
河北秦皇岛仙菜科Ceramiaceae红藻的分类研究*
2022-04-12 11:21:48刘美媛闫盼竹王雪聪王艺晓黄冰心丁兰平
广西科学 2022年1期
刘美媛,闫盼竹,王雪聪,闫 璟,张 瑶,王艺晓,楚 越,黄冰心,丁兰平
(天津师范大学生命科学学院,天津市动植物抗性重点实验室,天津 300387)
秦皇岛地处渤海西部,沿海岸岩礁与沙质底质交错,为渤海西部优质的大型海藻分布区。然而,针对该区域大型红藻的研究至今仅有零星报道[1-5]。仙菜科Ceramiaceae是重要的海洋大型红藻类群,在沿海岸水域生态系统中具有重要的作用和良好的应用价值[6]。仙菜科隶属红藻门仙菜目Ceramiales,由Dumortier[7]于1822年建立,目前中国已报道23属[6,8-11]。本科的主要特征:藻体为单轴型,极少数为多轴型,无皮层或部分具皮层或全具皮层。顶端生长,由顶端细胞横向或纵向分裂发育为一列中轴细胞。具皮层的种类,其皮层的发生最初在节部产生一圈围轴细胞,有的种类其围轴细胞不再分裂延伸,有的延伸到节间部分,形成节与节间均具皮层的状态。四分孢子囊有柄或无柄;精子囊生长于特殊分枝上,往往集合成簇,有的散生于皮层内;成熟囊果裸露或被总苞枝包围。
仙菜科红藻大部分物种因个体微小脆弱、形态多变[12]、常附生和不易肉眼分辨等,导致其物种分类上还存在许多不足[13],部分物种甚至仅有记录式描述,缺乏具体的特征描述。针对这些问题,本文对采自我国河北秦皇岛地区的仙菜科红藻进行分类研究,以期阐明该水域仙菜科的物种多样性,为其物种资源的保护和利用提供科学依据。
1 材料与方法
1.1 材料
以2019-2021年采自河北秦皇岛秦皇求仙入海处和山海关老龙头两地的仙菜科红藻样品为实验材料,共计30个样品,包括液浸标本(用5%甲醛海水溶液浸泡)、腊叶标本及冻存标本(液氮冷冻后于-20℃冰箱中保存),分别编号后保存于天津师范大学生命科学学院藻类实验室。
1.2 方法
1.2.1 外部形态观察
用普通光学显微镜和体式 Nikon SMZ25 显微镜观察藻体颜色并测量藻体大小,确定藻体分枝方式,是否全部具有皮层;观察顶端、基部特征,是否存在假根、四分孢子囊、囊果和精子囊等;对关键鉴定特征进行拍照并保存。
1.2.2 内部结构观察
选择样品的特征性部位,生物冷冻切片后,用亚甲基蓝溶液或0.5%苯胺蓝染液加1%盐酸对切片进行染色,并在荧光正置显微镜 Leica DM5000B 下观察、拍照。
1.2.3 解剖特征观察
选择藻体的生殖器官及内部结构等进行观察,包括藻体围轴细胞数量、皮层初始细胞排列方式及数量、内外皮层细胞大小及形态等,对关键鉴定特征进行拍照并保存。
1.2.4 物种鉴定
参考专业文献,根据样品的形态与解剖学特征对分类特征进行比对,鉴定物种[14]。
2 结果与分析
2.1 波登仙菜[图1:(a)-(m)]
CeramiumboydeniiGepp[15],1904,p.164,pl.460;Tseng[16],1983,p.126,pl.66,fig.4;郑柏林等[10],2001,p.34-35,fig.18;丁兰平等[11],2015,p.180.
描述:藻体深红色,质地较硬,通常丛生呈团状,附生,高6-15 cm。藻体顶部钳状,顶端较大幅度内弯,见图1:(a)、(g)。基部一些小枝缠绕在固着物上并用假根附着,见图1(c)。藻体在同一平面内呈假二叉分枝,主轴上每隔8-16个轴细胞发生分枝,侧轴上每隔9-17个轴细胞发生分枝,见图1:(a)、(b)。在老的藻体上有单生、对生或轮生小分枝,见图1(b)。轴细胞椭圆至圆柱状,见图1(f),围轴细胞8个,见图1(h),所有围轴细胞都产生皮层细胞。藻体全部具皮层,见图1:(d)、(e),分内外两部分,内层细胞由一层椭圆形细胞组成,外皮层由2-3层小球形细胞组成,见图1:(i)、(j)。囊果球形,具5-6瓣裂总苞,生长于枝末端或次末端,大小为(380-420) μm×(350-380) μm,见图1:(k)、(l)、(m)。
样品中没有发现雄性藻体。
模式标本产地:中国,日本。
国内地理分布:辽宁大连、旅顺,河北秦皇岛,山东烟台、青岛,浙江南麂岛。
样品编号:20190512006,20190512010和20210517020,采自河北秦皇岛山海关老龙头;20210518003,采自河北秦皇岛秦皇求仙入海处。
2.2 三叉仙菜[图2:(a)-(n)]
CeramiumkondoiYendo[17],1920,p.9;郑柏林等[18],1961,p.234;Nakamura[19],1965,p.155,fig.14;Boo et Yoon[20],1993,p.179-189,figs.1,5-6;RAN et al[21],1997,p.27-32,fig.1.
描述:藻体玫瑰红色,高6-30 cm,表面光滑。藻体上部直,具不等长分枝,见图2:(a)、(b),顶部内弯,呈弯钩状,见图2(c)。基部有盘状固着器固着于基质上。主轴直立,见图2(d),上有部分小枝,见图2(e),主要呈三叉分枝,存在少数二叉分枝和四叉分枝,见图2:(f)、(g)、(h)。具明显的节和节间,皮层完全,见图2(i)。藻体中心是一列轴细胞,呈圆柱状,在分枝顶端具一顶端细胞,由其产生围轴细胞。围轴细胞8个,见图2(k),较大,近椭圆或多角状。围轴细胞分生出皮层细胞,包围轴细胞,组成皮层。皮层细胞1-3层,近椭圆形,较小,排列紧密,见图2(j)。成熟囊果侧生,无柄,生于雌性藻体分枝或小枝的次末端或生育枝的末端,见图2(l),单个或成列,具4-5个总苞,直径60-100 μm,长140-650 μm,见图2:(m)、(n)。

(a)、(b)藻体上部;(c)藻体顶部;(d)藻体轴;(e)小分枝;(f)-(h)二叉分枝,三叉分枝,四叉分枝;(i)皮层细胞;(j)、(k)轴横切面显示皮层细胞和围轴细胞;(l)雌性藻体;(m)、(n)囊果枝
样品中没有发现雄性藻体。
模式标本产地:日本。
国内地理分布:辽宁大连,河北秦皇岛,山东烟台、青岛,浙江南麂岛。
样品编号:20190512003,20190512004,采自河北秦皇岛山海关老龙头;20190511008,20210330001,20210516010,20210518002,采自河北秦皇岛秦皇求仙入海处。
2.3 孙氏仙菜(新拟名)[图3:(a)-(q)]
CeramiumsungminbooiJ.R.Hughey & G.H.Boo[22]2016,p.216,figs.1-5.
描述:藻体呈玫瑰红色或红色,浓密且呈扇形,生境中聚集呈球形,高3-8 cm,见图3(a),藻体直立,呈假二叉分枝,见图3:(b)、(c)、(h)。具明显的节和节间,仅在节处有皮层,见图3:(d)、(e),主轴上有部分钳状小枝,见图3:(d)、(f),顶端稍弯曲,见图3:(c)、(i)。围轴细胞5个,见图3(q),围轴细胞分生出皮层细胞,于围轴细胞外部组成皮层。皮层由2-3层皮层小细胞组成,见图3:(o)、(p)。四分孢子囊直径30-40 μm,从分枝的围轴细胞正面突起且强烈突起,见图3:(j)、(k)、(l)、(m),顶端分枝由7-8个细胞组成,见图3(n)。
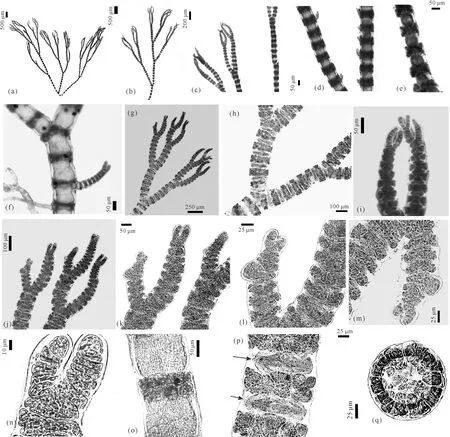
(a)藻体整体;(b)、(c)藻体上部;(d)、(e)藻体直立枝表面观;(f)小分枝;(g)藻体顶部;(h)藻体假二叉分枝;(i)藻体顶端分枝;(j)四分孢子囊枝;(k)-(m)四分孢子囊细节;(n)藻体顶端分枝显示为7-8个细胞;(o)藻体皮层表面观;(p)轴细胞及皮层细胞,箭头指示轴细胞;(q)轴横切面显示5个围轴细胞
样品中没有发现生殖藻体。
模式标本产地:韩国。
国内地理分布:河北秦皇岛。本种为我国新纪录种。
样品编号:20210329014,20210517020,采自河北秦皇岛山海关老龙头;20210516015,采自河北省秦皇岛秦皇求仙入海处。
2.4 多姿对丝藻[图4:(a)-(g)]
AntithamniondefectumKylin[23,24],1925,p.46,fig.27;1956,p.348,fig.276B;Smith[25],1944,p.75;Abbott et Hollenberg[26],1976,p.573;刘剑华[27],1984,p.40-47,pl.2,figs.1-5;郑柏林等[10],2001,p.13-14,fig.6;丁兰平等[11],2015,p.180.
描述:藻体暗红色,纤细,直立,丝状,高0.5-2.5 cm,见图4(a)。主轴由匍匐基部产生,主轴上的侧枝无小枝对生,见图4(b)。主轴和侧枝的细胞上部具有对生小枝。小枝细胞基本等径,见图4:(c)、(d)、(g)。小枝除基部方形细胞外,向轴面产生篦状羽枝,见图4(d)。藻体顶端小枝密集,外形呈圆锥形,藻体顶部的小枝和篦状羽枝细胞较粗短,枝端细胞较钝,见图4(f);下部小枝和篦状羽枝细胞细长,顶端尖细,见图4:(c)、(d)。主轴细胞长110-205 μm,宽40-100 μm,长为宽的2-3倍,见图4:(b)、(c);小枝细胞长55-75 μm,宽20-25 μm;腺细胞位于小枝基部第二或第三个细胞生出的短小篦状羽枝上,介于这种枝末端的2-5个细胞内侧,长25-35 μm,宽15-20 μm,见图4:(c)、(d)、(g)。精子囊丛生在小枝向轴面的精子囊枝上,见图4(e)。

箭头处为腺细胞;(a)藻体整体;(b)藻体下部及主轴侧枝;(c)对生小枝及腺细胞;(d)蓖状羽枝;(e)精子囊枝;(f)藻体顶端;(g)小枝细节
模式标本产地:美国华盛顿。
国内地理分布:山东青岛,河北秦皇岛。本种为渤海沿岸的新纪录种。
样品编号:20210330014,采自河北秦皇岛秦皇求仙入海处。
2.5 绢丝藻[图5:(a)-(i)]
Callithamnioncorymbosum(Smith)Lyngbye[28],1819,p.125,tab.38c;刘剑华[6],1979,p.25-37,pl.2,figs.1-4.
ConfervacorymbosaSmith,1812,pl.2352[29].
描述:藻体鲜红色,纤细,质地柔软,丛生成簇,高1.5-4.5 cm,见图5(a),常附生于其他藻体上。藻体上部呈伞房状,分枝密集,见图5(b),顶部细胞钝圆,常具有早落性的无色毛,见图5:(f)、(g)。藻体轴由单列细胞组成,基部有许多分枝根样丝,附着于基质上,根样丝细胞大小为(40±5) μm×(140±9) μm,见图5(d)。直立枝明显,呈互生分枝,见图5(c),每一分枝呈二叉分枝,见图5:(e)、(f)、(g),直立枝细胞大小为(58±6) μm×(160±20) μm,分枝细胞大小为(23±3) μm×(70±5) μm,见图5:(f)、(g)。四分孢子囊卵形,无柄,生于藻体上部小枝内侧近基部,单生或2-3个集生,见图5:(h)、(i)。

(a)藻体标本;(b)藻体下部;(c)直立枝上的互生分枝;(d)匍匐枝顶端根样丝;(e)匍匐枝中部;(f)藻体顶端区域细胞;(g)匍匐枝细胞;(h)、(i)四分孢子囊
样品中没有发现生殖藻体。
模式标本产地:大西洋北部。
国内地理分布:河北秦皇岛,山东青岛,福建厦门和平潭,海南西沙群岛。
样品编号:20210516012,20210516013,20210516016,20210518001,采自河北秦皇岛秦皇求仙入海处。
3 讨论
仙菜属由Roth于1797年建立[30],本属广泛分布于世界各地,是红藻门中最大的属之一[9,10,12,31]。传统分类根据皮层、顶端结构、分枝模式、是否存在不定枝、围轴细胞的数量、四分孢子囊和囊果位置等来区分物种[19,22,32]。
孙氏仙菜Ceramiumsungminbooi为我国新纪录种,其最为明显的特征是四分孢子囊从分枝的围轴细胞正面产生且强烈突起,这在仙菜属中比较少见。孙氏仙菜与C.cimbricumH.E.Petersen相似,主要区别在于C.cimbricum围轴细胞为6-7个,而孙氏仙菜的围轴细胞为5个[9,21,33]。Kim[34]报道C.cimbricum分布于韩国,Gittenberger等[35,36]也报道了荷兰瓦登海18个地方分布的“C.cimbricum”,但不能确定是否为本地种。Hughey等[22]则认为,瓦登海样品很可能是来自亚洲的孙氏仙菜。
波登仙菜Ceramiumboydenii在我国黄海、渤海及东南沿海均有生长,比较常见。报道于国外的CeramiumriosmenaeB.Y.Won et T.O.Cho、C.interruptumSetchell et Gardner、C.sinicolaSetchell et Gardner和本种类似,区别在于C.riosmenae的藻体和囊果均比波登仙菜的小得多[37]。C.interruptum和波登仙菜的主要区别为前者皮层不完全[38]。C.sinicola个体明显小于波登仙菜,且分枝间的间隔轴细胞个数也远远小于波登仙菜。
三叉仙菜Ceramiumkondoi在我国沿海均有分布,多见于辽宁大连、河北秦皇岛、山东青岛及浙江嵊山等海区,该藻从春季到夏季在沙石上或作为附生植物在其他藻类上大量生长[39]。此外,还常见于日本[17,19]、韩国[20]和俄罗斯远东地区[40]。该种分枝模式和皮层化程度多变,包括4个变型——f.abbreviatumNakamura、f.ambiguumNakamura、f.kondoiNakamura和f.trichotomumNakamura[19,41,42],但也存在争议[39]。该种与C.virgatumRoth和C.rubrumC.Agardh相似,但C.virgatum具有全皮层的轴[15,19,20]。该种也常与C.rubrum混淆,后者是一种著名的世界性藻类物种[39,43]。据Nakamura[41]报道,一些被鉴定为三叉仙菜的日本标本可能因其具有二叉分枝而被命名为C.rubrum,而一些被鉴定为C.rubrum的欧洲标本也可能被命名为三叉仙菜。Garbary等[44]发现,加拿大地区的C.rubrum没有明显的三叉分枝。Boo等[20]根据对三叉仙菜分枝模式的全年观察,认为三叉分枝方式是该种在分类学上的有效分类特征,可明确区分三叉仙菜和C.rubrum。三叉仙菜与波登仙菜在外形上也极为相似,区别在于三叉仙菜的老枝上无轮生小枝,而波登仙菜存在[10]。
对丝藻属Antithamnion由Nägeli于1847建立[45],分布在北极和南极地区以及亚热带至热带沿海植物区[46]。其主要特征为主轴丝状,由单列细胞组成,主轴和侧枝的细胞上部具有对生小枝,小枝的基部细胞近方形。该属的物种常通过轮生小枝的形态、腺细胞小枝的形态和位置、不定枝端部果胞期分枝的数量和不定侧枝的位置来区分[47]。以往在确定对丝藻属物种时,部分区分特征有时似乎被过分重视[46]。Gardner[48]指出,A.defectum、A.setaceumN.L.Gardner及A.pygmaeumN.L.Gardner十分相似,表现在小枝和篦状羽枝的形态及藻体大小等方面,主要不同点:A.setaceum初期藻体的顶端小枝细胞可形成毛,而多姿对丝藻则无此特征;A.pygmaeum的篦状羽枝和小枝细胞较长且顶端尖细,而多姿对丝藻小枝细胞短粗且顶端较钝。而Wollaston[49]认为,大多数藻类有无毛与其发育时期或环境条件有关,并不可作为明确的分类特征。且Norris等[50]指出,自然生长的A.pygmaeum经培养可具有像多姿对丝藻较钝的顶端。因此,Wollaston[49]将A.pygmaeum和A.setaceum作为多姿对丝藻的同物异名处理。A.sparsum由Tokida[51]根据库页岛南端阿尼瓦湾的样品进行首次报道,与多姿对丝藻在四分孢子体中的主轴细胞长宽比、腺体细胞的位置和数量是不同的。但Grant[52]提出,由于A.setaceum和多姿对丝藻之间没有显著的形态差异,因此作同种处理。Athanasiadis[53]在对其样本进行检查后,将A.sparsum和多姿对丝藻归为同物异名种。
绢丝藻属Callithamnion由Feldmann-Mazoyer[54]于1940年建立,分布于太平洋、大西洋和地中海区域。我国仅报道C.corymbosum1种[10,11,55]。绢丝藻和丽丝藻属Aglaothamnion的形态结构相似,但其细胞为单核。
4 结论
本研究利用生物冷冻切片技术,通过形态学和解剖学方法,鉴定出我国渤海地区5种仙菜科物种,分别为仙菜属3种,即波登仙菜CeramiumboydeniiGepp、三叉仙菜CeramiumkondoiYendo和孙氏仙菜CeramiumsungminbooiJ.R.Hughey et G.H.Boo;绢丝藻属1种,即绢丝藻Callithamnioncorymbosum(Smith)Lyngbye;对丝藻属1种,即多姿对丝藻AntithamniondefectumKylin。其中,孙氏仙菜为我国新纪录种,多姿对丝藻为渤海沿岸首次纪录。本研究结果丰富了我国渤海地区仙菜科的物种多样性,可为后续研究提供参考。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29 10:10:20
纺织高校基础科学学报(2022年3期)2022-11-12 03:11:36
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15 06:35:20
东方少年·阅读与作文(2021年5期)2021-06-15 03:06:18
汽车工艺师(2021年4期)2021-05-15 12:57:12
矿产勘查(2020年5期)2020-12-25 02:38:56
上海故事(2020年10期)2020-12-14 04:03:16
科技创新与应用(2017年20期)2017-07-15 20:43:02
湖泊科学(2015年4期)2015-02-17 06:43:49
应用海洋学学报(2014年2期)2014-11-26 01:20:48
