
对蜗牛Cepaea Nemo ralis多态性变异与杂合性相互关系的研究
2010-01-09 03:05高贵喜
太原师范学院学报(自然科学版) 2010年2期
高贵喜
(太原师范学院生物系,山西太原 030001)
对蜗牛Cepaea Nemo ralis多态性变异与杂合性相互关系的研究
高贵喜
(太原师范学院生物系,山西太原 030001)
Cepaea壳颜色与壳样式(pattern)上的多态性一直被认为与壳可见外型无关,而是由杂合优势所决定.检验这种说法的一种途径就是察看是否显性表型的变异性要比隐性表型的变异性低.研究对两组蜗牛壳进行观察,两组样本内个体数分别是1 972和5 423个,经分析,隐性表型的变异性比显性表型显著(P<0.05).在同一壳中,对壳测量所得数据进行相关分析,表性性状差异的证据未找到.如果变异的差异是真实的,那么将表明或者是可见的壳外型是不重要的,多态性由于杂合平衡决定的,或者是这种多态性平衡的进化是由壳外形的自然选择所维持的.
杂合性;表型性状变异;多态性;相关平衡;Cepaea
0 引言
陆地蜗牛Cepaea nemoralis和C.hortensis广泛分布于欧洲.它们生活在丛山边裸岩、沙地与落叶树相混合的林地到灌丛和草地的广泛生境中,大多数蜗牛在壳的颜色和斑纹上表现为多态性,这种多态性由完全显性基因控制.决定壳的颜色及斑纹有无是两个主要的基因位点,它们是相互紧密连锁的,同时基因位点的数目可修饰斑纹的表达[1].尽管形态频度(morph frequencies)在短距离内有变异,但这里很难有这样一群体,其任一形态频度没有在10%~90%之间.这通常视为连锁的不平衡性.自然选择与形态频度的决定相关联[2~4],但是多形态是以怎样方式维持下来的还是不清楚.
自然选择与可见的壳颜色和壳式样相联系,其证据是由于热量不同,在颜色上导致形态方面的相异,但也有可能是种群保持这种多形态性是由于杂合优势或杂种优势在不可见方面起作用.这方面的探索者是Goodhart[5].他解释道,大多数的陆地蜗牛作为一种平衡选择的结果在遗传上是异源性的.如果相关的基因对可见的影响是有害的,这种可见的表达就会被抑制,因为在其他蜗牛中,壳的形态是整齐划一的.如果壳的颜色和斑纹不能适应环境,那么它们就会在平衡多态性下被淘汰.Goodhart认为C.nemoralis和C.hortensis就属于后者.
为了更好地了解多态性,我们应当找到一种方法来验证杂交优势.现在经常应用的方法是通过对样本在不同水平的表型变化上酶基因位点来测定一定量的多态性[6~8].这种方法的预测是较低的形态变异和降低不对称性将会与较多的基因组杂合型相关联.在双壳软件动物中大量研究用来调查这种关系.杂合个体与纯合个体相比较,其表现出发育较快、生活的更长以及更能有效地合成蛋白质[9].当杂合性与生长速度正相关时,则与此类动物体积大小成负相关[10~11].有些研究材料支持杂合优势,也有些解释为超显性.杂合性和变异之间的关联是基因组相互作用的.
用随机方法从多态型种群中收集蜗牛壳,通过测定个体壳的变异来比较显性个体与隐性个体.显性性状组成的个体对应的等位基因可以是杂合的也可以是纯合的;而隐性组的个体相对应的等位基因则是纯合的.如果基因组杂合性与较低表型变异相关联,那么同一生活地的显性形态壳样本要比隐性形态壳样本在表型变异方面要小些.测定成年蜗牛壳三维数据,因为壳的变化记录了蜗牛的发育过程中所受到任何因子的影响.Foote和Cow uie[12]对蜗牛Theba pisana进行了三维度量,结果表明发育自动调节稳态(developmental homeostasis)在起作用.壳三维度量的遗传率(heritability)被看做为亲子回归已有记录,比如对于C.nemoralis其值为0.6[1].
1 材料和方法
收集的C.nemoralis样本来自于两方面.第一是蜗牛个体在一封闭状态下繁殖,其目的是用来检验环境对壳形态频度和壳大小的影响[13].对此群体每年进行监测,将成年个体加以收集,记录表型性状并测定壳的最大直径(精确到0.1mm).由于种群密度的增加,成年蜗牛的平均直径在下降,这可能是由于蜗牛遭受到一些竞争压力的缘故.有关这方面的生态学细节Cook和Cain[12]有过讨论.把粉红色个体(粉红色是显性表型)的宽度和黄色个体壳的宽度进行测量,测量的个体数目总共是1 792(其中黄色个体是18个).
蜗牛第二来源于A.J.Cain大量收集的标本,现存于曼彻斯特博物馆.这些随机收集的标本是A.J.Cain和他的合作者在过去45年中为研究Cepaes的多态性而收集的.此次使用材料只选择了由Burke[14]所收集的标本来研究.Burke在很短时间内从爱尔兰特定的地区收集了大量的标本.每个标本确定它的颜色(黄色或非黄色)、斑纹(有或无斑纹,无斑纹为显性)测量每一个壳的最大宽、高度和壳口的宽度(精确到0.1 mm),共测量了5 423个体.
对爱尔兰蜗牛来说,壳斑纹的平均频率是0.740±0.241.黄色壳平均频率是0.403±0.285.根据 Hardy-Weinberg公式,利用这些形态频率数据可以计算出基因型频率,我们可以计算在两基因位点上显性性状的杂和体平均频率.对斑纹基因位点,其值是0.905±0.110;对颜色基因位点,其值是0.774±0.150.每种性状,显性具有较高的杂合频率.
表型形状之间进行了两类型比较,一类是显、隐性性状的总变异(overall variability)如果遗传性杂合体确实在控制成年蜗牛壳的大小、式样起主要作用,那么显性性状的变异性要比隐性性状变异性低;第二类是相关性,如果差异没有不同,某一地点的蜗牛在度量上就会有较大相关性,即某地点蜗牛较其他地点的蜗牛在形态上更一致.
在比较表性性状的变异时,我们使用了封闭环境的蜗牛壳宽度方差和爱尔兰蜗牛壳三维测量总方差.这样做的目的是比较壳的形态由显性性状控制与隐性性状控制的差异.比较是在封闭蜗牛中淡红色与黄色之间进行,爱尔兰蜗牛的比较是在黄色与非黄色、具斑纹与不具斑纹之间进行.对单个样本,检验方法是F检验,因为样本是随机的,故样本中的个体数目也是不确定的,有时会出现一样本中的个体数目比另一样本的个体数目少得多.为了尽可能多地比较样本,我们对样本个体数目限定在大于和等于5,显然,小样本的偶然性影响是极有可能发生的.
F检验通常是有尾检验,即组间方差与组内方差所除.对于我们的研究,一尾检验是有意义的,因为离差方向被确定了.但是当单个检验结果组合起来,我们就应该确定离差方向了.为了做好上述及考虑到样本大小之差异,我们按下列步骤做一些处理:
1)对每一样本,较大方差被较小方差所除,同时差异方向做了记录.
2)F值或然率被计算,其方法是在观察自由度下,用数值积分法对F分布的对应段加以积分[15].
3)等值正态偏差由或然率代替,计算方法同第二项.
4)如果隐性性状方差比显性的小,则方差为负;反之为正.
5)把偏差值相加,再用偏差数目的平方根来除,这样得到单一正态偏差,次数值可以度量所有数据在特定方向上从整合到分散的方差比率.
为了验证相关性,我们计算了爱尔兰蜗牛3个测量值的相关系数r的平均值.显性性状应该具有较小的变异性.为了检验这点,将r值转化为 Hotelling的Z*值,次值实际上是方差的1/(n-1),这里代表组内的个体数目[16,17].在同一样本内,两性状(显性、隐性)平均值用t检验比较.将t值转化为正态离差来检验趋向性,这样可度量一种性状比另一种性状具有更大变异能力的趋向(表1).
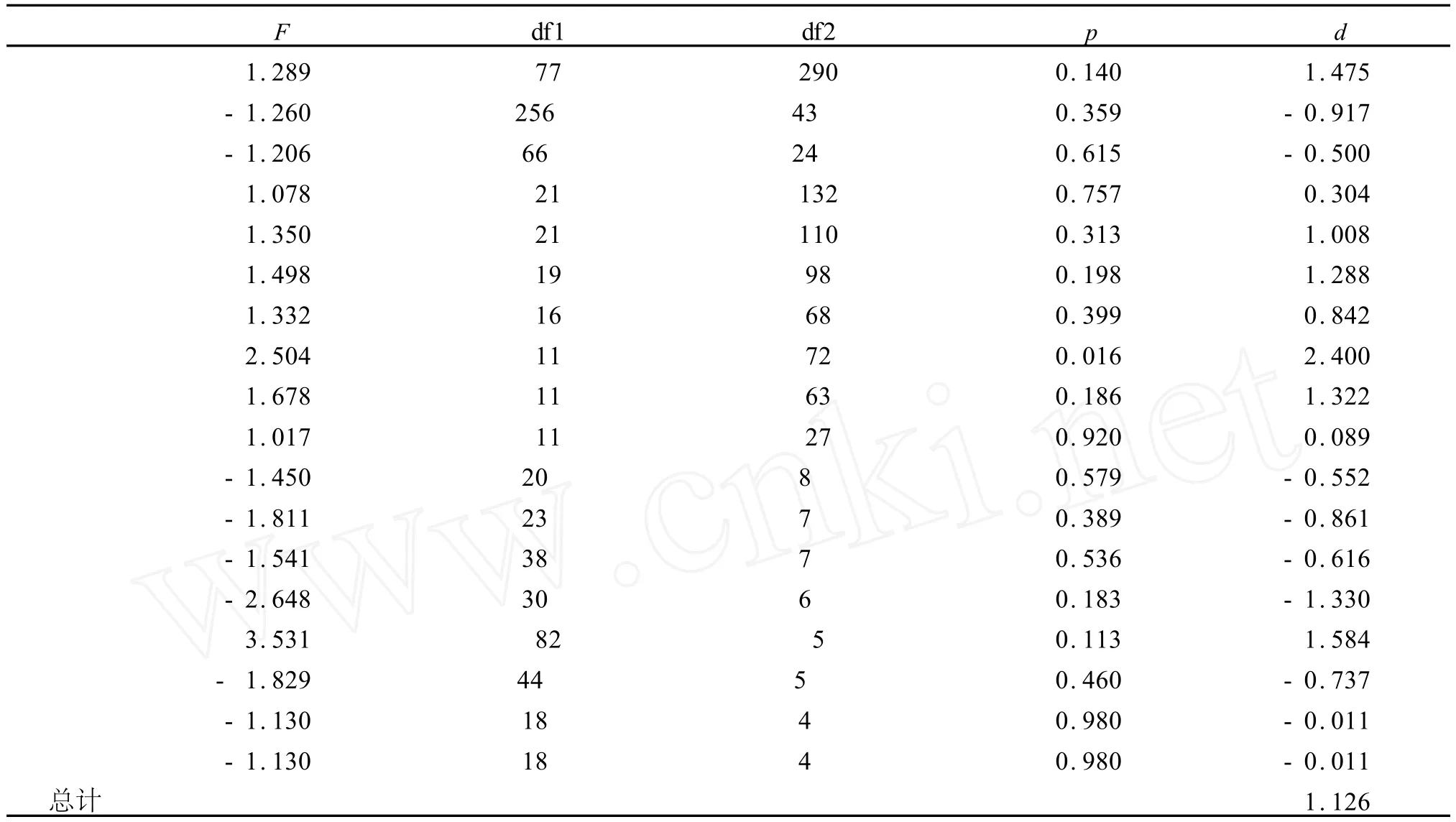
表1 封闭之下Cepaea nimora lis两种壳颜色壳宽度的F值Table 1 Shell breadth in two colour classesof Cepaea nimoralis Cepaea nimoralis,enclosure samples
2 结果
将封闭蜗牛群体资料分析结果列于表1中,如果黄色个体的方差比粉色个体的大,则F是正值,如果黄色的方差小,则F值是负的.最后一列给出了正态离散值,它相当于在一定样本大小下的F值.当这些数值结合在一起,总的正态离差值是1.126,次值不显著,同样方法也用在了爱尔兰蜗牛三维度量总方差上,对三种不同类型正态离差比较以及样本数目列于表2中,如果对颜色表型比较,由81个样本比较得正态离差是1.732(P<0.84,二尾;0.042,一尾).通过壳上斑纹有无而比较 59个样本得正态离差是 1.448(P<0.148,二尾;0.074,一尾).由此我们可以说,最大的差异应该通过黄色有斑纹的纯合子与其他非黄色表型的比较来看到.按此,我们对76个样本做了比较其正态离差是1.423,非常接近斑纹和无斑纹比较的数值.如若我们按颜色将两方面资料加以组合,所得表型离差是2.021,这个数值是显著的(P<0.043,二尾;0.022,一尾),见表2所示.

表2 爱尔兰 Cepaea nemoralis总变异的正态离差和或然率Table 2 Overall normal deviates and p robabilities for total variance of Irish Cepaea nimoralis
对全部资料,壳宽度和高度的相关系数是0.987;壳宽度与壳口宽度的相关系数是0.988;高度与壳口宽度相关系数是0.983.在具斑纹或无斑纹壳之间比较壳三维测定的相关系数,53个样本中,有18个总离差是正值,5个显著.测定离差总趋向值是0.032,指出没有一类性状表现出更大相关性.
3 讨论
Cepaea nemoralis种群在壳颜色和壳斑纹上总是多态性的.有关选择影响形态频率的途径被发现了许多[2~4],但是多态性是如何维持下来的一直未能得到解答.一种合乎情理的解释是由于多态性是可见的(visible),维持它延续的选择是可见的环境因子(p redator),大多数是勉力频率依赖性选择(exerting frequencydependent selection)[18].这种观点也可由类似的适应色的事实支持,比如鳞翅目中的拟态(mimicry)[19].文章开始提到Goodhard的假设认为可见的选择是基因位点组合的平衡而杂合基因组具有更高适合性(fitness)看见多态性是总杂合系统一部分的可能性应引起注意.本研究就是为了验证这种假设.
总离差在所有情况下均是正值,这就给出一些证据说明在壳颜色和式样上基因的变异,隐性基因变异要比杂合和显性纯合基因要大.对颜色组合分析结果在一尾和二尾都是显著的,当然这种显著性度并不是很强.另一方面,分析指出在两性状之间、两种度量之间没有相关性.为了获得更有力的证据来说明这种相互关系,需要更多收集蜗牛壳.同时现阶段研究表明,更进一步研究这个问题是值得的.然而,为了获得更有力证据来说明真正在形态上变异上的不同,就需要观察比我们现在所观察的5 000个蜗牛壳还要更多,这样就可以研究来自不同生境的蜗牛之间的遗传异质性.
当然,生物统计学上的巨大变异并不一定意味着壳的颜色或式样的不适应.如果多态性一直在一地保持足够长的时间,并且这种维持是在可见表型频率依赖性选择上[3],这或许是由连锁基因位点操纵着,这种连锁起着增加杂种优势.Cepaea nemoralis和C.hortensis是在更新世时期冰层向北退缩时的优势种,它们在壳的颜色上具有巨大的多态性,与它们有相互关联的南方种C.sylvatica和C.vindobonensis是在壳斑纹上具多态性.这是否表明足够时间跨度使北方种发展了一超基因系统(supergene system)来显示杂合子优势[20].
[1] Cook L K.The genetic of cepaea nemoralis[J].Heredity,1967,22:397-410
[2] Cain A J.Ecology and ecogenetics of terrestrialmolluscan populations[M].New York:Academic Press,1983:579-647
[3] Clarke B C,A rthur W,Horsley D T,et al.Genetic variations and natural selection in pulmontemolluscs[M].New York:Academic Press,1978:219-270
[4] Jones J S,Leith B H,Raw lings R.Poymorphism in Cepaea:a p roblem with too many solutions?[J].Ann.Rew.Ecol.Syst,1977,8:109-143
[5] Goodhart C B.Why are some snails visibly polymorphic and others not?[J].Biol.J.Linn.Soc.,1987,31:35-58
[6] Mitton JB,Grant M C.Associations among protein heterozygosity,grow thra te and developmental homeostasis[J].Rev.Ecol.Syst.,1984,15:479-499
[7] Palmer A R,Strobeck C.Fluctuating asymmetry measurement,analysis,paterns[J].Ann.Rev,Ecol.Syst.,1986,15:391-421
[8] Zouros E.On the relation between heterozygosity and heterosis:an evalution of the evidence from marine mollusks[J].Isozymes:Curr.Top.Biol.Med.Res.1987,15:255-170
[9] Beaumont A R.Genetic studies of labo rato ry reared mussels mytilus edulis:heterozygote deficiencies,heterozygousity and grow th[J].Biol.J.Linn.Soc.1991,44:273-285
[10] Zouros E,Foltz D W.The use of allelic isozyme variation for the study of heterosis[J].Isozymes:Curr.Top.Biol.Med.Res.1987,13:1-59
[11] Koehn R K.Diehil W J,Scott TW.The differential contribution of indiviual enzymesof glycolysis and p rotein catabolism to the relationship between heterozygosity and grow th rate in the coot clam,M ulinia lateral-is[J].Genetics,1988,118:121-130
[12] Foote M,Cow ie R H.Developmental buffering as amechanism for stasis:evidence from the pulmonate Theba pisana[J].E-volutiOn,1988,42:396-399
[13] Clarke B C.The evidence for apostatic selection[J].Heredity,1969,24:347-353
[14] Burke D P T.Variation in body colour in western Irish populations of Cepaes nemoralis(L)[J].Biol.J.Linn Soc.1989,36:55-63
[15] Davies R G.Computer p rogramming im quantitive biology[M].London:Academ ic Press,1971
[16] Heteling H.New light on the correlation coefficient and its transform s[J].J.R.Stastist,B,1953,15:193-232
[17] Sokal R R,Rohlf F J.The p rinciple and p ractice of statistics in biological research[M].Freeman,San Francisco:Biometry,1969
[18] Clarke B C.The evidence for apostatic selection[J].Heredity,1969,24:347-353
[19] Clarke C A,Sheppard PM.Further studieson the geneticsof mimetic butterfly papilio memnom(L)[J].Phil.Trans.R.Soc.B.,1971,263:35-70
[20] Cook L M,Cain A J.Population dynamic,shell size and morph frequency in experimental populationsof snail Cepaea nemo ralis(L)[J].Biol.J.Linn.Soc.,1980,14:259-292
Study of the Relation of M orphological Variation and Heterozygosity in the Snail Cepaea Nemoralis
Gao Guixi
(Department of Biology,Taiyuan Normal University,Taiyuan 030001,China)
It has been suggested that the polymo rphism for shell co lour and pattern in Cepaea may be unrelated to the visible appearance of the shells,butmaintained by heterosisor heterozygote advantage.A possible way to test this supposition is to examine whether phenotypic variance is lower in themorph exhibiting the dominant phenotype than in that with the recessive phenotype.This has been tested in two samplesof shells involving totalsof 1 792 and 5 423 individuals,respectively.Overall,there is a significantly greater variance in the recessive than the dom inant class(P<0.05).Correlation between measurements on the same shell was also examined.No evidence of difference between phenotypes was found.If the difference in variance is real,it may indicate either of the follow ings:1.that visible appearance is not important and the polymorphism ismaintained by a heterozygous balance;or 2.evolution of such a balance in a polymorphism maintained over a long time spanby selection acting on visible attributes of the shells.
heterozygosity;phenotypic variance;polymorphism;relational balance;cepaea
【责任编辑:王映苗】
1672-2027(2010)02-0130-04
Q 89;Q 3
A
2010-03-25
山西省留学基金资助.
高贵喜(1961-),男,山西岢岚人,太原师范学院生物系副教授.
猜你喜欢
阅读与作文(小学低年级版)(2021年3期)2021-05-07
种子(2021年3期)2021-04-12
趣味(数学)(2021年11期)2021-03-09
中外文摘(2019年2期)2019-01-30
中国生育健康杂志(2018年6期)2018-11-13
中学时代(2018年7期)2018-08-24
消费导刊(2017年24期)2018-01-31
辽宁大学学报(哲学社会科学版)(2017年3期)2017-06-21
外语教学理论与实践(2016年1期)2016-06-11
中学生理科应试(2016年7期)2016-05-14
