
孔雀新城疫病毒F基因的克隆及其变异分析
2010-08-07 01:39:12鞠厚斌曹火仁张维谊周锦萍
中国预防兽医学报 2010年7期
鞠厚斌,曹火仁,张维谊,刘 健,周锦萍
(1.上海市动物疫病预防控制中心,上海201103;2.上海市松江区动物疫病预防控制中心,上海201611)
新城疫(Newcastle disease,ND)是由 ND病毒(NDV)引起禽类的一种高度接触性、急性败血性传染病,多见于鸡和珍珠鸡,也能感染其他家禽和野禽[1]。该病广泛存在于世界各地,特别是强毒株可导致严重的内脏器官的损伤和神经系统疾病,死亡率高[2],对养禽业构成了极大威胁。目前,NDV疫苗的使用是预防该病的主要手段[3]。NDV F基因编码的融合(F)蛋白是病毒感染细胞的重要成分,通过它完成病毒囊膜与细胞膜的融合,导致病毒穿过细胞膜进入细胞内复制[4]。
本实验针对上海某珍禽场送检的2只病死孔雀进行剖检,结合其流行特点及发病情况疑以ND[5-7]。进行了NDV荧光RT-PCR检测和F基因的序列分析,现将结果报告如下。
1 材料和方法
1.1 发病情况 该养殖场从2008年11月不断出现发病孔雀,表现为精神沉郁,食欲减退,少数有神经症状,发病5 d~6 d后,有2只死亡。
1.2 剖检变化 送检的2例孔雀,剖检肝均有白色坏死灶或有萎缩,肠道萎缩、糜烂,其中一只胰腺和脾脏均有坏死灶。
1.3 细菌学诊断 分别无菌采集孔雀脑、心、肝、脾、肾、肺等组织样品,接种于Glunbia羊血琼脂上,置37℃培养24 h进行细菌分离检测。
1.4 NDV荧光RT-PCR检测 采集病鸽的脑、气管、气管粘液、肝脏、肾、肺等组织样品,研磨,加入适量PBS制成悬液,7 500 r/min离心5 min,取上清,使用TaqMan探针NDV荧光RT-PCR检测试剂盒(深圳匹基生物有限公司,批号:20081001)进行检测。
1.5 F基因的克隆
1.5.1 引物的设计与合成 参考GenBank登录的Guangxi10/2003株(DQ485261)F基因序列,设计了两条特异性引物P1、P2,扩增长度为1 884 bp,引物由上海英骏生物技术有限公司合成,分别为:
P1:5'-TGATCCATCTCGACTGCTTAT-3';
P2:5'-ACCCGTGTATTGCTCTTTG-3'。
1.5.2 病毒总RNA的提取 使用AxyPrepTMBody Fluid Viral DNA/RNA Miniprep Kit提取病毒 RNA,具体操作按照说明书进行。
1.5.3 F基因的扩增 使用TaKaRa公司的一步法试剂盒对提取的病毒RNA进行RT-PCR扩增。反应体系为50 μL,扩增程序为:50℃反转录30 min,94℃2 min;再按以下条件进行30个循环:94℃30 s,45℃30 s,72℃2 min;最后72℃延伸10 min。结束后取5 μL PCR产物进行琼脂糖凝胶电泳鉴定。
1.5.4 F基因的克隆及序列分析 鉴定阳性的样本PCR产物,由上海英骏生物技术有限公司测序。测序结果用DNAStar软件分析,同时与GenBank登录的其他NDV F基因的核苷酸序列及其编码的氨基酸序列进行同源性比较。
2 结果与讨论
2.1 细菌学诊断 Glunbia羊血琼脂培养皿中未见有细菌生长。
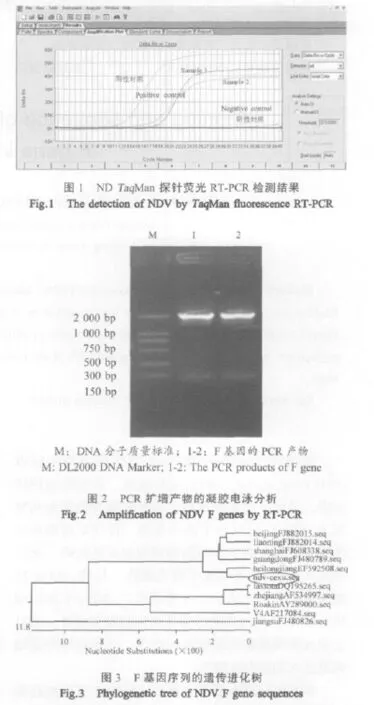
2.2 NDV荧光RT-PCR检测 2只孔雀病毒核酸检测结果均为NDV阳性(图1)。
2.3 目的基因扩增 应用所设计的特异引物,2份样品均扩增出预期的基因片段,大小为1884bp(图2)。
2.4 F基因的同源性分析与遗传进化树构建 为了进一步了解该毒株在遗传进化上的关系,应用DNAStar软件对该毒株的F基因与GenBank登录的10株国内外NDV F基因核苷酸进行比对,其同源性为81.1%~96.8%,推导的氨基酸序列同源性为78%~86.6%,其中与GenBank登录号为EF592508的黑龙江株同源性高达96.8%,与弱毒疫苗Lasota株的同源性只有81.1%。F基因的遗传进化树分析显示,该株与黑龙江毒株的遗传距离最近(图3)。但该株与Lasota株的同源性仅为81.1%,Lasota疫苗株免疫保护作用有待于进一步研究。
本研究中该毒株与黑龙江强毒株的核苷酸序列及其推导的氨基酸序列的同源性最高,遗传进化系谱树分析两者的亲缘关系最近,推测该毒株与黑龙江强毒株一样同属于强毒株,这为该毒株的毒力和致病性判断提供了分子生物学依据,这个推测与之前对NDV的生物学、免疫学特性以及NDV行业检测标准检测结果相符[9]。推导的氨基酸序列比对中,在117位~142位氨基酸区间内即融合多肽,NDV强毒株的融合多肽起始于苯丙氨酸(F),而NDV弱毒株融合多肽起始于亮氨酸(L),这也为NDV强弱毒的区分,从分子生物学角度提供了新的途径。然而Morrison等通过构建两株NDV强毒株的突变体证实融合多肽的氨基端苯丙氨酸与融合作用无关,与F0的裂解有关[10]。
在序列比对中还发现,虽然所得到的NDV F基因的核苷酸序列的全长为1 662 bp,与所比对的序列大小一致,但在F基因的核苷酸668处有一个T插入,并在867处缺失了一个T(图4),该毒株和黑龙江参考毒株核苷酸同源性为96.8%,但推导的氨基酸序列的同源性却为86.6%。虽然两者的遗传关系最近,对于是否由于T的插入和缺失增强了该病毒对孔雀的易感性,还有待深入研究。

随着畜禽业的发展,特种养殖在养殖业中的比例也不断增加,目前我国的免疫大都只局限于家禽,由于火鸡和孔雀的易感性较低[1],所以忽视了对孔雀等珍禽的免疫。本研究从孔雀内脏病料分离到NDV,通过病原学检测及对SPF鸡胚致病性指标的测定,确定感染的为强毒株。该毒株极易成为家禽和其它野生鸟类的传染源,因此提示特种畜禽的免疫工作也不可忽视,相关的病原感染来源及对其它鸟类的致病性等还有待进一步研究。
[1]殷震,刘景华.动物病毒学[M].2版.北京:科学出版社,1997:743-748.
[2]Alerander D J.Newcastle disease and other abian paramyxoviruse.Review[J].Rev Sci Tech,2000.19(2):443-462.
[3]Letellier C,Burny A,Meulemans G.Contruction of a pigeonpox virus recombiant:espression of the Newcastle disease vrius(NDV)fusion glycoprotein and protection of chickens against NDV challenge[J].Arch Virol,1991.118(1-2):43-56.
[4]Doms R W,Lamb R A,Rose J K,et al.Folding and assembly of viral membrane proteins[J].Virology,1993.193(2):545-562.
[5]武志强,于文涛,冯卫星,等.新城疫强毒(粤北株)的分离鉴定[J].中国兽医杂志,2006,42(11):52-53.
[6]俞伏松,胡奇林,陈少莺,等.1株鸡新城疫强毒株的分离与鉴定[J].中国兽医杂志,2001,37(8):24-25.
[7]王新卫,王岩,张建武.新城疫野毒株的分离鉴定[J].郑州牧业工程高等专科学校学报,2001,21(2):81-83.
[8]Toyoda T,Sakaguchi T,Hirota H,et al.Newcastle disease virus evolution.Ⅱ.Lack of gene recombination in generating virulent and abirulent strans[J].Virology,1989.169(2):273-282.
[9]Li Z,Sergel T,Razvi E,et al.Effect of clervage mutants on syncytium formation directed by the wild-type fusion protein of Newcastle disease virus[J].J Virol,1998.72(5):3789-3795.
[10]Morrison T,McQuan C,sergel T,et al.The tole of the amino terminus of F1 of the Newcastle disease virus fusion protein in cleavage and fusion[J].Virology,1993.193(2):997-1000.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
小哥白尼(野生动物)(2022年4期)2022-07-16 03:36:56
肝博士(2022年3期)2022-06-30 02:48:28
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
阅读与作文(小学高年级版)(2018年12期)2018-12-18 02:08:24
小天使·一年级语数英综合(2018年9期)2018-10-16 06:30:16
小青蛙报(2016年18期)2016-07-13 02:11:22
湖南畜牧兽医(2016年1期)2016-06-05 08:37:51
兽医导刊(2015年7期)2016-01-04 11:59:56
癌变·畸变·突变(2014年2期)2014-03-01 04:39:42
