
常见石莼科绿藻的生物学特征及分子系统学分析*
2010-01-05 08:20王绪敏宫庆礼
中国海洋大学学报(自然科学版) 2010年12期
刘 岩,刘 涛,于 丹,张 静,王绪敏,宫庆礼**
(1.中国海洋大学海洋生命学院,山东青岛266003;2.中国科学院北京基因组研究所,北京100029)
常见石莼科绿藻的生物学特征及分子系统学分析*
刘 岩1,刘 涛1,于 丹2,张 静1,王绪敏2,宫庆礼1**
(1.中国海洋大学海洋生命学院,山东青岛266003;2.中国科学院北京基因组研究所,北京100029)
生物学研究及种类鉴定是绿潮研究的热点和难点之一。本文系统阐述了石莼科绿藻常见种类的生物学特征,探讨了系统分类研究难点,并利用ITS、rbc L等基因序列以及质体蓝素氨基酸序列进一步研究分析了石莼科10种绿藻的序列结构特征和分子系统学关系。研究显示,石莼科浒苔属和石莼属绿藻rbc L和ITS序列差异未体现出属间分化的差异,根据rbc L序列和ITS序列分别构建的系统进化树中,石莼和孔石莼、肠浒苔、扁浒苔都聚类到同一进化枝上,而裂片石莼和网石莼、浒苔和缘管浒苔聚类到另一进化枝上,这显示出在分子进化水平上较为一致的结果,为浒苔属和石莼属系统分类深入研究提供一定的证据;根据质体蓝素所构建的系统进化树中,阿氏石莼和孔石莼在NCB I上发表的质体蓝素氨基酸序列在第1进化枝中,浒苔的质体蓝素氨基酸序列在第2进化枝中。但在第1进化枝中,孔石莼的3个质体蓝素氨基酸序列没有完全分布在同一个分支上。
石莼属;浒苔属;分类;ITS;rbc L
大型绿藻异常增殖现象越来越多地出现在美洲、亚洲和欧洲各国暖水性海域,这种因海水富营养化和异常气候条件引起的生态学现象引起了广泛地重视,为了与海洋微藻和小型浮游生物形成的赤潮(红潮,Red tides)相区别,将其称为绿潮(Green tides)。根据目前的报道,绿潮生物主要集中在绿藻门(Chlorophyta)石莼目(U lvales)石莼科(U lvaceae)的石莼属(U lva)和浒苔属(Enterom orpha)、刚毛藻目(Cladophorales)刚毛藻科(Cladophoraceae)硬毛藻属(Chaetomorpha)及刚毛藻属(Cladophora)。其中,以石莼属和浒苔属大型绿藻为常见绿潮生物种类。自20世纪末期以来,绿潮研究的重点除了发生机制及其规律的探讨外,其对绿潮生物的鉴定和种群区分则是另外一个研究的热点和难点[1-4]。本文在石莼科海藻常见海藻生物学特征的基础上,将综合有关分子学鉴定证据,进一步研究和探讨其分类、鉴定工作。
1 材料与方法
1.1 序列资料
选取GenBank收录的9种石莼科海藻的ITS、rbcL等基因序列和3种石莼科海藻的质体蓝素氨基酸序列资料,其中ITS序列8条、rbcL序列9条、氨基酸序列6条(见表1、表2)。
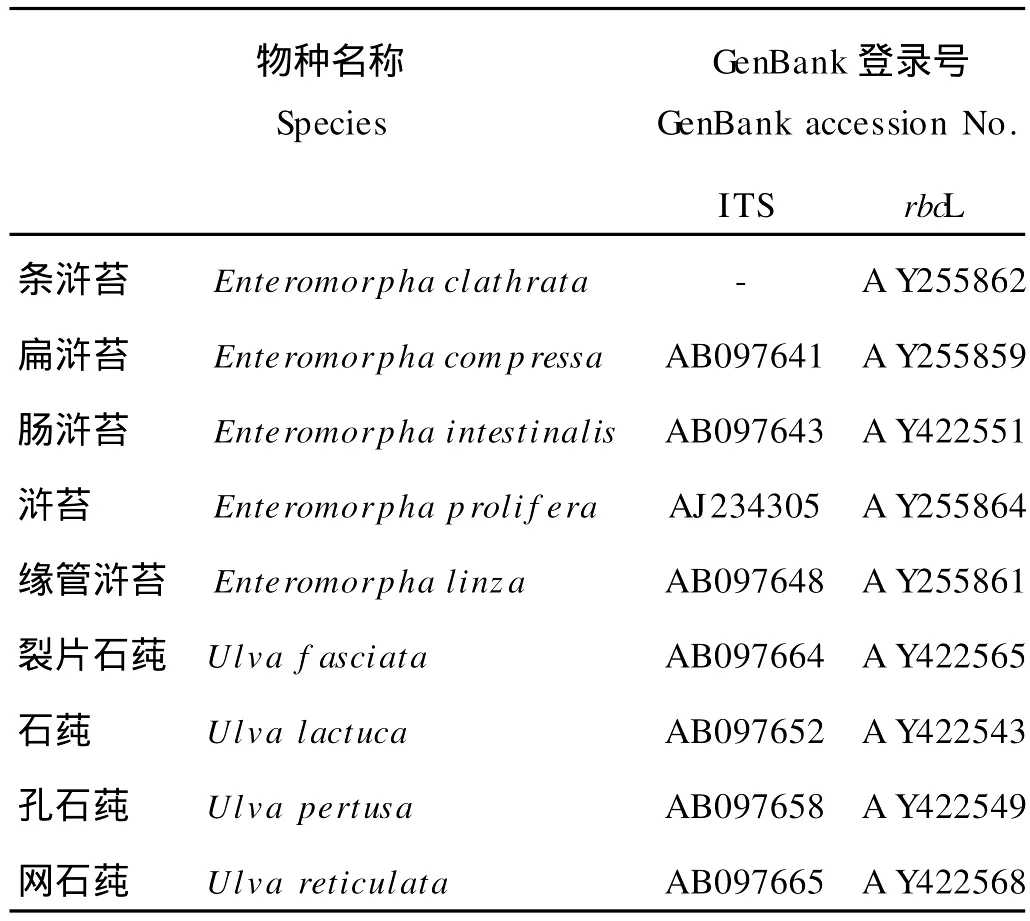
表1 石莼科海藻的ITS序列和rbc L序列Table 1 Sequences of ITS and rbc L of Ulvaceae

表2 石莼科海藻的质体蓝素序列Table 2 Sequences of plastocyanin of U lvaceae
1.2 分析软件
所涉及的分析软件包括:DNASTAR和M EGA 4。计算机运行环境为window s XP。
2 结果
2.1 ITS序列分析
DNASTAR软件分析的结果(见表3),浒苔属和石莼属的ITS序列的C+G%值在62.2%~65.4%之间,其中ITS1的C+G%值在65.4%~70.0%之间,ITS2的C+G%值在67.2%~74.8%之间,5.8s rDNA的C+G%值在48.1%~49.4%之间。可以看出,浒苔属和石莼属的ITS序列的C+G%值偏高。
用DNASTAR软件分别对浒苔属和石莼属的ITS1、5.8s rDNA、ITS2序列分别进行同源性分析(见表4~6)。
浒苔属和石莼属海藻5.8s rDNA序列高度同源,各物种间的同源性在97.5%~100%之间,长度都为160 bp,其中浒苔、缘管浒苔、裂片石莼、网石莼的5.8s rDNA序列完全相同。
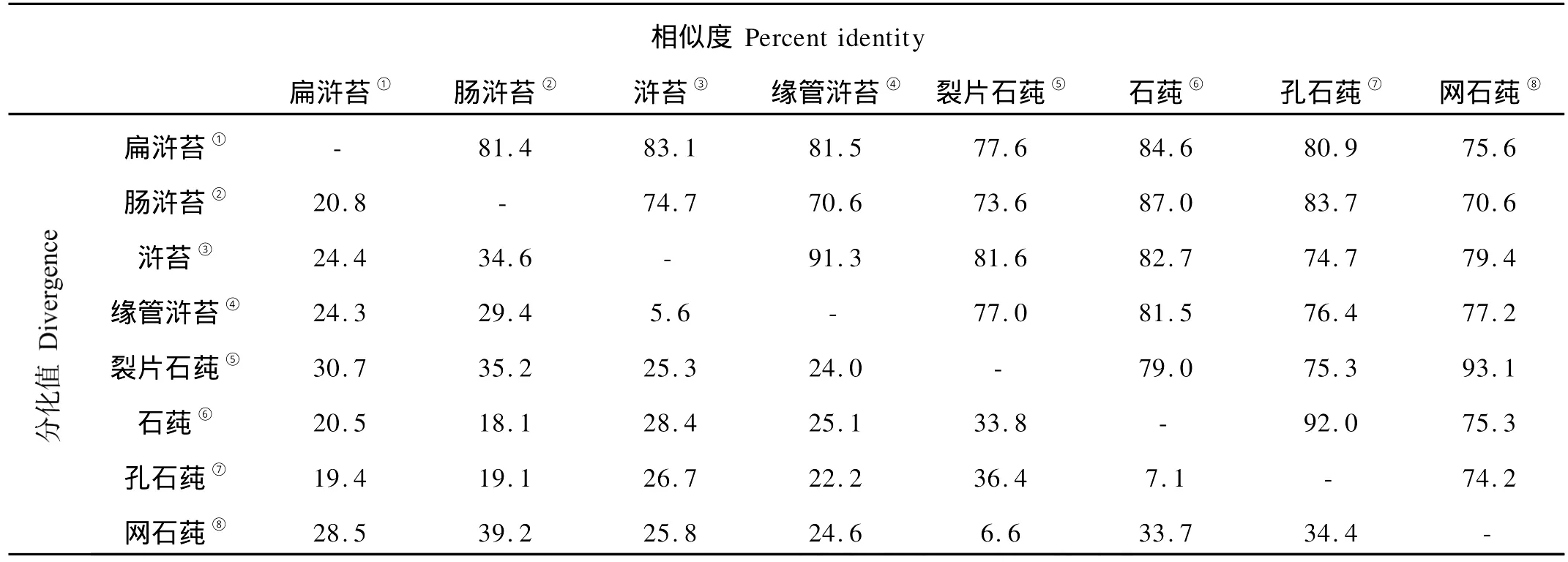
表5 浒苔属和石莼属ITS1序列同源性比对结果Table 5 Homology alignment of ITS1 between U lva and Enteromorpha
浒苔属和石莼属的ITS1序列的同源性在70.6%~92.0%之间,其中,缘管浒苔、浒苔的同源性为91.3%,石莼、孔石莼的同源性为92.0%。其它的物种之间的同源性没有表现出明显的属间差异,即浒苔属和石莼属各个物种的ITS1序列在属间的同源性与属内的同源性数值相近。
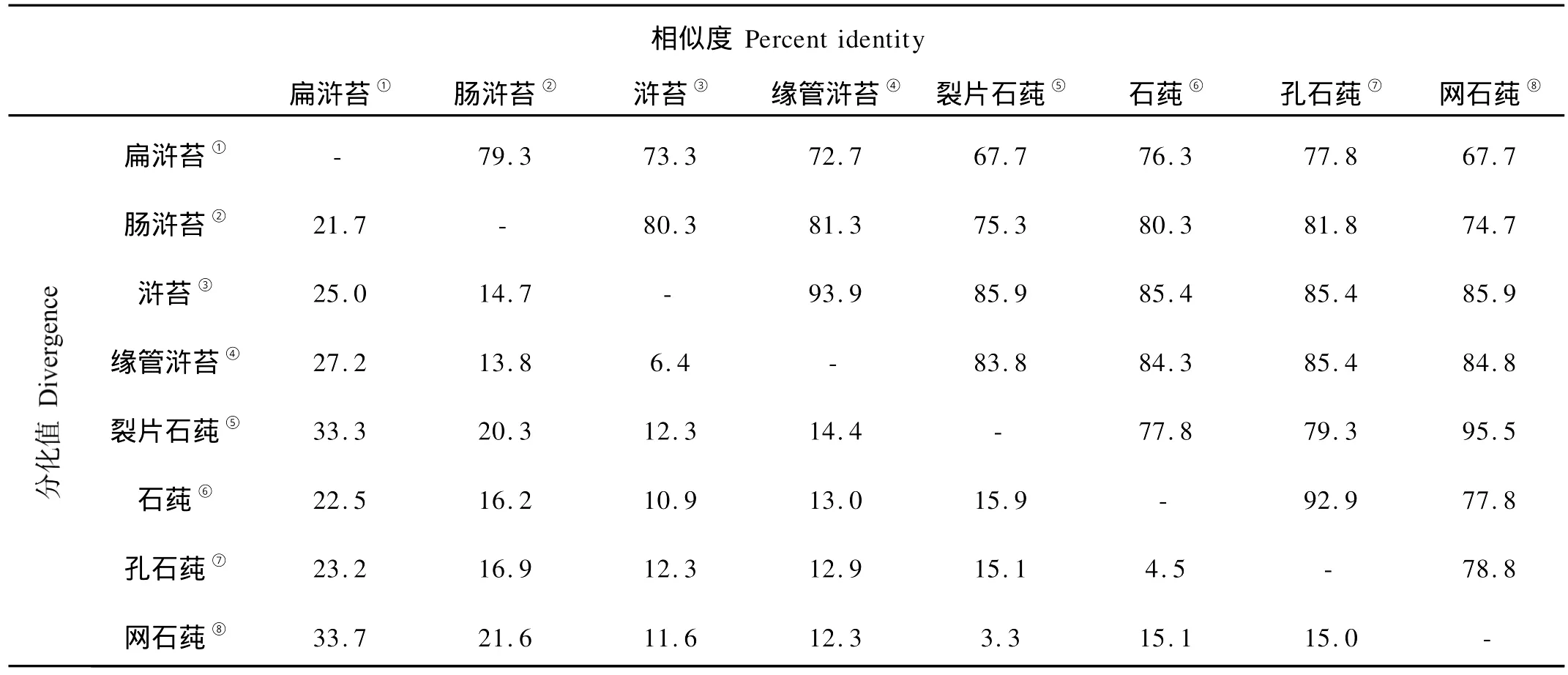
表6 浒苔属和石莼属海藻ITS2序列同源性比对结果Table 6 Homology alignment of ITS2 between Ulva and Enteromorpha
浒苔属和石莼属的ITS2序列的同源性在67.7%~95.5%之间,其中裂片石莼、网石莼的同源性为95.5%,缘管浒苔、浒苔的同源性在93.9%,石莼、孔石莼的同源性在92.9%,扁浒苔、裂片石莼的同源性为67.7%。浒苔属和石莼属其他物种的ITS2序列间的同源性与其ITS1序列的同源性比对结果相近。
以基因序列研究生物的遗传距离和系统进化关系时,实质上是进行其碱基差异及其排列方式的统计分析。因此,序列长度及其保守性(变异率)将直接影响到最终的分析结果。因为浒苔属和石莼属个别物种的ITS序列差异较小,且浒苔属和石莼属海藻的5.8S rDNA序列高度保守,因此,在对ITS序列进行遗传距离分析时,去除5.8S rDNA序列,用M EGA 4软件计算浒苔属和石莼属海藻的ITS1和ITS2序列的遗传距离(见表7)。
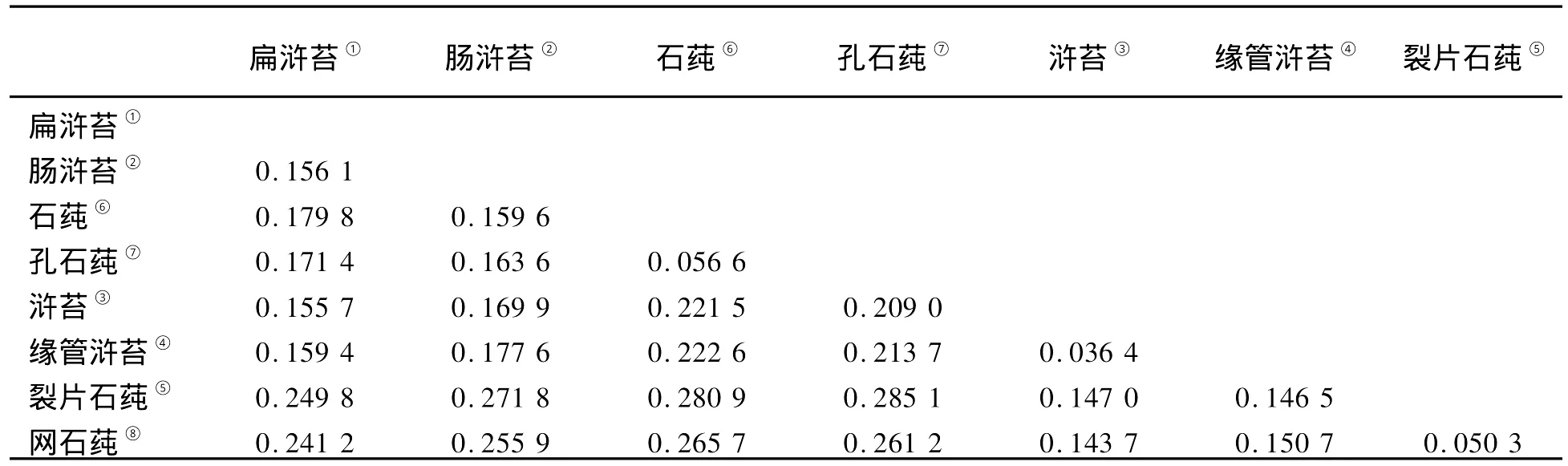
表7 浒苔属和石莼属海藻ITS1和ITS2序列遗传距离Table 7 Pairw ise distances of ITS1 and ITS2 between U lva and Enteromorpha
以ITS1、ITS2序列为依据,缘管浒苔、浒苔的遗传距离为0.036 4,裂片石莼、网石莼的遗传距离为0.050 3,石莼、孔石莼的遗传距离为0.056 6,以上3组遗传距离相对较近;裂片石莼、孔石莼的遗传距离为0.285 1,裂片石莼、石莼的遗传距离为0.280 9,以上2组遗传距离相对较远。从以上数据中可以看出,以ITS1、ITS2序列为依据,浒苔属和石莼属各个物种间的遗传距离分布混乱,未表现出明显的属间特异性。
用M EGA 4软件,采用NJ分析法,以ITS1、ITS2序列对浒苔属和石莼属各个物种建立系统树(见图1)。
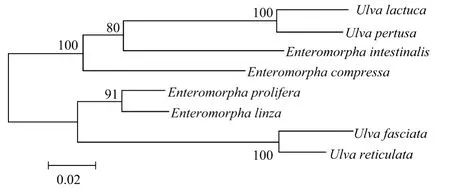
图1 浒苔属和石莼属海藻ITS序列系统发育进化树Fig.1 Phylogenic tree of ITSof U lva and Enteromorpha
2.2 rbc L序列分析
DNASTAR软件分析的结果(见表8),浒苔属和石莼属的rbc L序列的C+G%值在38.26%~39.14%之间,序列的C+G%值较低。
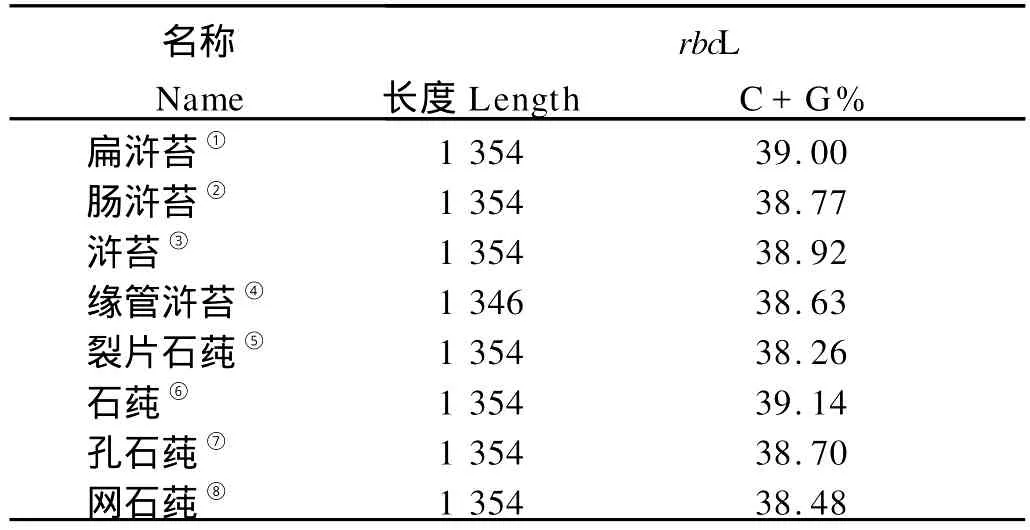
表8 浒苔属和石莼属rbc L序列长度及C+G%值Table 8 Lengths and values C+G%of rbc L of U lva and Enteromorpha
用CLASTAL X软件对浒苔属和石莼属海藻rbc L基因的核苷酸序列进行比对,并用M EGA 4软件,以rbc L基因的核苷酸序列,计算浒苔属和石莼属海藻的遗传距离(见表9)。
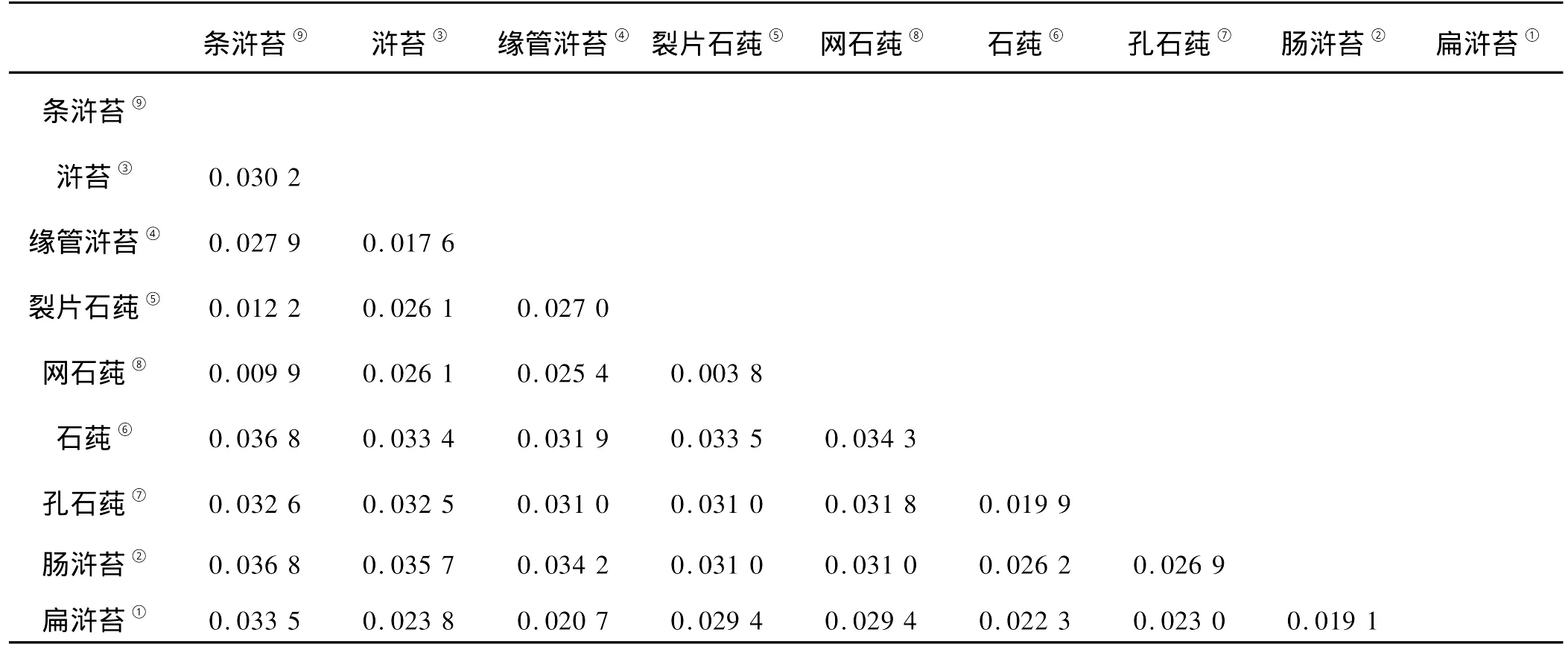
表9 浒苔属和石莼属海藻rbc L基因核苷酸序列遗传距离Table 9 Pairw ise distances of rbc L
以rbc L基因核苷酸序列为依据,网石莼、裂片石莼的遗传距离最近,为0.003 8。网石莼、条浒苔的遗传距离为0.009 9,裂片石莼与条浒苔的遗传距离为0.012 2,这2组的遗传距离相对较近;石莼、条浒苔以及肠浒苔、条浒苔之间的遗传距离值最大,为0.036 8。其它物种之间的遗传距离数值没有表现出明显的以现有分类的属为单位聚类的情况。同时,以rbc L基因的核苷酸序列为依据计算出的浒苔属和石莼属各个物种的遗传距离数值要明显小于以ITS序列为依据计算出的遗传距离数值。
用M EGA 4软件,采用NJ分析法,以rbc L基因核苷酸序列进一步对浒苔属和石莼属海藻建立系统树(见图2)。
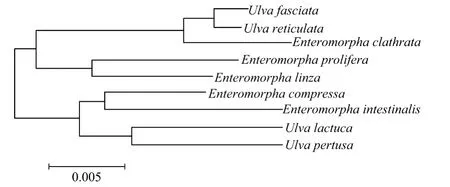
图2 浒苔属和石莼属海藻rbcL序列系统发育进化树Fig.2 Phylogenic tree of rbcL of Ulva and Enteromorpha
2.3 质体蓝素序列分析
用CLASTAL X软件对浒苔属和石莼属海藻质体蓝素的氨基酸序列进行多重序列比对,并用M EGA 4软件计算浒苔属和石莼属海藻的遗传距离(见表10)。
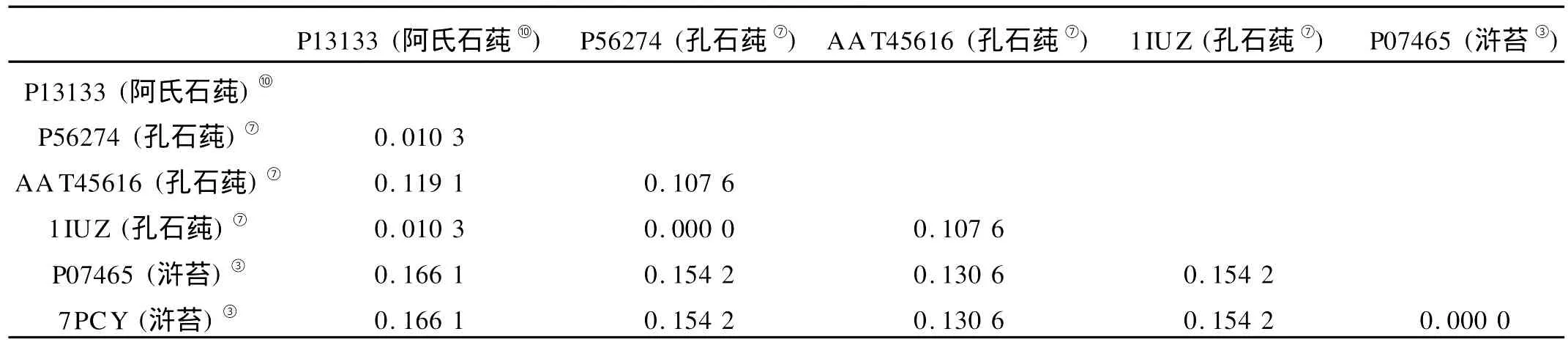
表10 浒苔属和石莼属海藻质体蓝素序列遗传距离Table 10 Pairw ise distances of p lastocyanin of U lva and Enteromorpha
以质体蓝素的氨基酸序列为依据,孔石莼的1IUZ和P56274的遗传距离为0.000,浒苔的P07465和7PCY的遗传距离为0.000,浒苔的P07465和阿氏石莼的P13133的遗传距离最大,为0.166 1。
用M EGA 4软件,采用NJ分析法,以质体蓝素的氨基酸序列进一步对浒苔属和石莼属海藻建立系统树(见图3)。
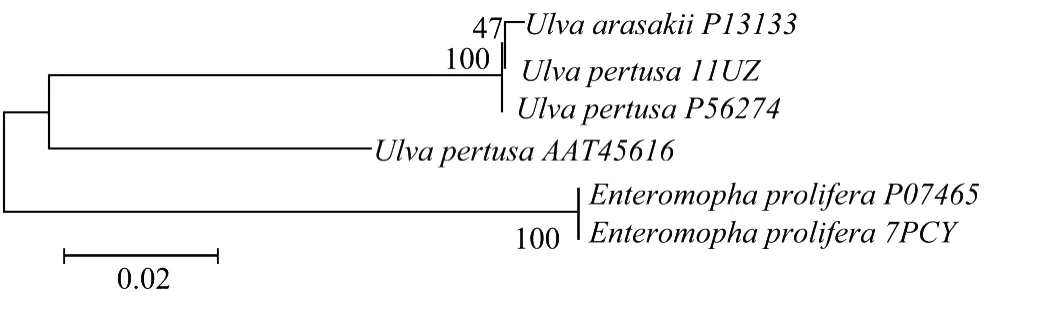
图3 浒苔属和石莼属海藻质体蓝素序列系统发育进化树Fig.3 Phylogenic tree of plastocyanin of Ulva and Enteromorpha
3 分析与讨论
3.1 石莼科藻类的生物学特征
石莼科(U lvaceae)海藻的藻体呈叶片状或为中空圆柱状,由1层或2层细胞组成,叶状体的构造根据不同的种、属而不同。细胞单核,有1个侧生色素体,色素体内含1至多个淀粉核,藻体基部细胞延伸成固着器。无性生殖时,产生游孢子,孢子囊产生具4根鞭毛的游孢子,成熟后经母细胞壁的开孔逸出,游孢子萌发成具有假根的丝状体或不规则的细胞,再进一步发育成叶状体。有性生殖时,产生具双鞭毛的配子。多数物种异配生殖,只有在不同藻体产生的配子间才能接合,合子立刻萌发成新藻体。石莼科藻类的生活史多数为同形世代交替,孢子体和配子体具有相同的形态特征。孢子体上营养细胞形成游孢子囊,核经过减数分裂,产生游孢子,游孢子萌发成配子体;配子体产生配子,配子接合成二倍体的合子,由合子再发育成孢子体。石莼科的大多数物种分布于沿海潮间带岩石上,少数种生于咸淡水或淡水中。
3.1.1 石莼属海藻的生物学特征及其分类 石莼属(U lva)海藻藻体为多细胞叶状体,由2层细胞组成,基部由营养细胞延伸成假根丝,形成固着器,固着于岩石上。细胞内有1个细胞核及1个杯状色素体,其中含有淀粉核。生活史为双元同形。无性生殖过程中,由孢子体边缘的营养细胞开始形成孢子囊,孢子囊在形成的过程中,边缘细胞叶绿体移往细胞的一边,同时细胞向外生出小突起,细胞第一次分裂时的分裂面与叶状体表面垂直,为减数分裂;第二次分裂面与第一次垂直,分成4个细胞,如此继续分裂,每一孢子囊产生8~16个游孢子。孢子成熟后由囊上突起小孔逸出,游孢子具4根鞭毛,离开母体后,游动片刻,即附着在岩石上,失去鞭毛,分泌细胞壁,1~2 d内开始萌发为配子体。有性生殖为异配生殖,由配子体形成配子囊,配子的形成与游孢子相似,但每一配子囊产生16~32个配子,成熟的配子亦由囊上突起的小孔逸出。配子离开母体后,进行异配接合,结合后的合子2~3 d内开始萌发,长成孢子体。配子体有时也能进行孤雌生殖。
我国近海石莼属常见种类有以下4种:石莼(U.lactuca),孔石莼(U.pertusa),裂片石莼(U.fasciata)和砺菜(U lva.cong lobata)[5]。
(1)孔石莼(U.pertusa)藻体幼期绿色,成体为碧绿色,体形变异很大,有卵形、椭球形、披针形和圆形等,但都不规则。边缘略有皱或稍呈波状。藻体叶面常有大小不等不甚规则的穿孔,并且随着藻体长大,几个小孔可裂为一个大孔,最后使藻体形成几个不规则的裂片状。藻体高10~40 cm。固着器盘状,柄不明显。藻体单独或2~3株丛生,中实,基部较厚。藻体横切面观,细胞纵长方形、角圆,长为宽的2~3倍。边缘细胞为亚方形,长宽相似或略高,体厚70μm左右或以上。上部体厚70μm,边缘常较薄,藻体下部逐渐增厚,约130~180μm,基部约500μm。细胞含有1个细胞核,1个大型色素体。孔石莼为北太平洋西部特有的暖温带性海藻,生长在中潮带及低潮带和大干潮线附近的岩石上或石沼中。
(2)石莼(U.lactuca)藻体近似卵圆形、宽叶状,边缘略有波状。藻体黄绿色,藻体长10~30 cm,藻体边缘中实,切面观细胞形状为亚方形,体厚45μm左右。石莼为泛温带型种,生活在海湾内、中潮带及低潮带的岩石或石沼中。
(3)砺菜(U.cong lobata)自藻体基部边缘向基部深裂,形成许多裂片相互重叠呈花形。藻体鲜绿色,茂密丛生,藻体长2~4 cm,略扩展呈团块状。藻体边缘中实,边缘扭曲,切面观细胞形状为长方形、角圆,上部及边缘部的细胞长度与宽度相同或略大于细胞宽度,下部细胞随着藻体的增厚,细胞增大,细胞壁稍厚,细胞形态呈棱柱状,细胞腔长度为宽度的1.5~2倍。藻体上部为薄膜质,上部体厚30~50μm、下部体厚100~125μm,基部稍硬。砺菜为北太平洋西部特有暖温带性海藻,生长在中潮带和高潮带岩石或石沼的边缘。
(4)裂片石莼(U.fasciata):不规则二叉式分枝,形成或多或少的线形裂片。藻体草绿色,藻体长15~60 cm,藻体边缘中实,叶片边缘平滑或具有不规则的齿状突起,有时呈波状。藻体的基部略宽,向上分叉1~2次,并逐渐窄细。切面观细胞形状为长方形,细胞壁较厚,体厚70~80μm。裂片石莼为亚热带性海藻,生长在风浪较小的大干潮线附近的岩石或低潮带的石沼中。
3.1.2 浒苔属海藻的生物学特征及其分类 浒苔属(Enteromorpha)海藻绿色,藻体单条或有分枝,圆管状中空,有时部分稍扁。藻体无柄,成熟时从基部细胞生出假根丝形成盘状固着器固着在基质上。藻体壁由1层细胞组成。细胞内有1个细胞核和1个片状色素体,位于原生质体的表面,一般含有1个淀粉核。浒苔属藻类生活史为双元同形。有性生殖过程中,由配子体产生2根鞭毛的配子,配子放散后经异配接合为合子,合子萌发,直接发育成新个体。配子有时进行单性生殖。浒苔属海藻也可进行营养生殖,由藻体细胞直接进行有丝分裂形成分枝体,基部分生的细胞逐渐分化为假根,从藻体脱离后形成新的植株。浒苔属约有40种,主要生活于海水和半咸水中,也可生活在含盐的内陆水体中。
我国近海浒苔属常见种类有条浒苔(E.clathrata),肠浒苔(E.intestinalis),扁浒苔(E.com pressa),管浒苔(Enterom orpha.tubulosa),曲浒苔(Enteromorpha.Flexuosa),浒苔(E.prolifera)和缘管浒苔(E.linza,也称长石莼[U.linza])等[5]。
(1)肠浒苔(E.intestinalis)藻体管状中空,部分稍扁,单条或基部有少许分枝,高10~20 cm;直径1~5 mm。单生或丛生,体表面常有许多皱褶或藻体扭曲。柄部圆柱形,上部膨胀如肠形。除基部细胞稍呈纵列外,藻体其他部分的细胞排列不甚规则。细胞表面观直径10~23μm,圆形至多角形,细胞内有1个杯状色素体,内含1个淀粉核,横切面观偏于单层藻体的外侧。全年各月都能生长和繁殖。体厚16~39μm。肠浒苔为泛冷温带性种,生长在海湾内潮间带的岩石上或石沼中,淡水流入处也可生长。
(2)浒苔(E.prolifera)藻体呈暗绿色或亮绿色,藻体长可达1~2 m,宽可达2 cm。管状或扁压,有明显的主枝,多细长分枝或育枝,分枝的直径小于主干。柄部逐渐尖细,藻体基部或至少在分枝基部的细胞排列成纵列,但上部细胞不纵列或纵列不明显。细胞表面观直径16μm,圆形至多角形,细胞内一般含1个淀粉核,有时多个,色素体不充满。藻体厚15~18μm,可达26μm。切面观细胞在单层藻体的中央。浒苔为温带性海藻,生长于中潮带的石沼中。
(3)条浒苔(E.clathrata)藻体亮绿色或暗绿色,藻体长40 cm左右,多细长分枝,一般分枝2~3回,有时分枝呈毛状或较宽,分枝顶端细胞单列或多列,在同一藻体中可看到单列枝与多列枝,多列枝顶端有1~12个单列的细胞。藻体自基部到顶端细胞纵列。细胞表面观直径较大,方形或长方形,长20μm左右,宽30~50μm,细胞内淀粉核2~3个,色素体充满或不充满,有时在同一藻体上,主干色素体不充满,分枝色素体充满。体厚26~70μm。切面观细胞在单层藻体的中央。条浒苔为泛暖温带性海藻,多生长于中潮带、内湾泥底、潮间带洼地中。
(4)扁浒苔(E.com pressa)藻体亮绿色,藻体长可达50 cm。藻体有许多分枝,基部分枝较密,上部较疏。分枝基部略为缢缩,上部较粗大,分枝的形状和直径与主干相似,藻体上部扁压。藻体除基部外,细胞不排成纵列。细胞表面观直径10~27μm,细胞圆形至多角形,细胞壁薄,细胞内有1个淀粉核,色素体不充满。藻体厚8~20μm。切面观细胞在单层藻体的中央。扁浒苔为泛暖温带性海藻,多生长于中、低潮带的石砾、岩石或石沼中。
(5)管浒苔(E.tubulosa)藻体深绿色,藻体长度6~18 cm,细长单条,偶尔在基部有小分枝。藻体下部管状或近似圆柱形,上部有时略为扁压,自基部到顶端细胞整齐纵列,有时呈横列。细胞表面观直径11~15μm,方形或长方形,细胞内有1个淀粉核,色素体不充满。藻体厚23μm左右。切面观细胞在单层藻体的中央。管浒苔为泛亚热带性海藻,多见于内湾,特别是半咸水,生长于潮间带的石砾、岩石或各类物体上。
(6)曲浒苔(E.f lexuosa)藻体单条,圆柱状或管状,藻体长2 cm以上,有明显的柄部,向上逐渐膨胀如肠形。细胞长13~17μm,宽10~17μm,多角形,角圆,少数长方形,排成纵列,细胞壁厚,细胞内有1个淀粉核,色素体充满。藻体厚20~24μm。切面观细胞在单层藻体的中央或靠外侧。曲浒苔为泛亚热带性海藻,多生长于高、中潮间带的岩石或石沼中。
(7)缘管浒苔(E.linza),也称长石莼。藻体绿色,簇生.细长单条,体长为5~30 cm,体宽为0.2~3 cm,体厚约为50μm。藻体不分枝或自基部分裂出形态相同的分枝,藻体基部狭窄细长,中部渐宽,呈带状,藻体表面细胞正方形、长方形或多边形,细胞排列成纵列。自基部到顶端细胞整齐纵列,切面观,藻体中央呈2层结构,边缘部中空。
3.2 浒苔属和石莼属海藻的分类学探讨
浒苔属和石莼属海藻作为石莼科的分类阶元具有同形世代交替,孢子生殖、配子生殖和营养繁殖等共性特征。其属间鉴定主要是根据藻体是否中空(即管状或叶状)或成体细胞层数进行区分;属内的不同物种,则根据藻体藻体分枝数量或叶片是否分裂、细胞直径和排列方式、细胞内淀粉核数量等进行鉴别。
由于在不同环境条件下,石莼科海藻藻体形态及结构存在着广泛的变异,因此对其系统分类一直存在较多的争议。尤其是近年来,根据石莼科海藻的分子系统学研究的结果显示,以ITS序列和rbc L基因序列构建的系统发育进化树,浒苔属和石莼属海藻相互交错分布,因此,有人认为应把这2个属的海藻都合并在石莼属中[6-8]。但根据刘振宇等对孔石莼、U.arasakii和浒苔的质体蓝素氨基酸序列系统进化关系的研究结果来看,浒苔亲缘关系还是要远于原为石莼属的2种海藻[9]。
缘管浒苔形态结构和分子鉴定结果则进一步加强了浒苔属和石莼属之间的联系。缘管浒苔藻体基部和边缘呈中空管状,由单层细胞构成,类似浒苔属海藻;藻体的其余部分无管状结构,形成了由2层细胞构成的藻体,与石莼属海藻相近。1753年,Linnaeus最早发现该种时,将其定名为长石莼(U.linza L innaeus),其后瑞典分类学家Agardh(1883)根据特征将其转入到浒苔属内,命名为缘管浒苔[E.linza(Linnaeus)J Agardh],2种观点在很长一段时间同时存在,如Levring等许多学者同意Linnaeus命名为长石莼的意见,但也有不少学者认同Agardh将其分类到浒苔属的观点[10-12]。
3.3 浒苔属和石莼属海藻的分子进化
生物进化内在动力是基因变异,通过比对分析石莼科海藻的基因序列,可以探讨其在分子水平的进化差异,验证形态分类研究结果。
3.3.1 浒苔属和石莼属海藻ITS序列和5.8s rDNA序列的进化发育关系探讨 rDNA是由转录单位和非转录单位的间隔区组成的一个重复单位,其中18,5.8和28s rDNA组成1个转录单元。而非转录单位包括3个部分(见图4):①非转录间隔区(Non transcribed sequence N TS)位于rDNA相邻重复单位之间;②外转录间隔区(Extra transcribed sequence,ETS)位于18s rDNA的上游(Upstream);③内转录间隔区(Internal transcribed spacers,ITS)包括ITS1和ITS2,分别位于18s rDNA和5.8s rDNA,5.8s rDNA和28s rDNA之间。有时将ITS1,5.8s rDNA和ITS2统称为ITS区[13-14]。
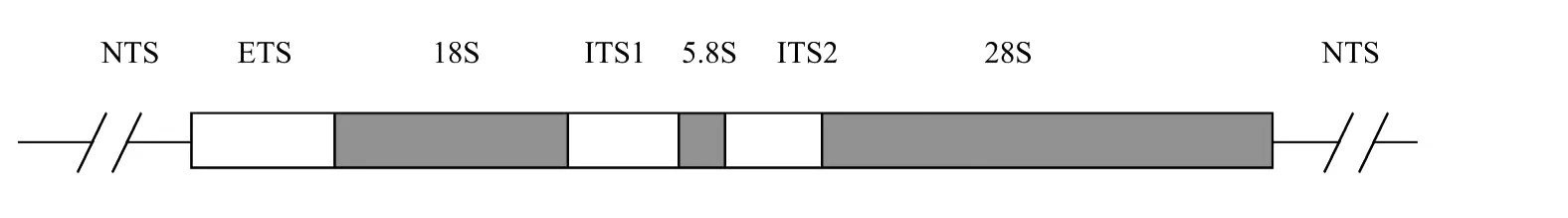
图4 真核生物rDNA一个拷贝的结构Fig.4 Structure of a repeating unit eukaryote rDNA
由于ITS存在于高重复的核糖体DNA中,进化速度快且片段长度不大,加上协调进化使该片段在基因组不同重复单元间十分一致,因而十分适合于进行各种分子操作,已成为植物系统与进化研究中的重要分子标记。
浒苔属和石莼属的5.8s rDNA序列仅在4位点上出现碱基的突变,其中,第1和第4个突变位点为浒苔属特有的变异位点,第2个突变位点为石莼属特有的变异位点,第3个突变位点为2者共有的变异位点。分别对出现碱基突变的物种进行种内的序列比对,从结果中可以得出,这些位点的突变是存在于不同物种之间的,并不是由测序的误差或个体差异造成的。说明,这些变异位点是在物种进化中形成的,具有物种特征。但同时也反映出,这些变异的位点并非在属这一分类阶元上是完全一致的,其实质上是不同物种在平行进化过程中对环境压力的适应。
在ITS序列系统发育进化树中,浒苔属和石莼属的各个物种分为2大进化枝。在第一进化枝中,石莼、孔石莼聚类在一起。在第二进化枝中,缘管浒苔、浒苔和裂片石莼、网石莼分别聚类在一起。从2个进化枝中的物种组成可以看出,浒苔属和石莼属的物种并没有完全的按照各自的属分别聚类在一起,而是交叉的分布在2个进化枝中。
ITS序列系统进化聚类以及5.8s rDNA序列比对结果说明,利用上述序列差异无法进一步将这2个属海藻明确地进行鉴别,同时,以ITS和5.8s rDNA序列进行缘管浒苔和浒苔,裂片石莼和网石莼鉴定时,其碱基位点差异极小或相同,因此在研究过程中需要加大测序的样本数量以保证测序和拼接结果的准确性。
3.3.2 浒苔属和石莼属海藻rbc L基因序列的进化发育关系探讨 核酮糖-1,5-双磷酸羧化酶/加氧酶(Rubisco)是叶绿体基质中的主要可溶性蛋白,此酶在光合作用的Calvin—Benson循环中固定CO2,而且也在C3植物的光呼吸过程中起作用。它由8个相同的大亚基和8个相同的小亚基组成。大亚基由叶绿体DNA编码并且由叶绿体的核糖体合成;小亚基由核基因编码。首先由细胞质内的核糖体合成前体蛋白,穿越叶绿体膜后再加工为成熟形式[15]。编码大亚基的基因简称为rbc L基因,编码小亚基的基因简称为rbcS基因。
高等植物rbc L基因在结构上和原核生物基因相似,由5’非编码区、编码区和3’非编码区3部分组成。5’非编码区具有可以和叶绿体16S rRNA 3’端附近互补的SD序列;3’非编码区具反向重复序列,能形成典型的茎环结构作为转录终止信号;多数rbc L基因还具有和原核生物基因启动子类似的共通序列[16-17]。
从浒苔属和石莼属海藻rbc L序列系统发育进化树中可以看出,浒苔属和石莼属的各个物种分为2大进化枝。此2个进化枝中的物种组成以及各个物种的分支情况都与以ITS序列为依据对浒苔属和石莼属建立的系统树情况相似。即浒苔属和石莼属的物种并没有完全的分别按照各自的属聚类在一起,而是交叉的分布在2个进化枝中。
综合分析浒苔属和石莼属海藻以rbc L和ITS序列为依据建立的系统树,以rbc L和ITS序列为依据,不能将浒苔属和石莼属中的各个物种鉴别开来;以目前发表的rbc L和ITS序列为依据,现有分类的浒苔属和石莼属中的各个物种间没有表现出明显属间的差异,且从系统树中可以看出,属间物种有明显的交叉情况。且从rbc L和ITS序列分别聚类的结果来看,石莼和孔石莼、肠浒苔、扁浒苔都聚类到同一进化枝上,而裂片石莼和网石莼、浒苔和缘管浒苔也聚类到另一进化枝上。这显示出在分子进化水平上较为一致的结果,将为浒苔属和石莼属系统分类深入研究提供一定的证据。
3.3.3 浒苔属和石莼属海藻质体蓝素序列的进化发育关系探讨 质体蓝素(Plastocyanin)是含有铜原子的蛋白质,普遍存在于包括蓝细菌、绿藻和高等植物在内的能进行有氧光合作用的物种中,具有从光合系统Ⅱ传递电子到光合系统I的重要功能。质体蓝素包含近100个氨基酸残基的多肽链以及I型铜离子(也称铜蓝位点);该铜离子结合在多肽链的4个氨基酸残基(即2个组氨酸残基、1个半胱氨酸残基和1个甲硫氨酸残基[18-20])上。
阿氏石莼、孔石莼、浒苔分为2大进化枝,其中,阿氏石莼和孔石莼在NCB I上发表的质体蓝素氨基酸序列在第1进化枝中,浒苔的质体蓝素氨基酸序列在第2进化枝中。但在第1进化枝中,孔石莼的3个质体蓝素氨基酸序列没有完全分布在同一个分支上,P56274和1IUZ聚类在一起,并且,从比对结果中可以看到,AA T45616与前2个序列存在10个氨基酸序列的差异,这可能是因为其材料来源的不同造成的。
以目前浒苔属和石莼属已公布的质体蓝素氨基酸序列来看,阿氏石莼、孔石莼、浒苔的聚类结果符合现有的浒苔属和石莼属的ITS、rcb L分子系统学研究结果,但是,现有发表的浒苔属和石莼属的质体蓝素氨基酸序列的物种较少,尚不足以说明其是否广泛适用于浒苔属和石莼属间各物种的分类鉴定。
在海藻物种的分类和鉴定方面,主要依据藻体的形态、结构、生殖方式、生活史类型等性状特征进行分类,尽管不同的海藻物种所持有的这些性状特征是遗传基因所决定的,但其在不同海洋环境条件下显示出了广泛的适应能力,表现为环境因素过多地影响到了表型特征。不仅在单细胞的微型藻类方面存在着巨大的争议,即使是在大型藻类常见种鉴定方面也出现了一些激烈的争议。二十世纪末期随着分子生物学技术和基因测序技术快速发展,藻类分子系统学的研究结果往往显示出经典分类学上的对物种甚至属一级定义的差异,这似乎更为充分地证实了藻类生物平行进化复杂机制。但从分子系统学研究手段本身而言,也存在很多需要进一步研究和探讨的问题。不同类群同一DNA序列的进化速率有所差异,同一类群中不同DNA序列的进化速率也不相同,因此应针对不同分类阶元选取相应的DNA序列;生物进化实质上是基因组的进化,因此,少数基因片段无法全面、准确地描述在物种水平上的系统发育关系和反映其演化历史。藻类具有核、线粒体和叶绿体3套基因组,从多个层次以及多个基因位点上系统地研究藻类形态结构进化、生态适应以及基因进化的内在关系则显得更为重要和迫切。
致谢:本文石莼属和浒苔属形态分类部分惠承中国科学院海洋研究所丁兰平研究员审订。
[1] Elina L,Pekka P.Evolution of the ITS sequences of ribosomal DNA in Enteromorpha(Chlorophyceae)[J].Hereditas,1997,126:17-23.
[2] Jaanika B,Christine M,Michael S.Molecular and mo rphological analysis of Enteromorpha intestinalis and E.com pressa(Chlorophyta)in the British isles[J].J Phycol,1998,34:319-340.
[3] J.Blom ster E M Hoey,C A Maggs.Species-specific oligo-nucleotide p robes fo rmacroalgae:molecular discrimination of two marine fouling species of Enteromorpha(Ulvaphyceae)[J].Molecular E-cology,2000,9(2):177-186.
[4] Inger A,Anders B.Structure and function of Rubisco[J].Plant Physiology and Biochemistry,2000,8,46:275-291.
[5] 钱树本,刘东艳,孙军,等.[M]青岛:中国海洋大学出版社,2005.
[6] Tan I H,Blomster J,Hansen G,et al.Molecular phylogenetic evidence for a reversible morphogenetic switch controlling the grossmorphology of two common genera of green seaweeds,U lva and Enteromorpha[J].Mol Biol Evol,1999,16(8):1011-1018.
[7] Shimada S,Hiraoka M,Nabata S,et al.Molecular phylogenetic analyses of the Japanese Ulva and Enteromorpha(Ulvales,Ulvophyceae),with special reference to the free-floating Ulva[J].Phycol Res,2003,51:99-108.
[8] Hayden H S,Blomster J,Maggs C A,et al.Linnaeus was right all along:Ulva and Enteromorpha are not distinct genera[J].Eur J Phycol,2003,38:277-294.
[9] 刘振宇,谢荔岩,吴祖建,等.孔石莼质体蓝素氨基酸序列分析和分子进化[J].分子植物育种.2005,3(2):203-208.
[10] 曾呈奎,张德瑞,张峻甫.中国经济海藻志[M].北京:科学出版社,1962.43.
[11] 杨君,安利佳,王茜,等.石莼属(U lva)和浒苔属(Enteromorpha)绿藻的RAPD分析[J].海洋与湖沼,2003,31(4):408-413.
[12] Tseng.Common seaweeds of China[M].Beijing:Science Press,1983.
[13] 邱芳,伏建民.遗传多样性的分子检测[J].生物多样性,1998.6:143-150.
[14] 陈琳琳,孔晓瑜,周立石,等.魁蚶核糖体DNA基因转录间隔区的序列特征[J].中国水产科学,2005,12(1):104-107.
[15] Shinozaka K,Sugiura M.The nucleotide sequence of the tobacco chloroplast gene for the large subunit of ribulose-1,5-bisphosphate carboxylase/oxygenase[J].Gene,1982,20:91-102.
[16] Masahiro S.The chloroplast genome[J].Plant Molec Biol,1992,19:149-168.
[17] 王艇,苏应娟,朱建明.叶绿体rbc L基因序列在植物系统学研究中的应用[J].武汉植物学研究,1999,17(增刊):8-14.
[18] Matthew R,Todd Y,Sabeeha M.Plastocyanin:Structural and functional analysis[J].Journal of Bioenergetics and Biomembranes,1994,26(1):49-66.
[19] Shibata N,Inoue T,Nagano C,et al.Novel insight into the copper-ligand geometry in the crystal structure of U lva pertusa plastocyanin at 1.6-resolution[J].J Biol Chem,1999,274(7):4225-4230.
[20] Bertini I,Bryant D A,Ciurli S,et al.Backbone dynamics of plastocyanin in both oxidation states[J].J Biol Chem,2001,276(50):47217-47226.
Biological Characteristics and Molecular Systematics Studies on Common Green Algae of Ulvaceae
L IU Yan1,L IU Tao1,YU Dan2,ZHANG Jing1,WANG Xu-M in2,GONG Qing-Li1
(1.College o f Marine Life Sciences,Ocean University of China,Qingdao 266003,China;2.Beijing Genomics Institute of Chinese Academy of Sciences,Beijing 100029,China)
Biological research and species identification is one of the hotspo ts and difficulties of green tides study.This paper describes the biological characteristicsof common green algae of U lvaceae and discusses the difficulties of systemic classification.ITS sequences,rbc L sequences and p lastocyanin amino acid sequences were used to analyze the molecular systematics of 10 green algae of Ulvaceae.Studies show ed that rbcL and ITS sequences between Ulva and Enteromorpha didnot reflect intergeneric differentiation.According to the phylogentic trees constructed respectively by rbc L sequences and ITS sequences,U lva lactuca,U lva pertusa,Enteromorpha intestinalis and Enteromorpha com pressa were clustered to the same evolutionary branch and Ulva fasciata,Ulva reticu lata,Enteromorpha prolifera and Enteromorpha linza were clustered to another,indicating consistent results at the molecular evolutionary level.It also p rovided evidence for further research between U lva and Enteromorpha.According to the phylogentic tree constructed by p lastocyanin am ino acid sequences,the sequence of U lva arasakii and the sequences of U lva pertusa were clustered to the first evolutionary branch and the sequences of Enterom orpha prolifera were clustered to the second.Three sequences of U lva pertusa were clustered to different evolutionary branches.
Ulva;Enteromorpha;classification;ITS;rbcL
Q944
A
1672-5174(2010)12-071-10
青岛市科技项目(09-2-5-5-hy);国家科技支撑计划项目(2008BAC49B01)资助
2009-10-26;
2010-01-10
刘 岩(1979-),女,讲师,博士生,研究方向为藻类学。Tel:0532-82032377;E-mail:qd_liuyan@ouc.edu.cn
**通讯作者:gongqingli@vip.sina.com
责任编辑 于 卫
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
广西科学(2022年1期)2022-04-12
作文周刊·小学四年级版(2022年8期)2022-03-11
中国果菜(2021年4期)2021-05-09
中国兽医杂志(2020年10期)2020-03-04
——有效的抗弓形虫药物靶标
中国预防兽医学报(2020年8期)2020-01-15
小学生必读(低年级版)(2019年5期)2019-08-30
水产科学(2019年4期)2019-07-23
食品与生活(2017年12期)2018-01-09
科技创新与应用(2017年20期)2017-07-15
