
华南沿海贝克喜盐草种子库与有性繁殖的时空格局及其关系
2025-02-16 00:00:00盘远方苏治南邱思婷邱广龙
广西植物 2025年1期

摘 要: "贝克喜盐草(Halophila beccarii)是全球具有灭绝风险的海草种类之一,被世界自然保护联盟(IUCN) 列为易危(VL)种。为深入了解华南沿海贝克喜盐草海草床土壤种子库与有性繁殖时空分布特征,以期为贝克喜盐草这一濒危物种的保护、管理及恢复提供科学依据,该研究以华南沿海贝克喜盐草为研究对象,研究各采样点雄花、雌花和土壤种子库密度的时空变化特征及其相互关系。结果表明:(1)雄花密度和雌花密度主要表现为义丰溪和诏安较高,其他地方较低;雄花密度和雌花密度随季节变化较大,总体表现为春季或秋季较高。(2)土壤种子库密度表现为花场湾最高,其次是洋浦,其他地方较低;各采样点土壤种子库密度随季节变化具有较高异质性。(3)线性回归结果表明,贝克喜盐草土壤种子库密度与雄花密度呈显著负相关(Plt;0.05),与雌花密度相关性不显著(Pgt;0.05)。(4)结构方程模型结果表明,季节变化是影响贝克喜盐草土壤种子库密度最关键的因素且季节变化对贝克喜盐草土壤种子库密度影响的直接效应(0.16)大于间接效应(0.008)。研究结果对深入理解贝克喜盐草种群的建立、更新和繁殖机制具有重要意义。
关键词: 海草, 贝克喜盐草, 土壤种子库密度, 雄花密度, 雌花密度, 结构方程模型
中图分类号: "Q948
文献标识码: "A
文章编号: "1000-3142(2025)01-0069-11
基金项目: "国家自然科学基金(32170399); 自治区直属公益性科研院所基本科研业务费项目(2022GMRC-01,2022GMRC-02); 广西科学院改革发展专项项目(2024YGFZ504-102)。
第一作者: 盘远方(1994—),硕士,助理研究员,主要从事海草与红树林生态学研究,(E-mail)yuanfangpan124@163.com。
*通信作者: "邱广龙,博士,研究员,主要从事海草与红树林生态学研究,(E-mail)gqiu@gxmarinesciences.ac.cn。
Temporal and spatial pattern and their relationship between
the seed bank and sexual reproduction of Halophila
beccarii along the coasts of South China
PAN Yuanfang1,2, SU Zhinan1,2, QIU Siting1,2, QIU Guanglong1,2*
( 1. "Guangxi Key Lab of Mangrove Conservation and Utilization, Guangxi Academy of Marine Sciences (Guangxi Mangrove
Research Center), Beihai 536000, Guangxi, China; 2. Observation and Research Station of Coastal Wetland
Ecosystem in Beibu Gulf, Ministry of Natural Resources, Beihai 536015, Guangxi, China )
Abstract: "Halophila beccarii is one of the sea grass species with a global risk of extinction and is listed as a vulnerable (VL) species by the International Union for Conservation of Nature (IUCN). To provide scientific evidence for the conservation, management, and restoration of the threatening seagrass H. beccarii, the temporal and spatial distribution pattern and their relationship between the soil seed bank and the sexual reproduction (male and female flowers density) of H. beccarii along the coasts of South China were studied. The results were as follows: (1) There were significant differences in "densities of the male flower and the female flower between the sampling sites, with notably higher densities at Yifengxi and Zhao’an than other sites. And these monitored parameters exhibited marked seasonal fluctuations, with a peak in spring or autumn. (2) The highest soil seed bank density was observed at Huachangwan, followed by Yangpu, while relatively low densities were investigated at other sites. Moreover, soil seed bank densities exhibited considerable heterogeneity and seasonal changes across all sites. (3) Linear regression analysis results revealed a significant negative correlation between soil seed bank density and male flower density(Plt;0.05), while the correlation with female flower density was not significant(Pgt;0.05). (4) Seasonal change was the primary factor influencing soil seed bank density in H. beccarii based on the results of the structural equation modeling (SEM). The direct effect of seasonal changes on soil seed bank density (0.16) was greater than the indirect effects (0.008). This research is of great significance for a deeper understanding of the establishment, renewal, "and reproductive mechanisms of the H. beccarii population.
Key words: seagrass, Halophila beccarii, soil seed bank density, male flower density, female flower density, structural equation modeling (SEM)
土壤种子库是植物种群更新、定居、存活、繁衍和扩散的重要基础,它的存在为植物在将来的演替或遭受干扰和破坏后的再恢复提供繁殖体(Kebrom amp; Tesfaye, 2000; Schwienbacher et al., 2010; 何芳兰等,2018)。土壤种子库中的种子作为基因传播的载体,能促进种群间基因的传递与交流,为整个种群提供优良性状(于婷等,2023)。同时,土壤种子库能够通过“储藏效应”延长种群的存活时间,在亲本植物死亡之后保障了物种、种群和基因型的延续,并且随着时间的推移在空间上分布了遗传多样性(Long et al., 2015; Widdig et al., 2020)。种子库对维持植物群落多样性,降低植物种群灭绝的风险和确保物种延续有着重要意义(Pereira-Diniz amp; Ranal, 2006; Erfanzadeh et al., 2013)。
贝克喜盐草(Halophila beccarii)是所有海草植物中形态最小的种类,也是所有海草植物中最古老的两个谱系之一,具有极高的保护价值(Short et al., 2010)。但受全球气候变化和人为干扰等因素影响,贝克喜盐草一直处于加速衰退的状态,已被世界自然保护联盟(International Union for Conservation of Nature, IUCN)列为受威胁物种红色名录中的易危物种,是当前全球面临灭绝风险的10种海草之一(Short et al., 2011)。与其他种类的海草相比,贝克喜盐草属于小型海草,由于生长速度快、种群周转速率快、种群变化大、形态矮小易受沉积物覆盖等原因,贝克喜盐草极其重要的生态功能并未引起重视(邱广龙等,2016)。贝克喜盐草与所有海草一样,既可以进行无性繁殖也可进行有性繁殖对种群进行更新和延续,不同生活型所采取的繁殖策略存在较大差异(Zakaria et al., 2002)。其中,一年生生活型贝克喜盐草种群有性繁殖期主要集中在某几个月内完成,随后种群进入衰退阶段,次年待环境条件适宜时储存于土壤中的种子开始萌发、长出新植株,随后进行种群建立和扩张(Zakaria et al., 1999)。而多年生生活型种群其有性繁殖期比较分散,一年中均能看到开花结果且种群的更新和延续主要依赖于无性繁殖即根状茎的水平生长(Zakaria et al., 1999)。因此,研究贝克喜盐草有性繁殖对深入理解其繁殖和更新机制具有重要意义。
受生活型、环境变化、气候变化等因素影响,贝克喜盐草土壤种子库和繁殖特征在时间和空间尺度上具有较高的异质性(Hosogi amp; Kameyama, 2004)。有研究认为,种子库的空间异质性主要由种子的输入(包括种子的散布和扩散)和输出(种子萌发、腐烂及捕食等)决定,而雌花和雄花数量对将来种子的输入起决定性作用,是影响贝克喜盐草土壤种子库特征和分布的重要因素(罗超,2021)。另外,种子在成熟后的一段时间内种子库的规模达到峰值,由于季节变化而导致不同环境因素的变化塑造了一年中种子库的动态(Onaindia amp; Amezaga, 2008)。目前,关于贝克喜盐草的研究主要集中在种群分布特征(邱广龙等,2016;郭晋宇等,2023)、种群动态(邱广龙等,2013)、形态特征(邱广龙等,2020)、物候(Zakaria et al., 2002)等基础研究,而涉及贝克喜盐草种子库和有性繁殖的研究资料非常匮乏(邱广龙等,2013;韦梅球等,2017),严重影响了人们对这一濒危物种繁殖策略的认识,也严重阻碍了对其保护和研究工作的开展。因此,研究贝克喜盐草种群种子库和有性繁殖是揭示贝克喜盐草种群更新和延续的重要基础,有助于更好把握贝克喜盐草种群的生长发育状况、种群动态和预测未来的演替方向,对贝克喜盐草种群的更新、恢复具有重要意义(杨磊等,2010)。鉴于此,本研究以华南沿海贝克喜盐草为研究对象,通过测定不同采样点贝克喜盐草雄花、雌花和土壤种子库密度,拟探讨以下问题:(1)分析华南沿海贝克喜盐草雄花、雌花和土壤种子库密度的分布特征;(2)土壤种子库密度与雄花、雌花密度存在何种关系以及季节变化对雄花、雌花和土壤种子库密度有何影响。研究结果对揭示贝克喜盐草种群建立、发展、自我恢复以及对其濒危机制的探讨和资源保育研究具有重要意义。
1 材料与方法
1.1 研究区域概况
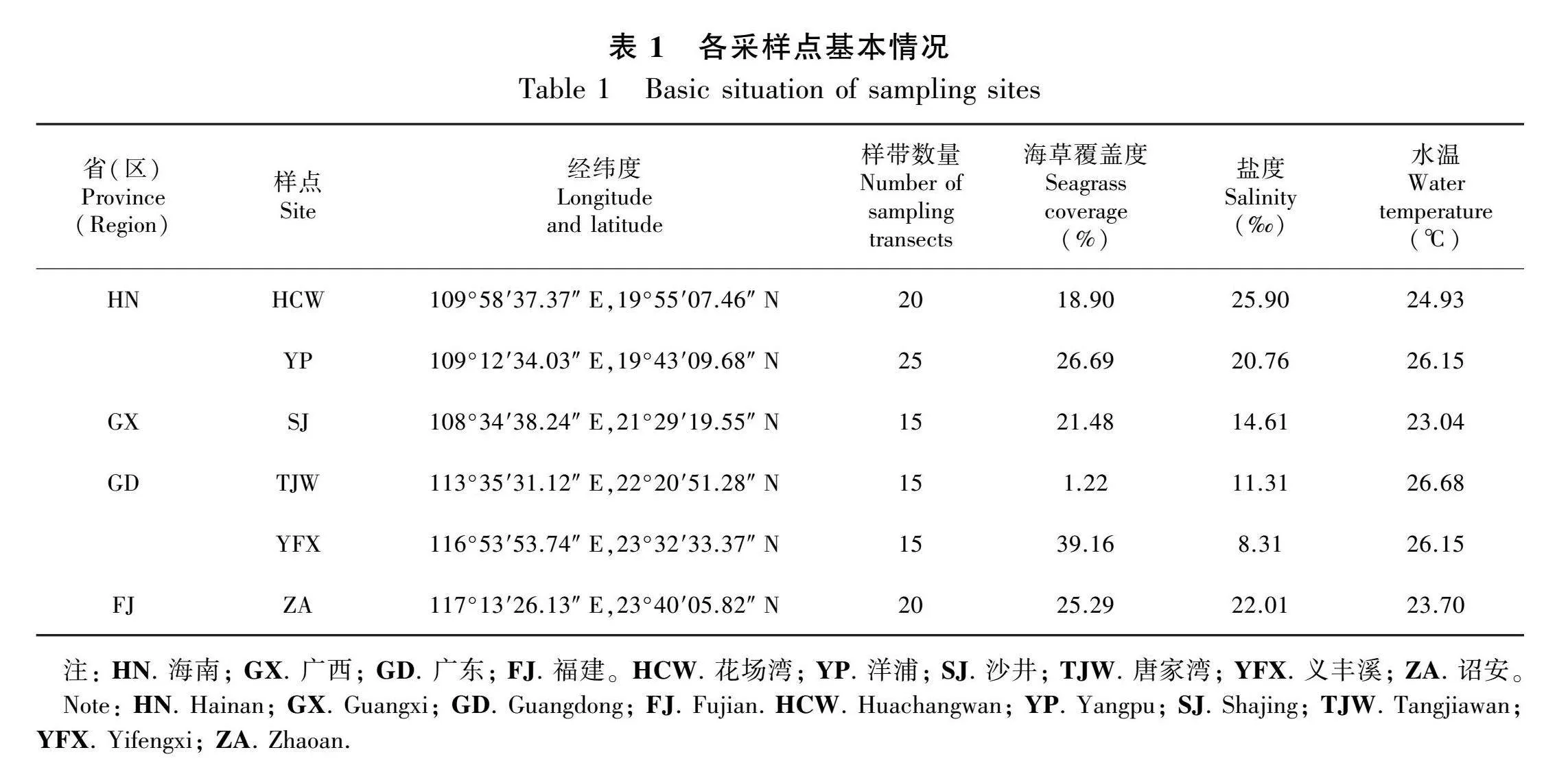
通过收集文献资料并结合实地踏查了解贝克喜盐草在华南沿海的分布情况(邱广龙等, 2013,2016;陈石泉等,2013;Jiang et al., 2020;郭晋宇等,2023),于2022年11月,我们将研究样地设定在海南省儋州市的洋浦(YP)和澄迈县的花场湾(HCW)、广西钦州市的沙井(SJ)、广东省珠海市的唐家湾(TJW)和汕头市的义丰溪(YFX)以及福建省漳州市的诏安(ZA),各采样点分布如图1所示。其中,沙井、唐家湾、义丰溪和诏安属于亚热带季风型海洋性气候,年均降水量为1 300~2 200 mm,降水主要集中在6─8月份,年平均气温18~23 ℃。海南的洋浦和花场湾属于热带季风型海洋性气候,年均降水量在1 600 mm以上,年平均气温23~26 ℃(Geng et al., 2022)。
1.2 样地设置和种群调查
根据贝克喜盐草海草床面积大小、分布、水文情况及与海堤的位置关系,设置垂直于海堤或岸边用于调查贝克喜盐草种群的样带(50 m),每条样带平行间隔50~100 m,共计110条样带。调查时,在每条样带的每间隔8 m处,即2、10、18、26、34、42 m处放置50 cm × 50 cm的样方框平行于样带用于拍照和估算贝克喜盐草覆盖度,同时记录大型藻类的种类及贝克喜盐草叶片附生植物的覆盖率。各采样点样带数量及基本概况如表1所示。
于2022年11月—2023年11月对各采样点进行样带调查。调查时间为2022年11月以及2023年2月、5月、8月和11月,每个季度对各采样点进行一次调查,共进行5次。
1.3 样品的采样与测定
为调查贝克喜盐草雄花密度、雌花密度和土壤种子库密度,在每条样带上用直径为7 cm的圆柱形采样器在样带上随机采集6个(10 cm)带土壤的海草样品(采样点尽可能均匀分布在整条样带上) (邱广龙等,2013),放入有编号的密封袋中,置于便携式冷藏箱内储存,随后带回实验室进行土壤种子库密度和有性繁殖特征的测定。
样品用50目标准筛筛洗后,将海草植物和土壤分开,海草植物用于统计贝克喜盐草雄花、雌花数量,土壤用于统计贝克喜盐草种子的数量(邱广龙等,2013;韦梅球等,2017)。根据样品统计到的雄花、雌花以及土壤中种子的数量计算海草床单位面积内贝克喜盐草雄花密度、雌花密度和土壤种子库密度。
1.4 统计分析
1.4.1华南沿海贝克喜盐草有性繁殖和土壤种子库密度的比较 采用单因素方差分析(one-way ANOVA)对不同采样点贝克喜盐草雄花密度、雌花密度和土壤种子库密度进行差异性统计分析。当贝克喜盐草雄花密度、雌花密度和土壤种子库密度在不同采样点上呈现统计检验显著情况下,即Plt;0.05时,再运用Tukey-Kramer HSD test进行多重比较。
1.4.2 线性回归模型 采用线性回归模型探讨贝克喜盐草土壤种子库密度与有性繁殖(雄花密度和雌花密度)相关性,即以贝克喜盐草的土壤种子库密度作为因变量,以雄花密度、雌花密度以及季节为自变量进行回归分析。
1.4.3 结构方程模型 本研究采用结构方程模型(SEM)进一步探讨季节如何影响贝克喜盐草有性繁殖及其与土壤种子库密度之间的相互关系及路径。结构方程模型是基于变量协方差矩阵分析多变量数据之间关系的综合性数据统计与分析方法,该方法是根据研究者的先验知识预先设定系统内因子间的依赖关系,不仅能够判别各因子之间的关系强度,而且能对整体模型进行拟合和判断(Spitale et al., 2009)。结构方程模型拟合精度与随机效应有密切相关,本研究中采样点(样地效应)和采样时间(时间效应)是影响模型预测结果准确性的主要组成部分,所以本研究所用的结构方程模型嵌套了样地效应和时间效应来探讨影响贝克喜盐草土壤种子库密度的因素。模型公式由下列方程表示:
X=ΛXξ+δ(1)
Y=ΛYη+ε(2)
η=Bη+Γξ+ζ(3)
式中: X、Y分别表示表示外生测量变量向量和内生测量变量向量;"η表示是内生潜变量向量;"ΛX表示X在ξ上的因子载荷矩阵;"ΛY表示Y在η上的因子载荷矩阵;"ξ表示外生潜变量;"B表示内生潜变量之间的作用路径系数矩阵;"Г表示外生潜变量对内生潜变量的影响路径系数矩阵;"δ表示外生指标的误差项;"ε表示内生指标的误差项;"ζ表示结构方程的残差项。
统计分析与绘图均在R 4.3.1软件的piecewiseSEM和ggplot2等程序包中完成。
2 结果与分析
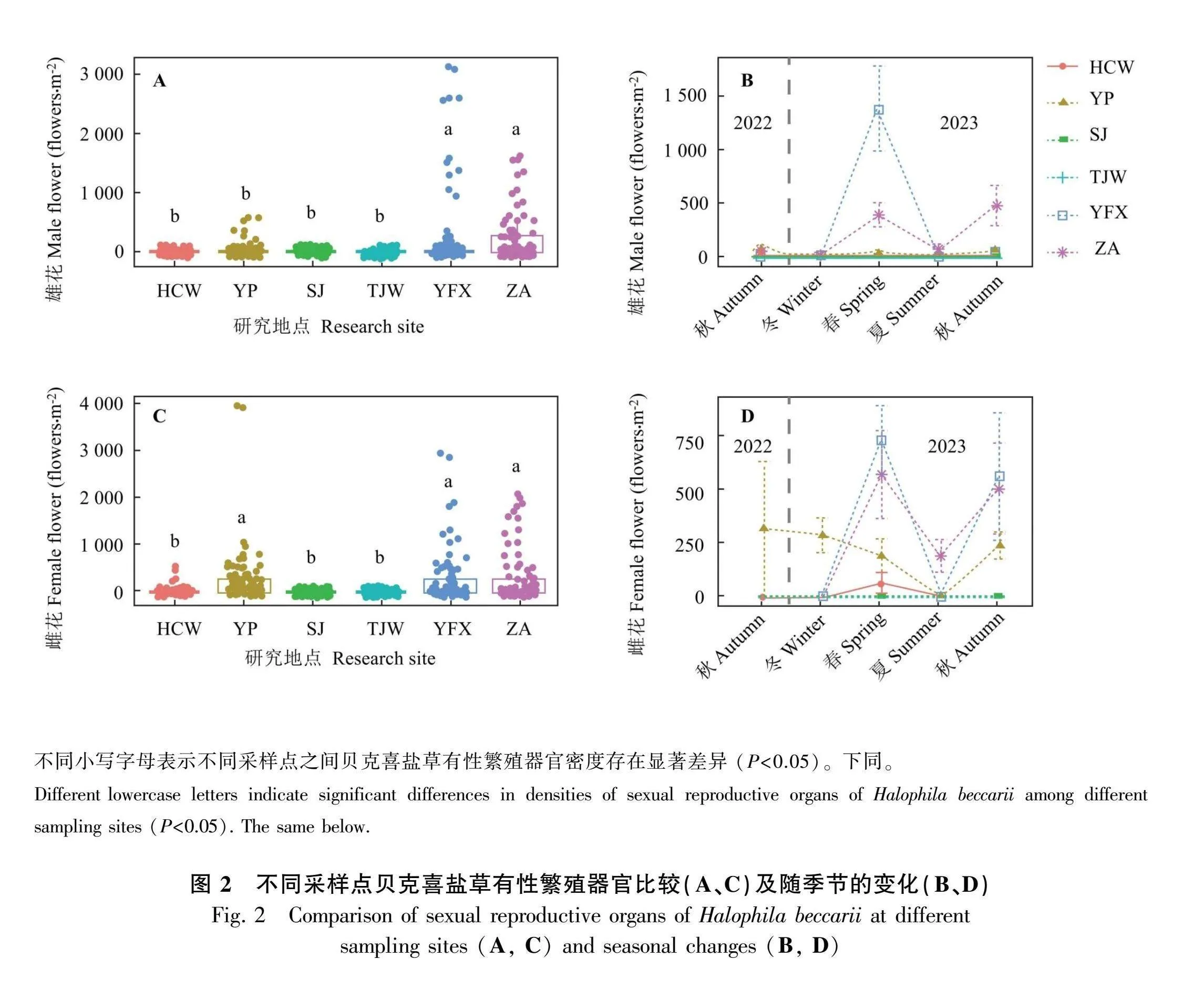
2.1 各采样点贝克喜盐草繁殖器官比较
不同采样点贝克喜盐草雄花密度和雌花密度存在较大差异。其中,雄花密度和雌花密度主要表现为义丰溪和诏安显著高于花场湾、洋浦、沙井和唐家湾(Plt;0.05) (图2:A、C)。从贝克喜盐草雄花密度来看,义丰溪于2023年春季最多,诏安于2023年春季和秋季最多,而其他采样点在调查过程中发现雄花密度非常少甚至为0 (图2:B);而从贝克喜盐草雌花密度来看,花场湾、义丰溪和诏安2023年春季和秋季普遍较高,洋浦除了2023年夏季外其他季节雌花密度均较高,而沙井和唐家湾在整个调查过程中均未发现雌花(图2:D)。尽管不同采样点贝克喜盐草的雄花密度和雌花密度差异较大,但是总体均表现为2023年春季和秋季较高。
2.2 各采样点贝克喜盐草土壤种子库密度比较
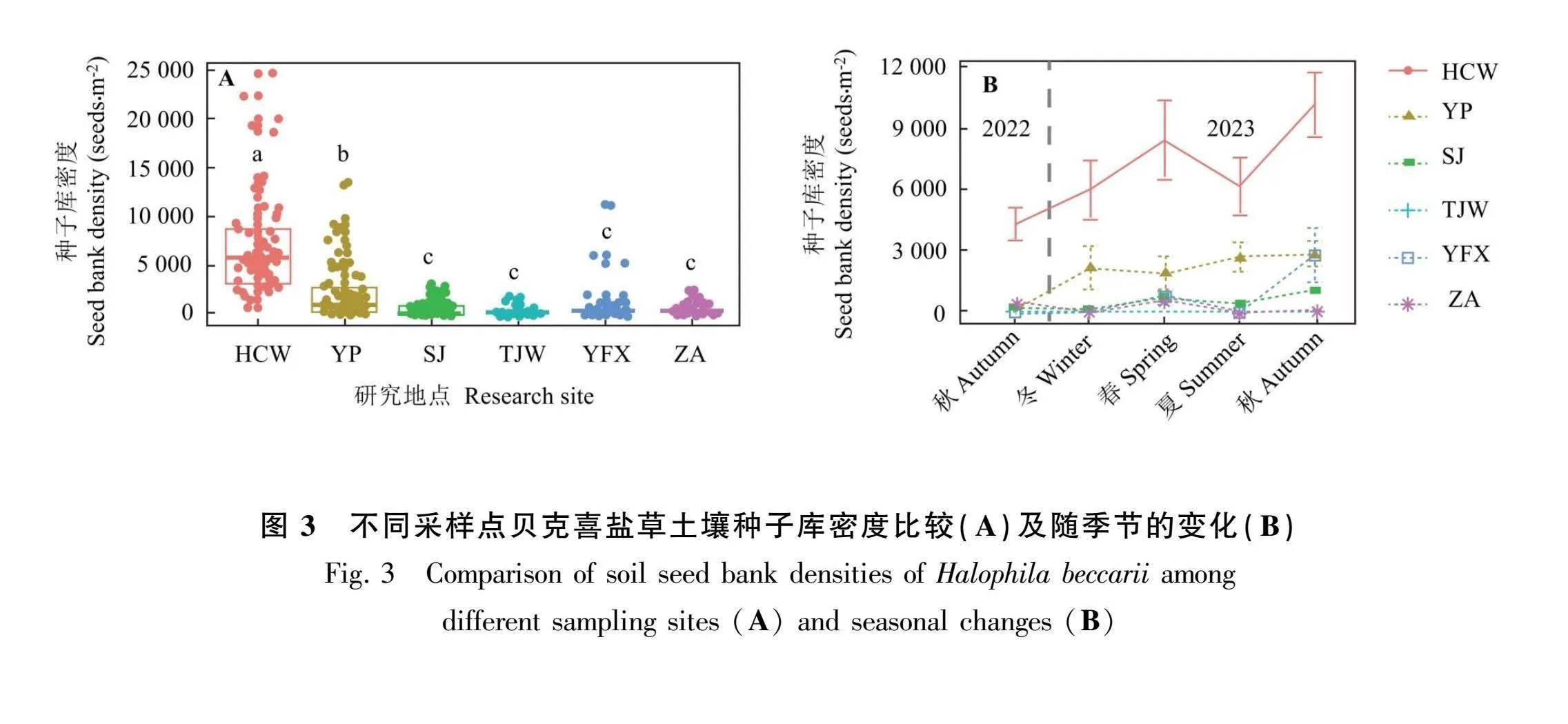
土壤种子库密度表现为花场湾最高,其次是洋浦,而沙井、唐家湾、义丰溪和诏安土壤种子库密度较低(图3:A)。不同季节各采样点贝克喜盐草土壤种子库密度具有较高的异质性,其中花场湾变化最明显,其次是洋浦,其余采样点相对较平缓(图3:B)
2.3 贝克喜盐草土壤种子库与繁殖特征的关系
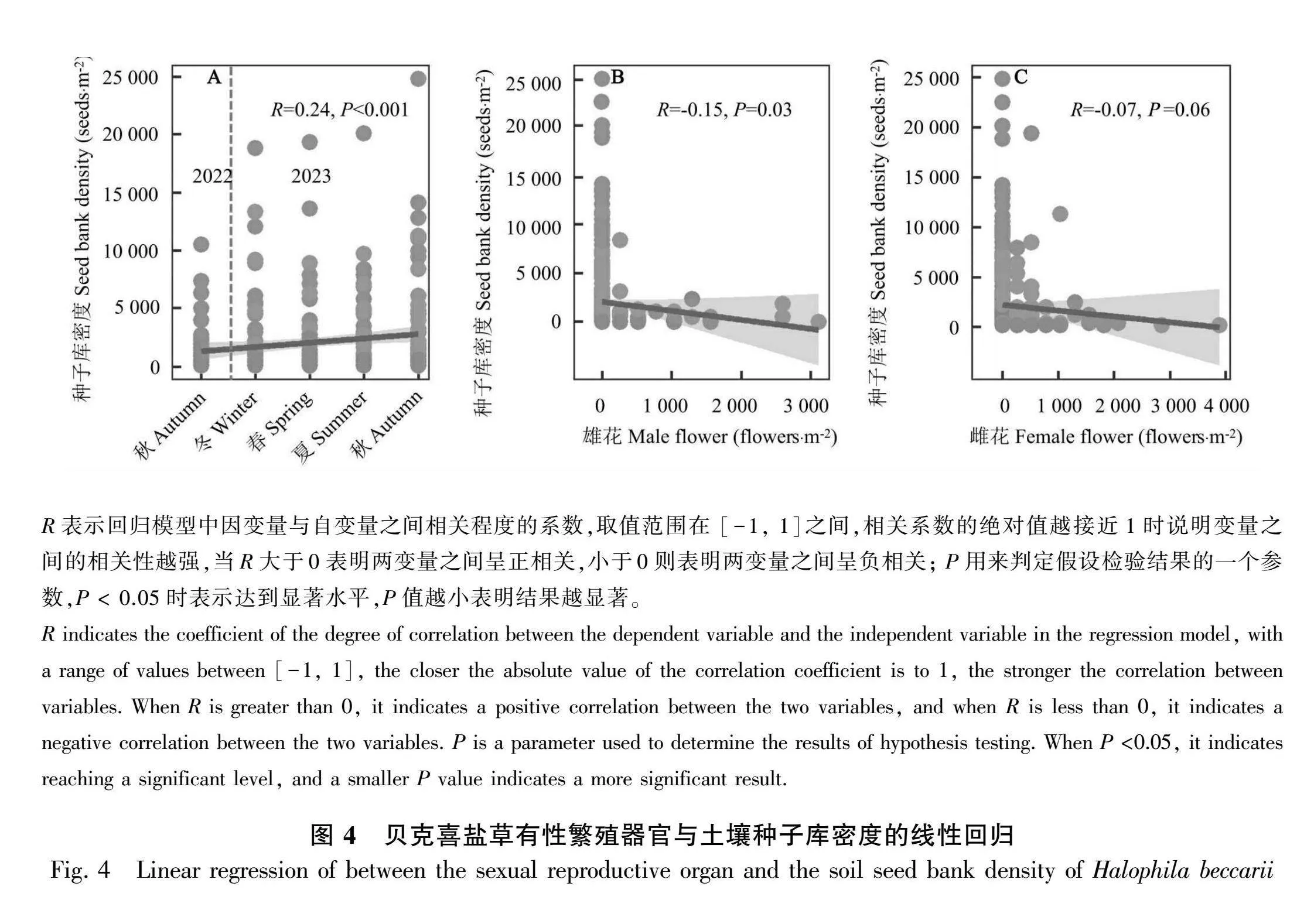
线性回归结果表明,贝克喜盐草土壤种子库密度与调查季节呈显著的正相关(Plt;0.001)、与雄花密度呈显著负相关(Plt;0.05) (图4:A、B)。而贝克喜盐草土壤种子库密度与雌花密度呈负相关,但相关性不显著(图4:C) (Pgt;0.05)。
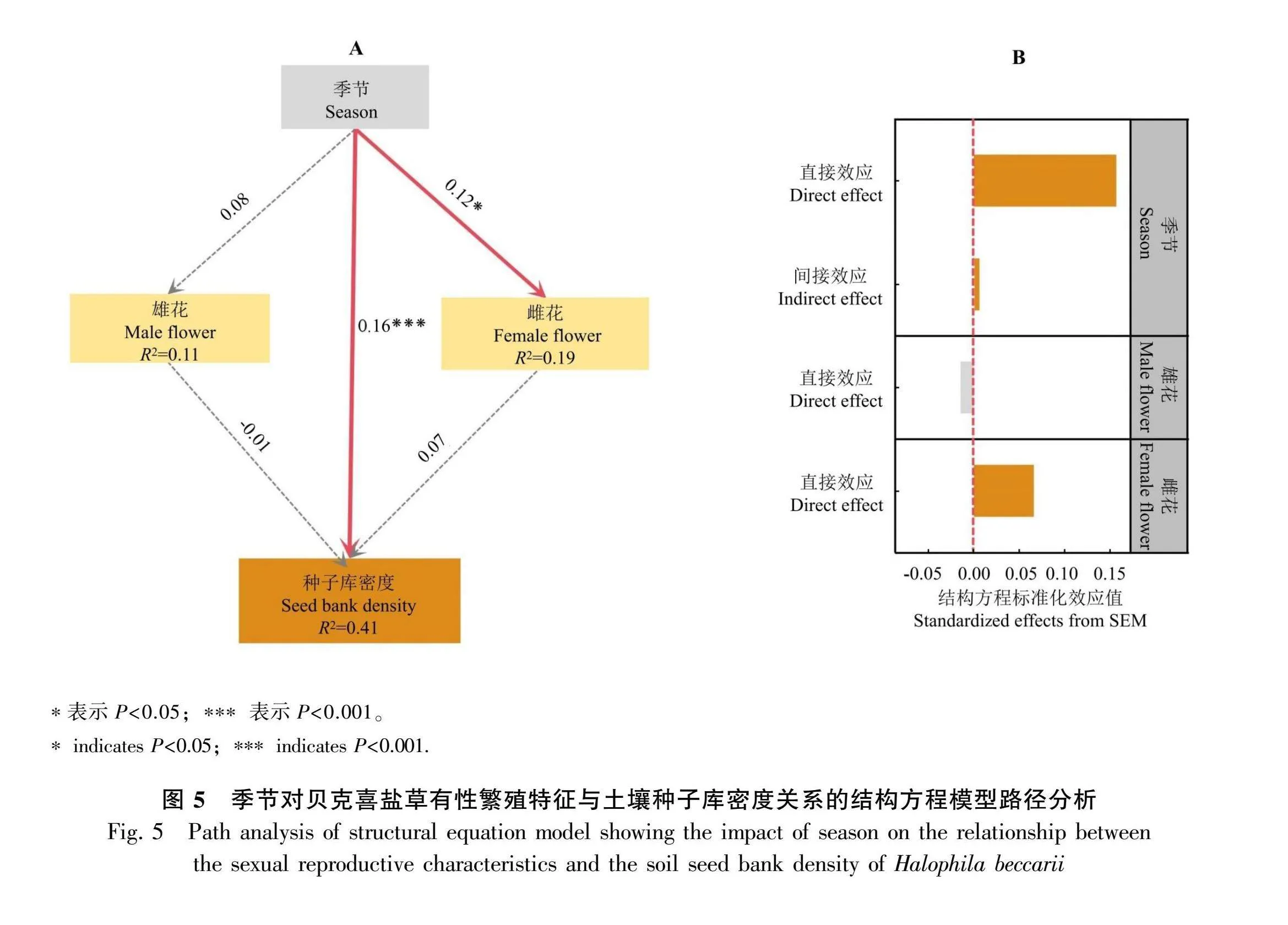
结构方程模型结果表明,季节变化是直接影响贝克喜盐草土壤种子库密度的关键因素,雄花和雌花密度对土壤种子库密度影响不显著(图5:A)。从结构方程标准化结果来看,季节变化对贝克喜盐草土壤种子库密度影响的直接效应值(0.16)大于间接效应值(0.008),雄花密度对贝克喜盐草土壤种子库密度影响的直接效应值为-0.01,雌花密度对贝克喜盐草土壤种子库密度影响的直接效应值为0.07 (图5:B)。
3 讨论
3.1 不同采样点贝克喜盐草雄花、雌花和土壤种子库密度差异分析
植物的繁殖器官数量表征植被繁殖机能和繁衍后代能力大小,通常来说繁殖器官的数量越多,繁殖能力也会相对提高(Smith amp; Johnson, 2022; García amp; López, 2024)。本研究发现,雄花和雌花密度均表现为义丰溪和诏安较高,其他地方较低且雄花和雌花密度随季节变化较大,总体表现为春季和秋季较高,这表明义丰溪和诏安的贝克喜盐草相对于其他采样点的海草具有较高的有性繁殖能力。繁殖器官数量的多少是一个复杂的生物学问题,通常受植物个体的生理状态、环境条件、遗传等多种因素共同影响(Hosogi amp; Kameyama, 2004; Onaindia amp; Amezaga, 2008)。邱广龙等(2013)对南亚热带的广西珍珠湾贝克喜盐草种群研究发现高峰期的雄花密度低于热带地区马来半岛的吉兰丹,而雌花密度则相反。受气候因素制约,贝克喜盐草分全年开花和只在雨季开花两种类型(Zakaria et al., 1999, 2002)。邱广龙等(2013)研究发现,珍珠湾贝克喜盐草种群繁殖期主要集中在雨季,2—3月份气温较低基本看不到繁殖器官且雌花的高峰期于8月初,而雄花高峰期于8月底,并未像热带地区一些种群那样终年开花结果, 与本研究发现(亚热带的义丰溪和诏安贝克喜盐草种群)的雄花和雌花花期主要集中于春季和秋季不一致。这可能与贝克喜盐草所生长的环境有关,我们在调查过程中发现义丰溪和诏安贝克喜盐草种群繁殖器官数量最多的是2023
年5月,其次是2023年11月。从地理位置来看,汕头的义丰溪样地位于义丰溪河口,而诏安样地位于东溪河口,两个采样点之间的直线距离仅有36.03 km且所处的纬度和潮汐相近,这可能造成两个采样点之间具有相似的微气候条件,所以导致两个采样点贝克喜盐草种群的繁殖器官数量出现相似的规律。而其他采样点贝克喜盐草繁殖器官数量在整个调查期间(2022年11月至2023年11月)均较少(除洋浦雌花外),甚至未发现。这可能与贝克喜盐草生活史周期(茂盛或衰退)有很大关系,如唐家湾样地于2023年2月、花场湾和沙井样地于2023年8月之后分别进入衰退期,直至2024年3月前海草床内未发现地上植被存在,所以也未统计到其繁殖器官数量。因此,不同地方贝克喜盐草的繁殖器官数量差异受海草生活史周期影响非常大,同时也受到其生长环境条件以及气候条件等多因素的影响。
土壤种子库是种群更新与拓展的重要基础,也是种群动态的重要组成部分,预示着种群的自我恢复能力,在连接过去、现在和将来的植物种群结构与动态中起着重要的作用(Li amp; Zhao, 2005; Yang et al., 2019 )。本研究发现土壤种子库密度为花场湾最高(每平方米6 991粒),其次是洋浦(每平方米2 023粒),最少的是唐家湾(每平方米96粒)。与广西珍珠湾贝克喜盐草种群相比,花场湾贝克喜盐草土壤种子库密度(每平方米6 991粒)远高于珍珠湾(每平方米2 728粒),洋浦土壤种子库密度(每平方米2 023粒)与珍珠湾接近,而其他采样点土壤种子库密度远低于珍珠湾(邱广龙等,2013)。土壤种子库状况与地上植被有着密切的关系,也可以反映贝克喜盐草生存环境的变化情况。有研究表明,当土壤种子库和地上植被均有时,表明该环境适宜该种群的建成;当土壤种子库有而地上无植被时, 表明该环境不适合该种群的建成;当土壤种子库无而地上有植被时,表明该环境不适合种子的保存;当土壤种子库和地上植被均无时,表明周围环境可能不适宜该种群的建成(Whipple, 1977)。但贝克喜盐草由于具有生长速度快、种群周转速率快等特点(邱广龙等,2020),特别是一年生种群(可在8个月内完成整个生活史过程),一年中不同月份的植物种群外貌有很大的变化(Zakaria et al., 2002)。所以当土壤种子库中有种子而地上无植被时,并不一定代表该环境不适合贝克喜盐草种群的建成,可能是贝克喜盐草处于生活史周期的衰退期。如我们在调查过程中发现花场湾和沙井样地2023年8月份之后进入了衰退期,土壤种子库中有种子但无植被,直至2024年3月份之后沙井样地已经逐渐恢复起来,花场湾于2024年4月份开始恢复,所以针对贝克喜盐草而言,有土壤种子库无地上植被且周围环境无重大变化或受到破坏时,贝克喜盐草可能暂时处于衰退期。另外,不同采样点之间的土壤种子库密度存在较大的差异,这与气候条件以及环境因素存在很大的关系。而本研究所涉及的采样点之间的跨度及距离较大,所以微气候、环境条件(如盐度、温度、潮汐、水文、营养盐以及土壤养分资源)存在较大差异,这可能是导致不同采样点土壤种子库密度差异的重要原因(Zakaria et al., 2002)。
3.2 贝克喜盐草土壤种子库与繁殖器官的关系
土壤种子库中的种子是由雌花的胚珠经过授粉和受精后发育而来,是植物延续生命的载体,同时也是植物向外传播和扩散的载体(Koornneef et al., 2002)。因此,土壤种子库密度与地上繁殖器官数量存在密切的关系。在线性回归中我们发现,贝克喜盐草土壤种子库密度与雄花密度呈显著负相关、与雌花密度呈负相关性但相关性不显著。有研究表明,土壤种子库密度与繁殖器官数量呈负相关关系是由竞争压力所导致,当土壤种子库密度较高时,种子之间为了争夺有限的生长空间和资源,种子之间的竞争变得激烈,这种情况下植物可能会通过减小雄花和雌花密度,以减轻竞争压力,提高植物个体的生存和繁殖成功率(Li et al., 2018; Chen amp; Liu, 2020)。在结构方程模型中我们发现,季节变化是影响贝克喜盐草土壤种子库密度和雌花密度最关键的因素,这与不同生活型贝克喜盐草种群存在较大关系。在调查过程中我们发现唐家湾样地于2023年2月、花场湾和沙井样地于2023年8月后分别进入衰退期,后期调查过程中未观察到贝克喜盐草地上植被存在,同时对样品分析时也未发现有存活的地下根状茎,由此可以确定上述采样点均为一年生生活型海草。洋浦、义丰溪和诏安样地在不同季度调查时其贝克喜盐草种群的面积、覆盖度和密度存在较大差异。例如,洋浦2022年11月直立茎密度最高(36 703.08 ind.·m-2),最低为2023年2月(2 459.86 ind.·m-2);义丰溪2023年11月直立茎密度最高(20 989.69 ind.·m-2),最低为2023年2月(2 367.47 ind.·m-2);诏安2023年8月直立茎密度最高(25 075.02 ind.·m-2),最低为2023年2月(5 066.97 ind.·m-2) (未发表数据)。由于每个季度仅进行一次调查,所以洋浦、义丰溪和诏安样地贝克喜盐草种群暂未确定是否为一年生生活型海草。由于贝克喜盐草具有周转速率快的特点,所以不管是一年生生活型还是多年生生活型,均有较大的季节变化。
此外,对于雌花密度与土壤种子库密度的关系,结构方程模型分析的结果与线性回归模型分析的结果有所差异。这可能与统计学因素有关,在建立线性回归模型时并未考虑不同采样点和采样时间之间的差异对结果产生的影响;采样点(样地效应)和采样时间(时间效应)等随机效应是影响结构方程模型准确度的重要组成部分,因此,结构方程模型在计算过程中将这些随机效应项纳入后模型会根据所纳入的随机效应赋予不同的权重,最后再根据不同的权重进行回归处理,所得出的结果更准确也更有说服力。有研究表明,土壤种子库密度和繁殖器官数量与季节存在密切关系,一般认为植物在雨季会增加植物的繁殖器官数量,以增加植物的繁殖成功率,相应的雨水充足的季节土壤种子库密度也有所增加(Garcia amp; Smith, 2022; Larios amp; Traveset, 2023)。受季节变化和植物生长周期的影响,季节变化对贝克喜盐草土壤种子库密度和繁殖器官数量关系较为复杂,并且还受到外界环境(海水和土壤因素)以及底栖动物取食的共同影响(Chen amp; Liu, 2020)。
4 结论与建议
通过对华南沿海贝克喜盐草土壤种子库和繁殖特征的调查,本研究分析了各采样点贝克喜盐草雄花、雌花和土壤种子库密度差异及其关系。结果表明,研究区所有采样点的贝克喜盐草雄花、雌花和土壤种子库密度具有较大的时空异质性。此外,贝克喜盐草土壤种子库密度与雄花密度主要呈负相关关系。从本研究的野外监测数据来看,对于生长快、周转速率快的贝克喜盐草而言,特别是一年生生活型的种群,每个季度只进行一次监测,其监测频度远远不够。由于本研究采样点比较多、纬度跨度也比较大,完成一次全面的监测需要消耗大量的时间、投入大量的人力物力,因此下一步可考虑仅对个别典型的海草床以及监测指标进行逐月监测,而针对非典型海草床和监测指标可开展季度监测,从而确保在有限资源的基础上得到更科学合理的监测结果。本研究结果对贝克喜盐草濒危机制的探讨和资源保育的研究,种群自我恢复潜在能力的评估,以及其保护对策的制定等都具有现实的理论意义。
参考文献:
CHEN S, LIU J, 2020. Effects of seasonal variation in soil seed bank density on reproductive organ allocation in a desert plant community [J]. Journal of Arid Environments, 181: 104246.
CHEN SQ, WU ZJ, WANG DR, et al., 2013. Research on the interspecific relationships of the seagrass bed communities in Hainan Island [J]. Marine Science Bulletin, 32 (1): 78-84. [陈石泉, 吴钟解, 王道儒, 等, 2013. 海南岛海草床群落种间关系研究 [J]. 海洋通报, 32 (1): 78-84.]
ERFANZADEH R, KAHNUJ SHH, AZARNIVAND H, et al., 2013. Comparison of soil seed banks of habitats distributed along an altitudinal gradient in northern Iran [J]. Flora Morphology Distribution Functional Ecology, 208: 312-320.
GARCIA Y, SMITH B, 2022. Influence of soil seed bank density on reproductive organ production in a temperate grassland ecosystem: A seasonal study [J]. Journal Plant Ecology, 15(3): 321-333.
GARCA M, LPEZ J, 2024. Variation in reproductive organ number and its implications for reproductive output in a tropical plant species [J]. Botanical Studies (1): 78-89.
GENG XX, CAI ZF, JIA SW, et al., 2022. Environmental determinants of the distribution of Halophila beccarii Ascherson in Hainan Island, China [J]. Sustainability, 14(20): 13491.
GUO JY, GENG XX, YIN L, et al., 2023.Distribution characteristics of Halophila beccariir resources in "Danzhou Bay, Hainan and their influencing factors [J]. Transactions of Oceanology and Limnology, 45(2): 124-132. "[郭晋宇, 耿晓晓, 尹雷, 等, 2023. 海南儋州湾贝克喜盐草资源分布特征及影响因素分析 [J]. 海洋湖沼通报, 45(2): 124-132.]
HE FL, GUO CX, MA JM, et al., 2018. Dynamic changes in soil seed banks and their relationships with aboveground vegetation during the decaying of Haloxylon ammodendron plantations at the edge of the Minqin oasis [J]. Acta Ecologica Sinica, 38(13): 4657-4667. "[何芳兰, 郭春秀, 马俊梅, 等, 2018. 民勤绿洲边缘梭梭林衰败过程中土壤种子库动态及其与地上植被的关系 [J]. 生态学报, 38(13): 4657-4667.]
HOSOGI D, KAMEYAMA A, 2004. Timing for the collection of topsoil from a deciduous forest for use as planting material in suburban Tokyo, Japan [J]. Ecological Engineering, 23(4): 371-386.
JIANG ZJ, CUI LJ, LIU SL, et al., 2020. Historical changes in seagrass beds in a rapidly urbanizing area of Guangdong Province: Implications for conservation and management [J]. Global Ecology and Conservation, 22: e01035.
KOORNNEEF M, BENTSINK L, HILHORST H, 2002. Seed dormancy and germination [J].Current Opinion in Plant Biology, 8(1): 33-36.
KEBROM T, TESFAYE B, 2000. The role of soil seed banks in the rehabilitation of degraded hillslopes in Southern Wello, Ethiopia [J]. Biotropica, 32: 23-32.
LI QY, ZHAO WZ, 2005. Advances in the soil seed bank of arid regions [J]. Advance in Earth Sciences, 20(3): 350-358.
LI Y, CHEN Y, JIANG, Y, et al., 2018. Influence of soil seed bank density on the regeneration of dominant tree species in a subtropical forest [J]. Journal Plant Ecology, 11(3): 419-428.
LARIOS E, TRAVESET A, 2023. Seasonal variation in soil seed bank density and its relationship with reproductive output in a Mediterranean shrubland [J]. Plant Ecology, 224(1): 67-78.
LUO C, 2021. Study on the characteristics of soil seed bank and its influencing factors and activation effect in arid desert mining area [D]. Beijing: Beijing Forestry University. "[罗超, 2021. 干旱荒漠矿区土壤种子库特征的影响因素和激活效应研究 [D]. 北京: 北京林业大学.]
LONG RL, GORECKI MJ, RENTON M, et al., 2015. The ecophysiology of seed persistence: A mechanistic view of the journey to germination or demise [J]. Biological Reviews, 90(1): 31-59.
ONAINDIA M, AMEZAGA I, 2008. Seasonal variation in the seed banks of naive woodland and coniferous plantations in Northern Spain [J]. Forest Ecology amp; Management, 126: 163-172.
PEREIRA-DINIZ S, RANAL MA, 2006. Germinable soil seed bank of a gallery forest in Brazilian Cerrado [J]. Plant Ecology, 183: 337-348.
QIU GL, SU ZN, FAN HQ, et al., 2020. Biological and ecological characteristics of intertidal seagrass Halophila beccarii and its conservation countermeasures [J]. Marine Environmental Science, 39(1): 122-126. "[邱广龙, 苏治南, 范航清, 等, 2020. 贝克喜盐草的生物学和生态学特征及其保护对策 [J]. 海洋环境科学, 39(1): 122-126.]
QIU GL, FAN HQ, LI ZS, et al., 2013. Population dynamics and seed banks of the threatened seagrass Halophila beccarii in Pearl Bay, Guangxi [J]. Acta Ecologica Sinica, 33(19):6163-6172. "[邱广龙, 范航清, 李宗善, 等, 2013. 濒危海草贝克喜盐草的种群动态及土壤种子库——以广西珍珠湾为例 [J]. 生态学报, 33(19): 6163-6172.]
QIU GL, SU ZN, ZHONG CR, et al., 2016. Distribution and community characteristics of threatened seagrass Halophila beccarii in Dongzhai Harbor, Hainan [J]. Guihaia, 36(7): 882-889. "[邱广龙, 苏治南, 钟才荣, 等, 2016. 濒危海草贝克喜盐草在海南东寨港的分布及其群落基本特征 [J]. 广西植物, 36(7): 882-889.]
SPITALE D, PETRAGLIA A, TOMASELLI M, 2009. Structural equation modelling detects unexpected differences between bryophyte and vascular plant richness along multiple environmental gradients [J]. Journal of Biogeography, 36: 745-755.
SHORT FT, COLES R, WAYCOTT M, et al., 2010. Halophila beccarii//IUCN 2010. The IUCN red list of threatened species [DB]. Version 2014.3.http://www.iucnredlist.org. Downloaded on 16 December 2014.
SHORT FT, POLIDORO B, LIVINGSTONE SR, et al., 2011. Extinction risk assessment of the world’s seagrass species [J]. Biological Conservation, 144(7): 1961-1971.
SMITH A, JOHNSON B, 2022. The relationship between reproductive organ number and reproductive success in flowering plants [J]. Journal Plant Reproduction (3): 45-56.
SCHWIENBACHER E, MARCANTE S, ERSCHBAMER B, 2010. Alpine species seed longevity in the soil in relation to seed size and shape-A 5-year burial experiment in the Central Alps [J]. Flora, 205(13): 19-25.
WHIPPLE SA, 1977. The relationship of buried, germinating seeds to vegetation in an old-growth Colorado subalpine forest [J]. Canadian Journal of Botany, 56(13): 15061509.
WIDDIG M, HEINTZ-BUSCHART A, SCHLEUSS PM, et al., 2020. Effects of nitrogen and phosphorus addition on microbial community composition and element cycling in a grassland soil [J]. Soil Biology amp; Biochemistry, 151: 1-37.
WEI MQ, ZHAO LJ, QIU GL, et al., 2017. Spatial distribution of the soil seed banks of seagrass Halophila beccarii at intertidal zone and the biological influential factors [J]. Guangxi Sciences, 33(2): 93-101. "[韦梅球, 招礼军, 邱广龙, 等, 2017. 潮间带贝克喜盐草土壤种子库空间分布及其生物影响因素 [J]. 广西科学, "33(2): 93-101.]
YANG L, WANG YR, YU JD, 2010. Review of research on seed banks of desert regions [J]. Acta Prataculturae Sinica, 19(2): 227-234. "[杨磊, 王彦荣, 余进德, 2010. 干旱荒漠区土壤种子库研究进展 [J]. 草业学报, 19(2): 227-234.]
YANG X, ZHANG J, LIU Q, et al., 2019. Seasonal dynamics of soil seed bank density and its impact on plant community composition in a subtropical forest [J]. Forest Ecology and Management, 448: 1-9.
YU T, HU XJ, XING HW, 2023. Spatial distribution characteristics of soil seed bank of "Malus sieversii population in Emin County [J]. Hubei Agricultural Sciences, 62(10): 119-123. "[于婷, 胡晓静, 邢皓文, 2023. 额敏县新疆野苹果种群土壤种子库空间分布特征 [J]. 湖北农业科学, 62(10): 119-123.]
ZAKARIA MH, SIDIK BJ, HISHAMUDDIN O, 1999. Flowering, fruiting and seedling of Halophila beccarii Aschers. (Hydrocharitaceae) from Malaysia [J]. Aquatic Botany, 65(1/2/3/4): 199-207.
ZAKARIA MH, BUJANG JS, ARSHAD A, 2002. Flowering, fruiting and seedling of annual Halophila beccarii Aschers. in Peninsular Malaysia [J]. Bulletin of Marine Science, 71(3): 1199-1205.
(责任编辑 周翠鸣)
猜你喜欢
知识窗(2022年10期)2022-07-06 05:23:24
学苑创造·B版(2022年10期)2022-05-30 12:25:35
军事文摘·科学少年(2022年11期)2022-05-30 10:48:04
老年博览·上半月(2022年10期)2022-05-30 10:48:04
智慧少年·故事叮当(2019年12期)2019-02-18 06:27:42
动漫界·幼教365(小班)(2018年5期)2018-05-14 11:52:08
现代商贸工业(2016年24期)2017-01-13 18:24:17
高教探索(2016年12期)2017-01-09 21:55:40
财经问题研究(2016年7期)2016-12-26 14:38:25
价值工程(2016年31期)2016-12-03 23:01:46
