
香菇渣对土壤微生物和酶活性的影响
2010-03-10 05:39:38魏孔丽陈京津张生香
湖南农业科学 2010年5期
谢 放,魏孔丽,陈京津,张生香
(兰州交通大学化学与生物工程学院,甘肃 兰州 730070)
目前,我国已成为世界上第一大食用菌生产国[1]。2006 年全国食用菌产量(鲜量)达 1 474.1 万 t,占全球总量70%以上;2007年出口创汇14.24亿美元,比2006年增加3.23亿美元,已经跃居我国继粮、棉、油、果、菜之后的第六大农副产品和重要的创汇农产品[2]。食用菌的大规模生产,致使大量的香菇渣废料(又名菌糠或菌渣)产生,菌渣被随地丢弃或燃烧,不仅造成了资源的极大浪费,对水体生态、食用菌生产环境和大气也造成严重的污染[1-3]。大量报道分析[1-4],菌渣中富含有机物、多种矿质元素以及食用菌菌体蛋白、次生代谢产物等多种水溶性养分,对改良土壤、提高土壤肥力有重要意义。因此,如何充分利用菌渣,使其变废为宝,已成为人们普遍关心的问题。
土壤微生物和土壤酶作为土壤的生物学标志在一定程度上能反映土壤肥力状况,衡量土壤肥力的好坏[5]。目前国内外在土壤微生物和酶的研究方面已有很多报道,许景伟等[6]研究了不同类型黑松混交林中土壤微生物和酶与土壤养分的关系。王晓凌等[7]研究了不同耕作方式对土壤微生物量及酶活性的影响。邱莉萍等[8]研究了重金属污染及EDTA对土壤酶活性的影响。肖嫩群等[9]也进行了稻草还田对土壤微生物和酶活性的研究。香菇渣富含有机质和生物活性物质,如果作为土壤改良剂直接还田,对土壤微生物和酶活性会产生怎样的影响,这方面的研究目前还未见报道。因此,本研究通过室内模拟试验,将不同比例的香菇渣(25%,50%,75%)加入土壤,通过测定主要土壤酶的活性和土壤微生物的动态变化规律,试图从这些土壤生物学指标的变化方面来研究菌渣作为一种生物肥料对土壤肥力的影响,为解决食用菌渣对环境的污染和其有效利用提供科学依据。
1 材料与方法
1.1 供试材料
供试土壤采自甘肃省皋兰县忠和镇盐池沟旱砂田西瓜地。采样时,先除去上面的沙层,后采集0~20 cm土样。供试菇渣为兰州市安宁区一蘑菇种植户废弃的香菇渣。
1.2 研究方法
1.2.1 试验方法 2008年11月19日采用五点法取土样,装入封口袋带回实验室,自然风干,粉碎,过2 mm筛备用。供试菇渣也在实验室自然风干,粉碎,过2mm筛,充分混匀备用。
模拟试验设5个处理(25%香菇渣+75%土壤、50%香菇渣+50%土壤、75%香菇渣+25%土壤),0%香菇渣(CK 1)和100%香菇渣(CK2)分别作为对照。将备用的香菇渣和土壤按以上设计的比例充分混匀,装进底部连有一塑料吸管(长8.5 cm,直径1.1 cm)的塑料杯(高15 cm、直径6 cm,底部中心小孔直径为1.1 cm)中,塑料吸管中塞有脱脂棉。然后将塑料杯放在钻有小孔(直径1.1 cm)的木板上(长45 cm、宽35 cm),最后一起放在盛有蒸馏水的铁盘(长40 cm、宽30 cm、深3.8 cm)中,让塑料杯底部的吸管充分接触水面,从而使土壤保持田间持水量,恒温(27℃±0.5℃)培养室黑暗培养。试验始于2008年5月19日,分别于培养20、40、60、80和100 d 5次取样,每次每处理3个重复,每重复9点法取样,混合为1个混合土样。样品混匀后,自然风干,分别过1mm筛和过2 mm筛。过1 mm筛用于测定微生物数量;过2mm筛的土样,4℃保存,用来依次分析土壤磷酸酶、几丁质酶和蔗糖酶活性。
1.2.2 测定方法 土壤微生物数量用稀释平板法测定[10],细菌计数用牛肉膏蛋白胨培养基;放线菌用高泽氏一号培养基;真菌用马丁-孟加拉红链霉素琼脂培养基。
土壤酶活性的测定方法:磷酸酶活性用磷酸苯二钠比色法,用mg酚/(g·h)表示;蔗糖酶活性用3,5-二硝基水杨酸比色法,用 μg葡萄糖/(g·h)表示[11];几丁质酶活性用N-乙酰葡萄糖胺比色法,用μg N-乙酰葡萄糖胺/(g·h)表示[12]。
1.2.3 数据分析 数据的整理与统计采用Excel软件完成。
2 结果与分析
2.1 香菇渣对土壤微生物的影响
2.1.1 香菇渣对土壤细菌的影响 在不同的培养时间,添加不同比例的香菇渣对土壤细菌数量的影响不同(图1)。从图中可以看出,未添加菇渣的土壤中(CK1)细菌数量相对较少,添加菇渣后各个处理土壤中的细菌数量明显增加。从培养20 d开始,各处理细菌的数量就迅速增加,排列顺序为75%香菇渣>CK2>50%香菇渣>25%香菇渣>CK1,至40 d时添加75%香菇渣土壤中的细菌数量最多,为114×104CFU/g DW,比CK1增加了20.5倍,且大于CK2中细菌的数量。其原因是75%香菇渣中含有丰富的有机物质,为土壤细菌的生长提供了充分的营养物质,因此细菌数量大幅增加。而CK2未加入土壤,测定的细菌数目只是菇渣中原有的细菌,因此细菌种类和数目相对较少。当培养60 d时,各处理土壤中的细菌数量迅速降低,这可能是微生物活动达到高峰后,可生物利用的基质逐渐减少,氧化还原电位降低及还原性有毒物质增加,抑制了微生物生长的原因。培养80 d后,添加25%和50%香菇渣土壤中的细菌数和CK1又出现了一个小高峰,这可能与微生物的二次生长有关。但不同的是,75%香菇渣土壤中细菌数量一直呈缓慢下降趋势,而CK2(100%香菇渣)的细菌数则出现持续上升趋势,这可能与香菇渣渐渐腐解为微生物活动提供持续能源和碳源有关。总体来看,添加香菇渣都能显著影响土壤中细菌的数量,而且在一定范围内与添加量成正比。
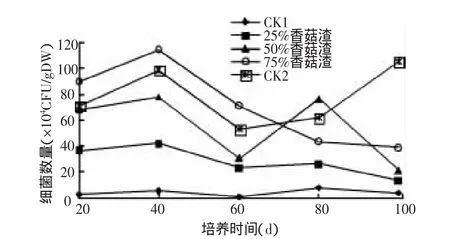
图1 香菇渣对土壤细菌数量的影响
2.1.2 香菇渣对土壤放线菌的影响 香菇渣对土壤放线菌数量的影响见图2。与CK1类似,25%和50%香菇渣处理的曲线变化较平缓,且增幅不大,说明香菇渣对土壤放线菌数量的影响不太明显。而添加75%香菇渣后,土壤中放线菌的数量显著增加,在培养20 d时就远远大于其他处理,为CK1(0%香菇渣)的25.2倍,为 CK2(100%香菇渣)的5.2倍;培养40 d时达到最大值,为72.3×104CFU/g DW。这与75%香菇渣为土壤放线菌提供丰富的纤维素、木质素、几丁质等营养物质,极大的诱导了放线菌数量的增加有关。之后随着时间的推移,75%香菇渣处理的放线菌数又开始缓慢下降,100 d时降到与其他处理相当的水平。而CK2的放线菌数量在培养80 d以前一直呈缓慢上升趋势,在80 d时达到32.5×104CFU/g DW后又急剧下降。与其他处理不同的是,CK2在80 d时出现高峰,这与放线菌常在有机质腐解的后期出现把难分解的组分转化为土壤的有机组分有关。由上可看出添加75%香菇渣对土壤放线菌数量的影响最大。
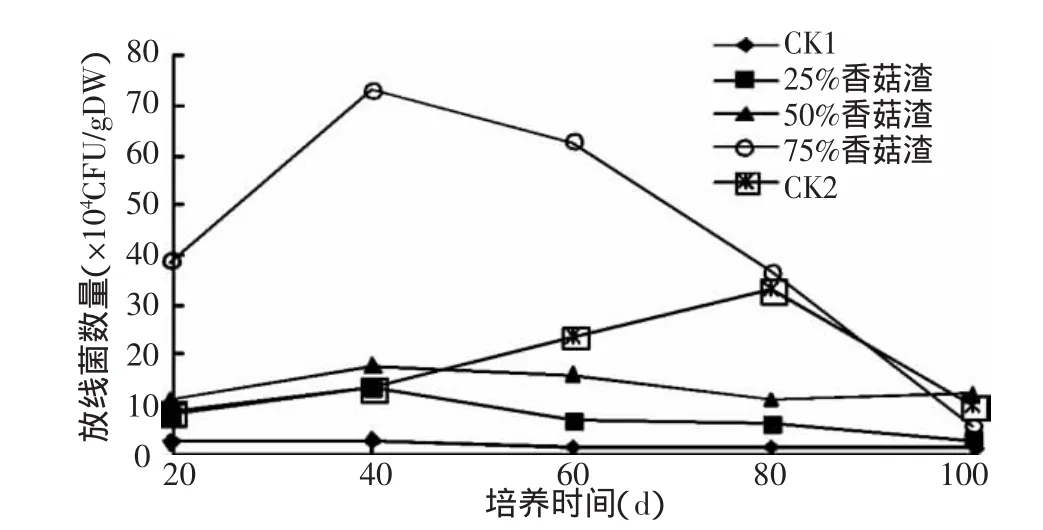
图2 香菇渣对土壤放线菌数量的影响
2.1.3 香菇渣对土壤真菌的影响 从图3看出,不添加香菇渣的土壤(CK1)中真菌数量非常少,在0.2×104~0.7×104CFU/g DW 之间。随着香菇渣添加量的增大,土壤中真菌数量也随之增加,排列顺序为CK2>75%香菇渣>50%香菇渣>25%香菇渣>CK1。其中,添加25%和50%香菇渣后真菌数量较CK1变化不明显,在60 d和100 d时数量最少;添加75%香菇渣后真菌数量出现显著变化,分别在培养40 d和80 d时出现高峰,达到30.6×104CFU/g DW和48.6×104CFU/g DW。与细菌和放线菌变化不同的是,CK2(100%香菇渣)在整个培养期,真菌数量始终大于其他处理,且在40~80 d时基本呈水平趋势,80 d后又迅速下降。这是因为纯香菇渣中含有丰富的有机质,而土壤真菌参与土壤中有机质的分解、腐殖质和团聚体的形成及土壤中的氨化作用[9],因此真菌数量大量增加,从而抑制了细菌和放线菌的生长,所以CK2的细菌和放线菌数量不是最多。之后由于大量微生物活动,使其可利用的营养物质迅速减少,因此培养80 d后微生物数量也急剧下降。
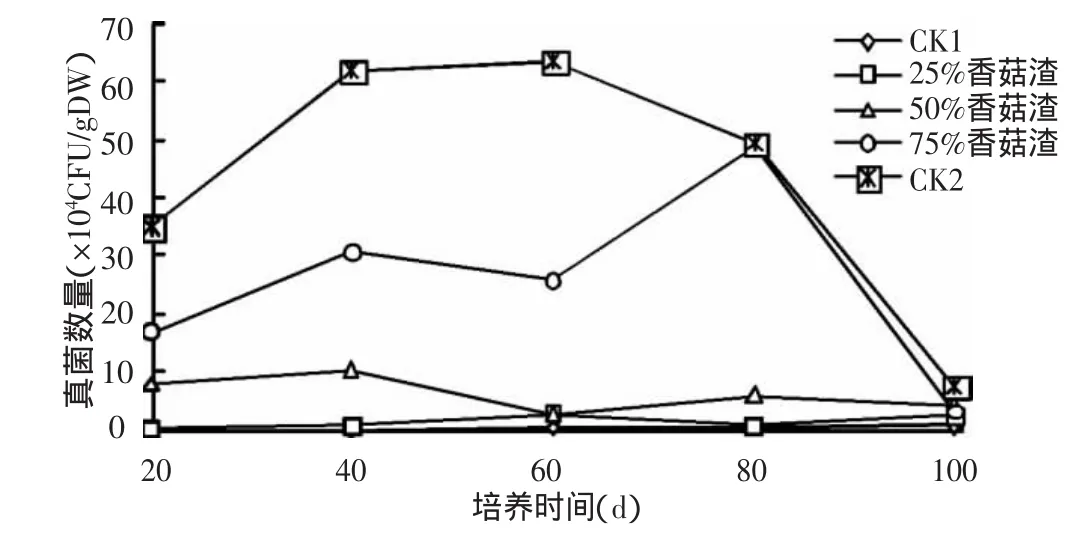
图3 香菇渣对土壤真菌数量的影响
2.2 香菇渣对土壤酶活性的影响
2.2.1 土壤磷酸酶活性 土壤磷酸酶是植物根系与微生物的分泌产物,与土壤磷素转化密切相关,其高低可以反映土壤速效磷的供应状况。图4反应了添加香菇渣对土壤磷酸酶活性的影响。从图中可以看出,25%和50%香菇渣处理的磷酸酶活性变化不大;而75%香菇渣的磷酸酶活性显著变化,在培养20 d时,较CK1大幅度提高了16.4倍,这说明香菇渣还田对土壤有机磷的矿化作用有影响。在整个培育期内,各处理的磷酸酶活性均呈现逐步下降的趋势,到培养100 d时下降到最低。这可能是因为土壤磷酸酶能促进菇渣中的有机磷化合物分解。但随着时间的推移,由于可供酶分解基质的减少和速效磷在土壤中的积累,磷酸酶活性受到一定的抑制作用,因此在整个培育期呈下降趋势。
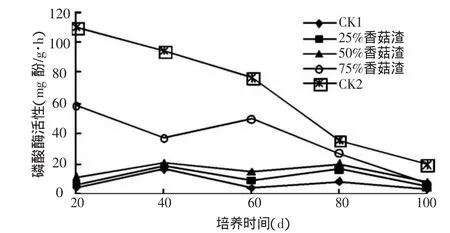
图4 香菇渣对土壤磷酸酶活性的影响
2.2.2 土壤几丁质酶活性 土壤几丁质酶能酶促几丁质生成几丁二糖,再经过几丁二糖酶作用,最后生成葡萄糖胺和乙酸。其酶促作用可使土壤增加矿化氮量,控制N循环速率的关键步骤[16]。从图5可看出,纯土壤(CK1)中几丁质酶活性非常低,随时间变化几乎呈直线趋势。添加香菇渣后几丁质酶活性开始发生变化,且与添加量成正比关系。添加25%和50%香菇渣后几丁质酶活性变化不太明显,而添加75%香菇渣的土壤中酶活性变化则很显著,在培养20 d时较CK1增加了3.3倍,培养60 d时达到最大,为 58.62 ug N-乙酰葡萄糖胺/(g·h),随后又开始下降。CK2(100%香菇渣)的几丁质酶活性始终大于其他各个处理,说明添加香菇渣对土壤中几丁质酶的活性有显著影响。

图5 香菇渣对土壤几丁质酶活性的影响
2.2.3 土壤蔗糖酶活性 蔗糖酶又名转化酶,是广泛存在于土壤中的一个重要的酶,它对增加土壤中易溶性营养物质起着重要作用。不同添加比例的香菇渣在不同培养时期蔗糖酶活性的变化如图6所示。从图中可以看出,各处理样的蔗糖酶在培养20 d时就显示了极大的活性,酶活性大小为25%香菇渣>CK2>CK1>50%香菇渣>75%香菇渣,其中25%香菇渣的蔗糖酶活性最高可能与土壤的C/N有关。这是因为菌渣作为农肥,其中的纤维素和木质素已被大量降解,又含有大量的菌体蛋白,具有微生物生命活动中所需的C/N,因此用作肥料时可提高土壤中的微生物数量,从而导致了酶活性的增加[3]。随着时间的推移,酶活性迅速下降,到培养40 d时各处理样的酶活性几乎降到最低,这与大量产物的抑制作用有关。之后酶活性又出现“先升后降”的趋势,在培养100 d时酶活性大小为75%香菇渣>CK1>25%香菇渣>50%香菇渣>CK2。说明香菇渣对土壤蔗糖酶活性的影响时间较短,且添加75%的香菇渣有利于对土壤肥力的长期作用。
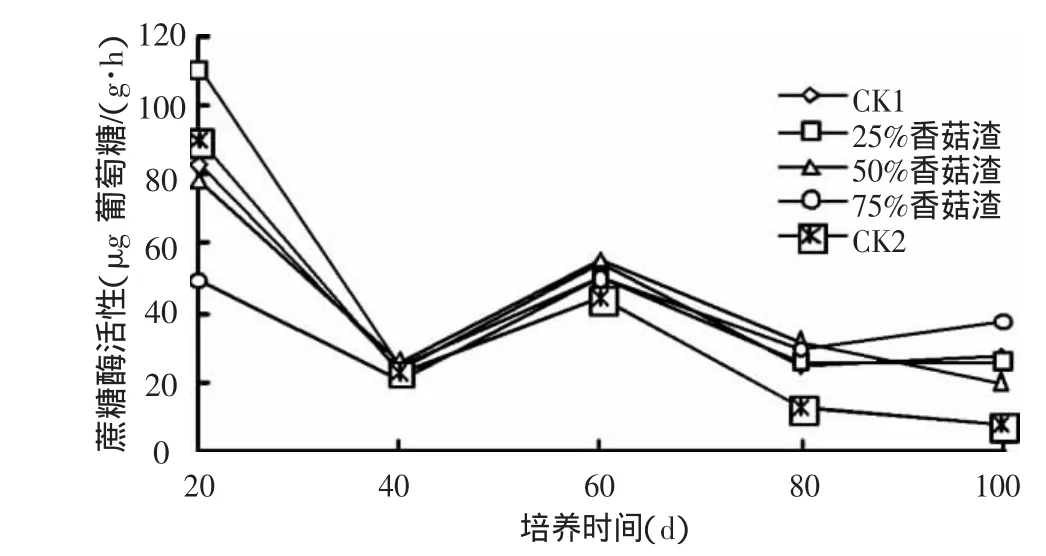
图6 香菇渣对土壤蔗糖酶活性的影响
3 讨论
3.1 添加香菇渣对土壤微生物数量的影响
土壤微生物是土壤中活的有机体,是最活跃的土壤肥力因子之一。细菌、放线菌和真菌是土壤微生物的三大类群,构成了土壤微生物的主要生物量,它们的区系组成和数量变化常能反映出土壤生物活性水平[17]。本研究中,添加不同比例的香菇渣均能促进土壤微生物活动,但对于不同种类的微生物其效应不同。添加香菇渣后细菌类群数量明显增加,反映出添加香菇渣后的土壤具有良好的微生物生态学特性。据有关报道[19],土壤细菌数量与土壤养分含量、速效氮和速效磷含量呈正相关,且基本达到显著或极显著水平,体现出细菌在土壤生物过程和土壤肥力形成中占据着重要或突出的地位,因此细菌数量的明显增加表明添加香菇渣对土壤肥力的有利作用。添加香菇渣也能显著增加土壤放线菌的数量。但是孙瑞莲等[13]研究发现,土壤放线菌与土壤各养分因子之间的相关性不明显,因此无法通过放线菌的数量来评价其土壤肥力的高低。熊英等[20]指出真菌数量的增加可能标志着土壤肥力的降低,但刘久俊等[21]证明生物覆盖可以显著提高杨树人工林根际土壤真菌数量,与有机物在土壤中被分解后形成具有良好通气、蓄水能力的腐殖质[3],为真菌活动创造良好的环境条件有关。因此,添加香菇渣后真菌数量的增加可能也与土壤肥力的提高有关,需要进一步研究微生物种类之间的消长关系以及种群变化规律来更详细的解释。
3.2 添加香菇渣对土壤酶活性的影响
土壤酶活性是反映土壤生物化学过程的重要指标。土壤中物质的转化、累积和分解是一个复杂的生物化学过程,这些过程几乎都是在各种酶的作用下完成的。因此土壤酶活性常作为评价土壤生态环境质量的重要指标[22]。本研究中,随香菇渣添加量的增大,磷酸酶和几丁质酶活性都显著增加,这与黄玉霞[18]研究施肥可以增加菜园土壤磷酸酶活性的结论一致。磷酸酶与土壤全氮、全磷、碱解氮、速效磷等含量有非常好的相关关系,因而可以作为土壤肥力的重要指标指示土壤肥力的变化。而对于土壤中几丁质酶活性对土壤肥力指示作用的研究目前尚未见报道。本研究中几丁质酶活性变化主要是因为香菇渣中含有的食用菌菌体残渣可诱导土壤几丁质酶活性的增加,从而对防治植物真菌病害起重要作用[23]。试验结果表明添加香菇渣确实能极大提高土壤几丁质酶活性,为以后利用香菇渣来防治土传病害提供了理论基础。另外,试验发现,添加不同比例的香菇渣对蔗糖酶活性影响不大,且随培养时间的延长,出现下降趋势。这与王俊华等[24]施有机肥后可增加大量的有机碳,为蔗糖酶提供更多的酶促基质,最大程度的提高蔗糖酶活性的结果不一致。究其原因可能是因为蔗糖酶活性与植物的组成和年龄有密切的关系[15],而本试验所用的是瓜地土壤,在试验中并未种植植物,因此植物的根系较少,土壤蔗糖酶的活性较低,这与周礼凯[15]的报道一致。另外随培养时间的延长,酶活性下降,可能是因为土壤微生物消耗导致土壤C/N失衡,不利于土壤酶活性的提高[25]。
由于本试验是在室内模拟条件下研究添加香菇渣对土壤生物活性的影响,为了缩短试验时间,通过提高试验温度来加快土壤的变化进程,但这并不能反应大田中生物变化规律的真实情况,因为实际田间情况可能比试验中发生缓慢。因此,本试验结果只能为菇渣还田提供一个理论参考,对自然条件下土壤微生物数量和酶活性的影响还应考虑到温度变化、日变化和季节变化等因素,需要进一步的田间试验才能得出。
4 结论
(1)添加香菇渣能够显著提高土壤中细菌、放线菌和真菌数量。其中,细菌的数量增加最多,放线菌次之,真菌最少。添加25%香菇渣和50%香菇渣对细菌、放线菌和真菌的影响不大。添加75%香菇渣则能显著增加土壤中微生物的数量。
(2)添加香菇渣虽然不能明显增加土壤蔗糖酶活性,但能显著提高土壤磷酸酶和几丁质酶活性,且75%香菇渣的变化最明显,说明菇渣还田能增加土壤中氮素营养和磷的含量。
(3)75%香菇渣还田量更有利于提高土壤生物活性,极大地增加土壤微生物数量和酶活性,从而改变土壤结构,增加土壤肥力。
[1] 王德汉,项钱彬,陈广银.蘑菇渣资源的生态高值化利用研究进展[J].有色冶金设计与研究,2007,28(3):262-266.
[2] 王传福,李淑珍.食用菌产业在现代农业中的战略地位及发展前景展望[J].河南农业,2008,(17):11-12.
[3] 李用芳,李学梅,李鹤宾.香菇木屑菌渣营养成分分析及在平菇菌种生产中的应用[J].微生物学杂志,2001,21(3):59-60.
[4] 侯立娟,代祖艳,韩丹丹,等.菌糠的营养价值及在栽培上的应用[J].北方园艺,2008,(7):91-93.
[5] 唐玉姝,魏朝富,颜廷梅,等.土壤质量生物学指标研究进展[J].土壤,2007,39(2):157-163.
[6] 许景伟,王卫东,李 成.不同类型黑松混交林土壤微生物、酶及其与土壤养分关系的研究 [J].北京林业大学学报,2000,22(1):51-55.
[7] 王晓凌,陈明灿,张 雷.不同耕作方式对土壤微生物量和土壤酶活性的影响[J].安徽农学通报,2007,13(12):28–30.
[8] 邱莉萍,张兴昌.Cu Zn Cd和EDTA对土壤酶活性影响的研究[J].农业环境科学学报,2006,25(1):30-33.
[9] 肖嫩群,张杨珠,谭周进,等.稻草还田翻耕对水稻土微生物及酶的影响研究[J].世界科技研究与发展,2008,30(2):192-194.
[10]李阜棣,喻子牛,何绍红.农业微生物学实验技术[M].北京:中国农业出版社,1996.
[11]关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.
[12]顾向阳,胡正嘉.一种测定土壤几丁质酶活性的方法[J].土壤通报,1994,25(6):284-285.
[13]孙瑞莲,朱鲁生,赵秉强,等.长期施肥对土壤微生物的影响及其在养分调控中的作用 [J].应用生态学报,2004,15(10):1907-1910.
[14]USEPA.2000.Bacill us thuri ngiensis Plant pesticides Registration Action Document:Preliminary Risks and Benefits Sections.U.S.Environmental Protection Agency,Office of Pesticide Programs.Washington,DC.
[15]周礼凯.土壤酶学[M].北京:科学出版社,1987.
[16]Ekenler M,TabatabaiMA.β-Glucosaminidase activity of soils:effect of cropping systems and its relationship to nitrogen mineralization[J].Biology and Fertility of Soils,2002,36(5):367-376.
[17]唐玉姝,魏朝富,颜廷梅,等.土壤质量生物学指标研究进展[J].土壤,2007,39(2):157-163.
[18]黄玉霞,李俊华,褚贵新,等.施肥对菜地土壤微生物和土壤酶活性的影响 [J].石河子大学学报(自然科学版),2007,25(5):552-557.
[19]蔡晓布,彭岳林,薛会英,等.不同培肥方式对西藏中部退化土壤微生物的影响研究 [J].中国生态农业学报,2004,12(1):108-110.
[20]熊 英,邓福英,唐庆兰,等.万鹤山鹭鸟保护区土壤微生物优势类群生态分布的研究 [J].广西农业生物科学,2005,24(1):58-62.
[21]刘久俊,方升佐,谢宝东,等.生物覆盖对杨树人工林根际土壤微生物、酶活性及林木生长的影响[J].应用生态学报,2008,19(6):1204-1210.
[22]刘建国,卞新民,李彦斌,等.长期连作和秸秆还田对棉田土壤生物活性的影响[J].应用生态学报,2008,19(5):1027-1032.
[23]孟 亮,崔德才.植物几丁质酶及其在抗真菌病害中的应用[J].生物技术通讯,2004,15(4):420-422.
[24]王俊华,尹 睿,张华勇,等.长期定位施肥对农田土壤酶活性及其相关因素的影响[J].生态环境,2007,16(1):191-196.
[25]刘恩科,赵秉强,李秀英,等.长期施肥对土壤微生物量及土壤酶活性的影响[J].植物生态学报,2008,32(1):176-182.
猜你喜欢
生物技术通报(2021年4期)2021-05-14 06:01:28
国际呼吸杂志(2019年22期)2019-12-09 09:20:36
中国酿造(2017年8期)2017-09-03 06:20:01
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:45
广西林业科学(2016年1期)2016-03-20 05:33:00
医学研究杂志(2015年8期)2015-06-22 14:00:57
应用海洋学学报(2014年4期)2014-11-22 07:43:54
天然产物研究与开发(2014年6期)2014-04-27 14:15:56
中国医学科学院学报(2013年6期)2013-03-11 20:26:04
中国医学科学院学报(2013年2期)2013-03-11 20:25:49
