·s).WUE increased with increasing PAR when the PAR ranged from 0-800 μmol/(m2·s),but decreased when PAR ranged from 800-3000 μmol/(m2·s).The results indicated that enhanced UV-B radiation would result in decreases of Pn,Gs,Ci,E or WUE ofalpine meadows leaves.
Keywords:UV-B radiation;alpine meadows;photosynthesis;straw-yellow gentian;Qinghai-Tibet plateau
位于大气平流层的臭氧层能吸收对生物有害的太阳紫外辐射(UV-B辐射,280~320 nm),而大量氟氯烃化合物等的使用会引起臭氧层衰减[1].据联合国环境署(UNEP)[2]评估,与 1980年相比,1997—2000年南半球中纬度地区臭氧层每年减少6%,北半球中纬度地区冬春季臭氧层减少4%,夏秋季减少2%,全球年均减少约3%.自蒙特利尔公约执行以来,大气平流层中 O3消耗物浓度开始下降,但臭氧层厚度仍比20世纪70年代低,未来几十年内也难以很快恢复,如果对氯氟烃化合物排放量不加以限制,估计到 2060年臭氧层将减少 16%,2075年将比1985年减少40%.臭氧层破坏使地表UV-B辐射强度增加,大气中O3浓度每减少1%,地面生物有效紫外辐射强度将增加约2%.UV-B辐射强度自20世纪90年代在北半球持续增加,20世纪末超过其自然变化范围[3].UV-B辐射增强对地球上生物和生态系统功能及人类本身都构成严重威胁[4],揭示UV-B辐射强度对生态系统影响及制定有效的防护对策是目前面临的重大环境问题[3].
UV-B辐射强度变化对陆地生态系统影响在20世纪70年代引起广泛关注,随着对气候变化问题更广泛的关注,UV-B辐射强度变化对陆地生态系统影响问题在 90年代更成为研究热点.如FLINT等[5]分析发现UV-B辐射强度改变将改变植物生理生态过程;KAKANI等[6]发现UV-B辐射强度增加使植物光合作用发生极大改变; OUDEJANS等[7]发现UV-B辐射强度的增加使植物群落结构改变;PAOLETTI[8]总结了UV-B辐射强度增加对地中海森林植物的影响;SULLIVAN[9]总结了UV-B辐射强度的增加对北美树木的影响.在我国,UV-B辐射强度对陆地生态系统的影响研究在20世纪80—90年代也引起了关注,包括对植物生理生化属性和生理生态过程影响的研究[10-12],集中在农田作物或实验室短期生理过程研究,对高山植物也有研究[13],但主要分析了 UV-B辐射增强对植物光合作用及有关参数的短期影响.对田间条件下这些生理参数与其他环境要素的关系,以及不同光合辐射强度下 UV-B辐射增加对高寒草甸植物光合作用及相关生理参数影响考虑不够.
青藏高原是我国UV-B辐射强度最为强烈的地区,随着大气平流层臭氧层破坏的加剧,UV-B辐射强度进一步增强[13-16].高寒草甸是青藏高原典型生态系统,长期受到UV-B辐射变化的影响,目前的认识还很有限[17].系统地开展 UV-B辐射强度增加对高寒草甸植物叶片光合作用及相关生理参数日变化的影响,对科学认识UV-B辐射变化对高寒草甸生态系统影响及制定对策具有重要意义.为此,选择高寒草甸的典型组成植物——麻花艽(Gentiana straminea Maxim),通过田间UV-B辐射增强试验,测定分析了UV-B辐射强度变化对高寒草甸典型植物叶片净光合作用速率及相关生理参数的影响,以期为科学认识UV-B辐射强度变化对高寒草甸生态系统的影响提供依据,为进一步开展相关研究提供参考.
1 材料与方法
1.1 研究区概况
研究区位于青海省海北州祁连县峨堡镇(37°58′N,100°55′E),海拔3 512 m,属于高原大陆性气候.受高海拔的制约,该地区空气稀薄,大气透明度高,气温极低,年无霜期约130 d左右,年均气温约1.0℃,最冷月平均气温约 -13.2℃,最热月平均气温约12.8℃,年降水量为406 mm,年蒸发量为1 449.9 mm,太阳辐射强度为6 176 MJ/m2,风速约为5 m/s.植被主要为高寒草甸和灌丛草甸,优势高寒植物为高山嵩草(Kobresia pygmaea clarke)、麻花艽(Gentiana straminea Maxim)、矮嵩草(Kobresin humills)、垂穗披碱草(Etymus nutans)、异针茅(Stipa aliena)、美丽风毛菊(Saussurea supera)、线叶龙胆(Gentiana farreri balf)、垂穗鹅观草 (Roegneria nutans)、少叶早熟禾(Poa paucifoia keng)和雪白委陵菜 (Potentillanivealinn),灌 丛 为 金 露 梅(Hamamemelismois oliv)等.土壤类型为高寒草甸土,土层厚度一般为50~60 cm.
1.2 UV-B辐射增强处理与测定
2008年5月,在地势平坦地段选择了观测场并设立样地(面积666 m2).样地内均匀设立162块样方(正六边形,边长 0.80 m),9块样方为不架设UV-B灯(对照)、架设20 W和40 W的UV-B灯辐射强度处理,每个处理设3个平行.把UV-B灯悬挂在高寒草甸样地上方60~80 cm,每日07:00—19:00进行UV-B辐射照射,植物生长期间调整灯管高度以保持试验期辐射剂量基本一致,供试灯管为北京电光源厂生产的20 W和40 W的UV-B灯(UV-B 313),灯管包上一层纤维素双乙酸脂薄膜(又称AC膜,厚度为0.13 mm),用于过滤掉波长小于280 nm与生态不相关的少量致死性紫外线.经CALDWELL[18]公式转换,20 W 增强样地近地面UV-B生物有效辐射剂量约为10μW/cm2,40 W增强样地近地面UV-B辐射剂量约20μW/cm2,分别相当于测定区大气平流层中臭氧层衰减5%和10%后增加的UV-B辐射强度.在2008年6月1日开始进行UV-B辐射处理,每天以UV-B型紫外辐照计(北京师范大学光电仪器厂生产)定期测定297 nm处的 UV-B辐射强度,观测时间为 07:00—19:00,每h测1次,照射处理60 d后,测定植物光合作用及相关的生理参数.
1.3 麻花艽叶片光合作用及相关生理参数和环境要素测定
由于高寒草甸分布区生长季短,8月是高寒草甸植物生长旺盛期,为了分析UV-B辐射强度改变对高寒草甸典型植物生长旺盛期光合作用及相关生理参数日变化的影响,在天气晴朗的 8月 3日07:00—19:00进行观测,每h测定1次.需要说明的是,试验设计在7—9月都进行环境要素和植物光合作用相关参数日变化观测,但研究区生长季天气日变化较大,整日晴朗无云的天气少,多次观测因受到云遮挡和大风的影响,观测效果差,所以仅以8月3日比较有效的观测结果进行了分析.高寒草甸中植物组成多样,选择典型组成植物——麻花艽,每种UV-B辐射处理及对照下随机选择4株生长健壮的植株,测定上部充分展开叶片.以LI-6400光合作用测定系统(LI-COR,USA)测定选定植物叶片的净光合速率、蒸腾速率、气孔导度、胞间 CO2浓度、叶温及大气光合有效辐射强度、气温、相对湿度、大气 CO2浓度和大气饱和水气压,每次测定重复10~15次,取其平均值.
为分析UV-B辐射强度改变对高寒草甸典型植物光合作用及相关生理参数对光合有效辐射强度的响应差异,在天气晴朗的2008年8月17日和9月19日07:00—18:00,以LI-6400便携式光合测定系统测定麻花艽叶片光合作用相关生理生态参数,利用6400-02B LED红蓝光源,设定光合有效辐射强度为3 000,2 800,2 600,2 400,2 200,2 000,1 800,1 600,1 400,1 200,1 000,800,600,400,200, 100,50和 0μmol/(m2·s)梯度,叶室温度设为15℃,以6400-01C气源控制叶室内 CO2浓度为350μmol/mol.在测定样方内随机选取4株生长健壮麻花艽植株,测定上部充分展开叶片.当仪器显示各项指标变异系数小于1%,记录麻花艽叶片的净光合速率、蒸腾速率、气孔导度、叶温及光合有效辐射、气温、相对湿度和大气CO2浓度.测定完毕后采集麻花艽叶片计算叶面积,并进行光合系统叶面积换算.根据光合作用速率和蒸腾速率换算水分利用效率.
1.4 数据分析
利用SPSS 11.0软件,以方差分析法分析UVB辐射强度变化对麻花艽叶片光合作用的影响,以多重比较法分析差异的显著性,以相关分析法确定麻花艽叶片光合作用速率与大气温度、叶片温度、光合有效辐射强度、胞间 CO2浓度、大气饱和水气压和气孔导度的关系.
2 结果与分析
2.1 环境要素日变化
由图1可知,大气环境因子日变化趋势明显: 2008年8月3日,气温在07:00—09:00迅速上升,在09:00—11:00相对平稳,在11:00—13:00又上升,在13:00—14:00下降,在14:00—16:00又逐渐上升,在16:00—19:00逐渐下降,总体上在09:00—14:00较高,在13:00最高.叶温和大气饱和水气压变化趋势与气温变化趋势类似,也在09:00—14:00较高,在13:00最高.光合有效辐射强度在07:00—09:00逐渐上升,在 09:00—13:00相对平稳,在13:00—19:00逐渐下降.
差异性检验表明,8月3日不同时段这些环境要素在3种UV-B辐射处理下差异不显著,8月3日平均值的差异也不显著(P>0.05)(见表1).
2.2 UV-B辐射增加对麻花艽叶片净光合速率的影响
2008年8月3日,麻花艽叶片净光合作用速率日变化趋势明显.07:00—09:00呈上升趋势,09:00—13:00呈下降趋势,13:00—15:00又呈上升的趋势,15:00—19:00呈下降趋势;在 08:00—10:00达最高峰.不同UV-B辐射强度处理下麻花艽叶片净光合作用速率变化趋势基本一致,平均值差异显著(P<0.05),在对照下较高,在 20 W UV-B辐射下次之,在40 W UV-B辐射下较低(见表1).但07:00—09:00,11:00,14:00和16:00差异不显著(P>0.05)(见图2).

图1 2008年8月3日大气环境因子日变化Fig.1 The daily change of atmosphere factors on August 3rd,2008

表1 2008年8月3日麻花艽叶片净光合速率及相关生理参数与环境因子的平均值Table 1 The mean of net photosynthetic rate or related physiological parameters of straw-yellow gentian and environmental factors on August 3rd,2008
由图3可知,2008年8月和9月,麻花艽叶片净光合作用速率随着光合辐射强度发生改变.麻花艽叶片净光合速率都在光合有效辐射强度为 0~800μmol/(m2·s)时随光合有效辐射强度增加而增加,在2 200~3 000μmol/(m2·s)时变化不大.在相同光合有效辐射强度下,对照较高,在20 W和40 W UV-B辐射下较低,不同光合有效辐射强度下的平均值差异显著(P<0.05)(见表2),8月光合有效辐射强度0~400μmol/(m2·s)时差异不显著(P>0.05)(见图3).
2.3 UV-B辐射增加对麻花艽叶片气孔导度的影响
2008年8月3日,麻花艽叶片气孔导度呈现明显日变化趋势:在07:00—08:00较高,之后呈下降趋势,08:00—11:00呈上升趋势,11:00—13:00又呈下降趋势,13:00—15:00呈上升趋势,15:00—19:00呈下降趋势.麻花艽叶片气孔导度在对照下较高,在20 W UV-B辐射下次之,在40 W UV-B辐射下较低,8月3日平均值差异显著(P<0.05) (见表 1),在 08:00—10:00,13:00—14:00和16:00—19:00差异不显著(见图2).
由图4可知,2008年8月和9月,麻花艽叶片气孔导度都随光合有效辐射强度增加而呈增加趋势.相同光合有效辐射强度下,麻花艽叶片气孔导度在对照下较高,在20 W UV-B辐射下次之,在40 W UV-B辐射下较低(见图4),不同光合有效辐射强度下平均值差异显著(P<0.05)(见表2).
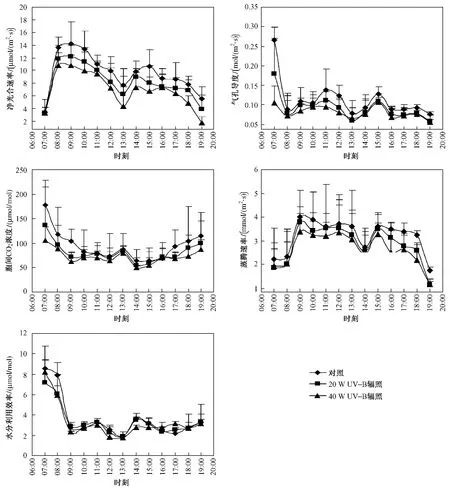
图2 2008年8月3日麻花艽叶片光合作用及相关参数日变化Fig.2 The daily change of photosynthesis and related physiological parameters of straw-yellow gentian on August 3rd,2008

表2 麻花艽叶片净光合速率及相关生理参数在不同光合有效辐射强度下的平均值Table 2 The mean of net photosynthetic rate or related physiological parameter of straw-yellow gentian under different UV-B radiation
2.4 UV-B辐射增加对麻花艽叶片胞间CO2浓度的影响
2008年8月3日,麻花艽叶片胞间CO2浓度呈日变化过程:在 07:00—10:00较高,之后下降,10:00—12:00相对平稳,12:00—15:00呈下降趋势,15:00—19:00又呈上升趋势.总体上在日出和日落时较高,中午较低.麻花艽叶片胞间CO2浓度在对照下最高,在20 W UV-B辐射下次之,在40 W UV-B辐射下最低,8月3日平均值差异显著(P<0.05)(见表1),在10:00—16:00差异不显著(P<0.05)(见图2).

图3 不同UV-B辐射强度下麻花艽叶片净光合速率对光的响应Fig.3 Response of leaf photosynthetis of straw-yellow gentian to photosynthetical active radiation under different UV-B radiation

图4 不同UV-B辐射强度下麻花艽气孔导度对光的响应Fig.4 The response of stomatal conductance of straw-yellow gentian to photosynthetical active radiation under different UV-B radiation
图5显示,2008年8月和9月麻花艽叶片胞间CO2浓度随着光合有效辐射强度的变化而改变.麻花艽叶片胞间CO2浓度在光合有效辐射强度为0~800μmol/(m2·s)时随光合有效辐射强度增加而呈迅速下降趋势,在800~3 000μmol/(m2·s)时变化不大.在相同光合有效辐射强度下,麻花艽叶片胞间CO2浓度在40 W UV-B辐射下较低,在20 W UV-B辐射和对照下较高(P<0.05)(见图5),不同光合有效辐射强度下平均值差异也显著(P<0.05)(见表2).
2.5 UV-B辐射增强对麻花艽叶片蒸腾速率和水分利用效率的影响
图2显示,2008年8月3日麻花艽叶片蒸腾速率也呈日变化趋势:在09:00—13:00和14:00—18:00较高,在 07:00—09:00呈上升趋势,在09:00—13:00相对平稳,在13:00—14:00呈下降趋势,在14:00—15:00又呈上升趋势,在15:00—18:00相对平稳,在18:00—19:00又呈下降趋势.并且在对照下较高,在20 W UV-B辐射下次之,在40 W UV-B辐射下较低,8月3日平均值差异显著(P<0.05)(见表1),在08:00—09:00和14:00差异不显著(P>0.05)(见图2).

图5 不同UV-B辐射强度下麻花艽胞间CO2对光的响应Fig.5 The response of intercellular CO2concentration of straw-yellow gentian to photosynthetical active radiation under different UV-B radiation
图6显示,2008年8月和9月,麻花艽叶片蒸腾速率随光合有效辐射强度增加而呈现增加趋势.相同光合有效辐射强度下,在40 W UV-B辐射下较低,在20 W UV-B辐射和对照下较高(见图6),不同光合有效辐射强度下平均值差异显著(P<0.05)(见表2).
图2显示,2008年8月3日麻花艽叶片水分利用效率也呈明显的日变化趋势:在07:00—08:00较高,之后下降,09:00—11:00呈上升趋势,11:00—13:00呈下降趋势,13:00—15:00又呈上升趋势,15:00—19:00平稳.麻花艽叶片水分利用效率在对照下最高,在20 W UV-B辐射下次之,在40 W UV-B辐射下最低,但 09:00—11:00,16:00和18:00—19:00差异不显著(见图2),8月3日平均值差异也不显著(P>0.05)(见表1).

图6 不同UV-B辐射强度下麻花艽蒸腾速率对光的响应Fig.6 The response of transpiration rate of straw-yellow gentian to photosynthetical active radiation under different UV-B radiation
图7显示,2008年8月和9月,麻花艽叶片水分利用效率在光合有效辐射强度也随光合有效辐射而发生改变.麻花艽叶片水分利用效率在光合有效辐射强度为0~800μmol/(m2·s)时随光合有效辐射强 度 增 加 而 呈 增 加 趋 势,在 800 ~3 000 μmol/(m2·s)时随光合有效辐射强度增加而呈下降趋势.相同光合有效辐射强度下,在40 W UV-B辐射下较低,在20 W UV-B辐射和对照下较高,不同光合有效辐射强度下平均值差异显著(P<0.05) (见表2),在0~400μmol/(m2·s)差异不显著(P>0.05)(见图7).
2.6 麻花艽叶片光合速率日变化与其他环境因子的关系
麻花艽叶片光合作用受多种环境因素影响.2008年8月3日,麻花艽叶片净光合作用速率受叶片气孔导度、胞间 CO2浓度、叶片温度、大气温度、光合有效辐射和大气饱和水气压的共同影响,净光合作用速率与叶片气孔导度、叶片温度、大气温度、光合有效辐射和大气饱和水气压呈正相关,与叶片胞间CO2浓度呈负相关,与光合有效辐射相关系数最高,与大气温度和叶片温度相关系数次之,与胞间CO2浓度、气孔导度和大气饱和水气压相关系数较低(见表3).

图7 不同UV-B辐射强度下麻花艽叶片水分利用效率对光的响应Fig.7 The response of water use efficiency of straw-yellow gentian to photosynthetical active radiation under different UV-B radiation
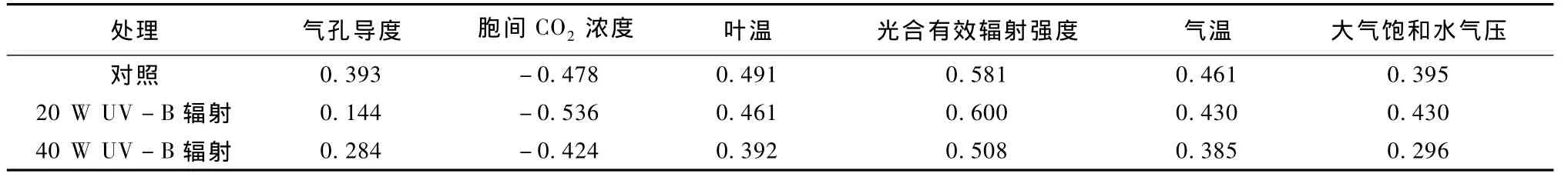
表3 麻花艽叶片净光合速率与环境因子及其他生理变量的相关系数Table 3 The correlation coefficient between net photosynthetic rate of straw-yellow gentian with environmental factors
3 讨论
UV-B辐射增强对植物光合作用有显著影响.如左园园等[10]研究UV-B辐射强度对2年生青榨槭幼苗的影响发现,UV-B辐射增强处理50 d使青榨槭幼苗每日净光合速率降低;陈章和等[19]研究不同UV-B辐射强度处理下南亚热带森林木本植物光合作用差异发现,UV-B辐射增加处理30 d或60 d使幼苗的净光合速率降低,较低UV-B辐射处理水平下净光合速率比对照低,较高UV-B辐射水平比较低水平处理下低;王春乙等[14]总结得出UV-B辐射增强将使植物光合速率降低.该研究表明,UV-B辐射增强使麻花艽叶片净光合速率降低.然而,不是所有植物都对 UV-B辐射变化敏感,一些高海拔植物不受 UV-B辐射增加影响[20].实验室研究发现,一些植物有适应 UV-B辐射机制.如人工种植的麻花艽,在不同强度UV-B辐射处理下,麻花艽叶片光合作用短期内有一定的抑制作用,随着处理时间的增加,该植物便很快适应强UV-B辐射处理[21],表明UV-B辐射变化对植物影响过程比较复杂.
UV-B辐射增强对植物水分代谢也有显著影响.如左园园等[10]研究 UV-B辐射强度对2年生青榨槭幼苗影响表明,UV-B辐射增强处理50 d使青榨槭幼苗每日水分利用效率降低;王春乙等[14]总结得出UV-B辐射增强将使植物蒸腾作用减弱.该研究表明,UV-B辐射增强使麻花艽叶片蒸腾作用及水分利用效率降低.
UV-B辐射增强对植物水分代谢和气孔导度也有显著影响.如陈章和等[19]研究不同 UV-B辐射强度处理下南亚热带森林木本植物光合作用差异发现,UV-B辐射增加也明显降低了幼苗叶片气孔导度,较低UV-B辐射处理幼苗叶片气孔导度比对照低下,较高UV-B辐射处理下幼苗叶片气孔导度比较低处理低,UV-B辐射对阳生性植物影响较耐荫植物影响大.该研究表明,UV-B辐射增强使麻花艽叶片气孔导度和胞间CO2浓度降低.
UV-B辐射增加对植物光合作用及相关生理过程影响受大气环境多种因素影响[12].PHOENIX等[11]研究发现,UV-B辐射增加和降水改变影响瑞典亚极地石楠被,但降水和UV-B辐射增加对石楠群落的影响存在相互抵消的作用;CALDWELL等[4]研究发现,UV-B辐射增强使植物对高温忍耐增加,并且高温影响植物对UV-B辐射变化的敏感性;DORMANN等[12]研究发现,极地温度升高和UV-B辐射增加使植物和养分循环改变.该研究表明,麻花艽叶片光合作用速率受叶片气孔导度、胞间CO2浓度、叶温、气温、光合有效辐射强度和大气饱和水气压影响,并且与叶片气孔导度、叶温、气温、光合有效辐射强度和大气饱和水气压呈正相关,与叶片胞间CO2浓度呈负相关,其中与光合有效辐射强度相关系数最高,且净光合速率在光合有效辐射强度为0~800μmol/(m2·s)时随光合有效辐射强度增加而增加的幅度大;气孔导度和蒸腾速率随光合有效辐射强度增加而增加;胞间CO2浓度在光合有效辐射强度为0~800μmol/(m2·s)时随光合有效辐射强度增加而呈迅速下降趋势;水分利用效率在光合有效辐射强度为0~800μmol/(m2·s)时随光合有效辐射强度增加而呈增加趋势,在800~3 000 μmol/(m2·s)时呈下降趋势.说明高寒草甸植物光合作用相关参数受光合有效辐射强度变化影响较大.UV-B辐射增加和其他环境因素改变对陆地生态系统结构与功能影响较为复杂[4,6],对这些影响还需要深入研究.
UV-B辐射增强对植物光合作用影响与植物生理生化过程有关,包括UV-B辐射增强对植物细胞酶系统,DNA,光合叶绿素,类囊体膜功能,叶片气孔行为和解剖结构的影响等[13-14,22].该研究发现,UV-B辐射增加对麻花艽叶片光合作用及相关生理参数的影响,可能与UV-B辐射增强后破坏了麻花艽叶片光合系统、酶系统和细胞膜有关.另外,UV-B辐射增强对麻花艽生理代谢过程也可能产生破坏性影响,对此需进一步研究.
4 结论
a.高寒草甸区大气环境因子日变化趋势明显. 2008年8月3日,气温在09:00—14:00较高,在13:00最高,叶温和大气饱和水气压与气温变化趋势基本类似.光合有效辐射强度在07:00—09:00逐渐上升,09:00—13:00相对平稳,13:00—19:00逐渐下降.
b.UV-B辐射增强使麻花艽叶片净光合作用、气孔导度和胞间 CO2浓度降低.2008年8月3日,麻花艽叶片净光合作用速率在对照下最高,在20 W UV-B辐射下次之,在40 W UV-B辐射下最低,且在08:00—10:00最高;气孔导度在07:00—08:00较高,胞间 CO2浓度在日出和日落时较高.净光合速率在光合有效辐射强度为 0~800 μmol/(m2·s)时随光合有效辐射强度增加而增加,在2 200~3 000μmol/(m2·s)时变化不大;胞间CO2浓度在光合有效辐射强度为 0~800 μmol/(m2·s)时随光合有效辐射强度增加而呈下降趋势,在800~3 000μmol/(m2·s)时变化不大;气孔导度随光合有效辐射强度增加而呈增加趋势.
c.UV-B辐射增强使麻花艽叶片蒸腾速率和水分利用效率降低.2008年8月3日,麻花艽叶片蒸腾速率和水分利用效率在对照下较高,在20 W UV-B辐射下次之,在40 W UV-B辐射下较低,且在09:00—13:00和14:00—18:00较高;水分利用效率在日出后较高.麻花艽叶片蒸腾速率随光合有效辐射强度增加而呈增加趋势;水分利用效率在光合有效辐射强度为0~800μmol/(m2·s)时随光合有效辐射强度增加而呈增加趋势,在800~3 000 μmol/(m2·s)时呈下降趋势.
d.多种环境因素共同影响麻花艽叶片光合作用速率.麻花艽叶片净光合作用速率与叶片气孔导度、叶温、气温、光合有效辐射和大气饱和水气压呈正相关,与叶片胞间 CO2浓度呈负相关,其中与光合有效辐射强度相关系数较高,与气温和叶温相关性次之,与胞间 CO2浓度、气孔导度和大气饱和水气压相关性较低.
[1]MADRON IS,MCKENZIE R L,BJ¨ORN L O,et al.Changes in biologically active ultraviolet radiation reaching the earth's surface[J].Journal Photochemistry and Photobiology B:Biology,1998,46(1/2/3):5-19.
[2]United Nations Environment Programme (UNEP).The environmental effects assessment panel report for 2006[M/ OL]// Environmental effects of ozone dep letion and its interactions with climate change:2006 assessment.Nairobi,Kenya:United Nations Environment Program(UNEP),2008[2008-03-04].http://www.unep.org/ozone,http://www. unep.ch/ozone.
[3]MCKENZIE R L,AUCAMP P J,BAIS A F,et al.Changes in biologically-active ultraviolet radiation reaching the Earth's surface[J].Photochemical& Photobiological Sciences,2007,6 (3):218-231.
[4]CALDWELL M M,BORNMAN J F,BALLARÉC L,et al. Terrestrial ecosystems,increased solar ultraviolet radiation,and interactions with other climate change factors[J].Photochemical&Photobiological Sciences,2007,6(3):252-266.
[5]FLINT S D,RYEL R J,CALDWELL M M.Ecosystem UV-B experiments in terrestrial communities:a review of recent findings and methodologies[J].Agricultural and Forest Meteorology,2003,120(1/2/3/4):177-189.
[6]KAKANIV G,REDDY K R,ZHAO D,et al.Field crop responses to ultraviolet-B radiation:a review[J].Agricultural and Forest Meteorology,2003,120(1/2/3/4):191-218.
[7]OUDEJANS A M C,NIJSSENA,HULS JS,etal.The reduction of aboveground Calamagrostis epigeios mass and tiller number by enhanced UV-B in a dune-grassland ecosystem[J].Plant Ecology,2001,154(1/2):37-48.
[8]PAOLETTI E.UV-B and mediterranean forest species:direct effects and ecological consequences[J].Environ Pollut,2005,137(3):372-379.
[9]SULLIVAN JH.Possible impacts of change in UV-B radiation on North American trees and forests[J].Environ Pollut,2005,137 (3):380-389.
[10]左园园,刘庆,林波,等.短期增强UV-B辐射对青榨械幼苗生理特性的影响[J].应用生态学报,2005,16(9):1682-1688.
[11]吴杏春,林文雄,郭玉春,等.植物对UV-B辐射增强响应研究进展[J].中国生态农业学报,2001,9(3):52-55.
[12]赵平,曾小平,孙谷畴.陆生植物对UV-B辐射增量响应研究进展[J].应用与环境生物学报,2004,12(1):122-127.
[13]师生波,贲贵英,赵新全,等.增强UV-B辐射对高山植物麻花艽净光合速率的影响[J].植物生态学报,2001,25(5): 520-524.
[14]王春乙,郭建平,郑有飞.二氧化碳、臭氧、紫外辐射与农作物生产[M].北京:气象出版社,1997:127-188.
[15]李英年,赵亮,徐世晓,等.祁连山海北高寒草旬紫外辐射与气象要素的关系[J].干旱区研究,2008,25(2):266-273.
[16]江灏,季国良,师生波,等.藏北高原紫外辐射的变化特征[J].太阳能学报,1998,19(1):7-12.
[17]周党卫,韩发,朦中华,等.UV-B辐射增强对植物光合作用的影响及植物的相关适应性研究[J].西北植物学报,2002,22(4):1004-1010.
[18]CALDWELL M M.Solar UV irradiation and the growth and development of higher plants[C]//GIESE A C.Photophysiology. New York:Academic Press,1971.
[19]陈章和,朱素琴,李韶山,等.UV-B辐射对南亚热带森林木本植物幼苗生长的影响[J].云南植物研究,2000,22(4): 467-474.
[20]TURUNEN M,LATOLA K.UV-B radiation and acclimation in timberline plants[J].Environ Pollut,2005,137(3):390-403.
[21]岳向国,韩发,师生波,等.不同强度的UV-B辐射对高山植物麻花艽光合作用及暗呼吸的影响[J].西北植物学报,2005,25(2):231-235.
[22]蒋高明.植物生理生态学[M].北京:高等教育出版社,2004.
Effects o f Enhanced UV-B Radiation on the Photosynthesis and Re lated Physiological Param eters of Straw-Yellow Gentian (Gentiana stram inea Maxim)
WU Jian-guo
Chinese Research Academy of Environmental Sciences,Beijing 100012,China
X06
A
1001-6929(2010)11-1395-10
2010-05-03
2010-07-07
中央级公益性科研院所基本科研业务专项(2007KYYW05)
吴建国(1971-),男,甘肃张掖人,研究员,博士,主要从事气候变化生态影响及其适应研究,wujg@craes.org.cn.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
湖北农业科学(2021年13期)2021-08-04
新疆农业科技(2021年2期)2021-07-03
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
空间科学学报(2020年6期)2020-07-21
中学生物学(2020年12期)2020-03-30
天津城建大学学报(2015年5期)2015-12-09
空间控制技术与应用(2014年2期)2014-04-17
