
微管壁上二聚体间相互作用能与微管长度关系探究
2025-02-03 00:00:00胡健
西北师范大学学报(自然科学版) 2025年1期
关键词:模型



摘要:基于微管中空和规则的周期性结构,将其璧上的原丝纤维视为一维铁电系统.每个结构单元αβ二聚体可视为“双势阱”模型,可移动电子可能处在“α势阱”中,也可能处在“β势阱”中,分别对应方向向上和向下两种赝自旋量子态,采用“赝自旋”量子理论探讨了微管壁上一根原丝纤维上二聚体之间相互作用能与微管长度的关系.
关键词:“双势阱”模型;赝自旋理论;二聚体间相互作用能;微管长度
中图分类号:Q 616""" 文献标志码:A""" 文章编号:1001-988Ⅹ(2025)01-0041-04
DOI:10.16783/j.cnki.nwnuz.2025.01.008
收稿日期:20231220;修改稿收到日期:20240610
基金项目:上海市教师专业发展工程资助项目(10110N240032-A07)
作者简介:胡健(1979—),男,山西保德人,讲师,硕士.主要研究方向为细胞骨架微管动力学行为.
Email:shdxhj2005@163.com
The relationship study between the interaction energy
of dimers on the microtubule wall and the lengths of microtubule
HU Jian
(School of Sciences,Shanghai Institute of Technology,Shanghai 201418,China)
Abstract:Considering the hollow and periodic structure of microtubule(MT),the protofilaments of the MT are regarded as a one-dimensional ferroelectric system.Each αβ dimer unit can be treated as a“double potential well”model,and the mobile electron may be in“α potential well”also in“β potential well”,corresponding to two pseudo-spin quantum states in the direction of up and down respectively.The relationship between the dimers interaction energy of a protofilament on the MT wall and MT lengths was investigated by the“pseudo-spin”quantum theory.
Key words:“double potential well”model;pseudo-spin theory;interaction energy of dimers;lengths of microtubule
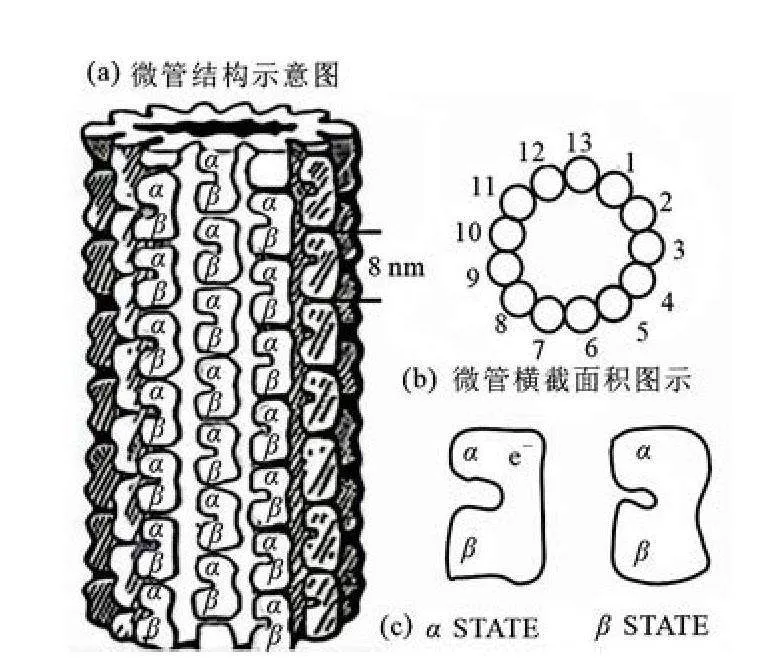
微管普遍存在于真核细胞的细胞质中,是细胞骨架的重要组成部分和功能组件,在神经细胞内含量丰富,是细胞组织和信息处理的中心[1].从X射线衍射实验得知[2],微管外径约为25 nm,内径约为14 nm,它是由13根微管蛋白原丝纤维螺旋盘绕而成的中空桶状管,中空部分充满液体水.微管是细胞质骨架中直径最大的纤维,其长度范围从几十纳米到几百微米不等.
电子晶体衍射实验进一步表明[3],组成微管壁的原丝纤维是由α及β微管蛋白单体以αβ二聚体形式(长度约为8 nm)存
在且以首尾顺次相连的方式聚合而成,其结构如图1所示.每个α或β微管蛋白单体的长度约为4 nm,每一个二聚体中均有一个可移动电子,当电子处于α微管蛋白中时称为α构象,处于β微管蛋白中时称为β构象[4].由于微管壁上原丝纤维上的每一个二聚体中均有一个可移动电子,故每个二聚体均可看作是一个电偶极子.因此Mavromato等[5]认为,微管是一种含水的铁电体,微管蛋白分子的有序排列是微管蛋白二聚体电偶极子和微管管腔中有序水分子相互作用的结果.
Fig 1The structure of microtubule
由于微管蛋白溶液中的微管是一种极性结构,故微管始终处于聚合和解聚不断调整的动态不稳定状态之中,因此微管壁原丝纤维上的二聚体始终处于振动状态[6-8],这对细胞内能量输送和信息传递发挥着极其重要的作用;同时二聚体间的相互作用会激发其内部电子的隧道效应[9],伴随着电子的隧穿,二聚体将发生α构象与β构象之间的相互转换,在转变过程中二聚体的形状将发生很大的变化,这对细胞的生命活动非常重要,而原丝纤维上二聚体间的相互作用能量在上述生命活动中发挥着极其重要的作用.文中以“赝自旋”理论为基础,定量计算了微管壁上一根原丝纤维上二聚体之间的相互作用能量与微管长度之间的关系.
1" “赝自旋”理论
微管中空和规则的周期格子结构显示出用量子物理的微观尺度来研究它的必要性[10-11].微管壁上原丝纤维上的αβ二聚体的结构可看作是一个“双势阱”模型,
由于每一个二聚体中均有一个可移动电子,故每个二聚体可视为一个赝自旋态.电子可能处在“α势阱”中,也可能处在“β势阱”中,分别对应方向向上和向下两种赝自旋量子态.对于一维赝自旋系统[12],两个二聚体间的相互作用项可以表示为
=12∫∫Ψ+(x)Ψ+(x′)e2εx-x′×
Ψ(x)Ψ(x′)dxdx′,(1)
其中,Ψ+(x)和Ψ(x)为满足费米对易关系的粒子场算符;
Jij=-4v+-+-(i,j),(2)
vλυγδ(i,j)≈∫∫ω*λ(x-i)ωυ(x-i)e2εx-x′×
ω*γ(x′-j)ωδ(x′-j)dxdx′,(3)
其中,vλυγδ(i,j)为二聚体i和j之间的耦合项,与两二聚体之间的距离有关,λ,ν,γ和δ分别表示两个二聚体可能的4种量子自旋态;ωλ(x-i)和ω*λ(x-i)分别代表瓦尼尔函数及其变形式.
一根原丝纤维上二聚体间总的相互作用能量可表示为
J0=∑jJij.(4)
其中Jij表示两个二聚体i和j之间的耦合能量.
2" 结果与讨论
微管壁上的原丝纤维是由α及β微管蛋白单体(每个α或β微管蛋白单体的长度约为4 nm)以αβ二聚体形式存在且以首尾顺次相连的方式聚合而形成.二聚体的结构可看作是一个“双势阱”,为了获得具体有效的计算结果,利用两个谐振子势来代替上述双势阱,采用量子微扰论方法经计算得到相邻两个二聚体之间的耦合强度J12≈0.1987/ε[ev],对于微管蛋白二聚体,其介电常数ε通常取10.
对于某一二聚体而言,其所在的原丝纤维上其它二聚体对它总的相互作用能量与该二聚体所处的位置和微管长度均有关,主要存在两种可能的情况,一种是二聚体i处在原丝纤维的中间,如图2所示.
Fig 2The schematic diagram of the dimer in the
middle of the protofilament
原丝纤维上其他二聚体对其总相互作用能可以表示为
J0=∑jJij=
-4e2ε1ri,i+1+1ri,i+2+…+1ri,i+N++
1ri-1,1+1ri-2,i+…+1ri-N-,i=
e2ε1ri,i+1+1ri,i+1+ri+1,i+2+…+
1ri,i+1+ri+1,i+2+…+ri+N+-1,i+N++
1ri-1,i+1ri-2,i-1+ri-1,i+…+
1ri-N-,i-N-+1+ri-N-+1,i-N-+2+…+ri-1,i=
-e2ε1ri,i+11+12+13+…+1N++
1ri-1,i1+12+13+…+1N-=
J121+12+13+…+1N++
1+12+13+…+1N-.
另一种情况是二聚体恰好处在原丝纤维的一端,如图3所示,该情况下原丝纤维上其他二聚体对它其总相互作用能可以表示为
J0=∑jJij=-4e2ε
1ri,i+1+1ri,i+2+…
+1ri,i+N+=
e2ε1ri,i+1+1ri,i+1+ri+1,i+2+…+
1ri,i+1+ri+1,i+2+…+ri+N+-1,i+N+
-e2ε1ri,i+11+12+13+…+1N+=
J1,21+12+13+…+1N+.
以情况二为例,N++1代表一根原丝纤维上二聚体(长度约8 nm)的个数,经计算可得不同微管长度对应的J0(表1),其变化规律如图4所示.
Fig 3The schematic diagram of the dimer at one end
of the protofilament
从图4可以看出,对于有限长度的微管,其原丝纤维上二聚体间的相互作用能随微管长度的增加呈现出自然对数级递增关系,符合微管在细胞中的生命活动规律,这种相互作用能量是微管正常生命活动所需能量的重要来源之一.
3" 结论
微管的中空和规则的周期格子结构以及微管内壁的有序水层提示了几个量子场方面的信息,文中将微管璧上的原丝纤维视为一维动力学系统,在“双势阱”模型的基础上采用“赝自旋”量子理论研究了微管壁上一根原丝纤维上二聚体之间相互作用能与微管长度的关系.通过计算发现,原丝纤维上二聚体间的相互作用能随着微管长度的增加按照自然对数级关系递增.由于微管蛋白溶液中的微管是一种极性结构,在一定条件下,微管一端发生聚合反应使微管延长,原丝纤维上二聚体间的相互作用能不断累积,达到一定长度后另一端发生解聚反应使微管缩短,称为“踏车”现象,微管始终处于聚合和解聚不断调整的动态不稳定状态之中[13].因此微管壁原丝纤维上的二聚体始终处于振动状态,这对细胞内能量输送和信息传递发挥着极其重要的作用;同时二聚体间的相互作用会激发其发生α构象与β构象之间的相互转变,这在细胞的生命活动中非常重要,而原丝纤维上二聚体微管蛋白间的相互作用能可能是上述生命活动的主要能量来源之一,因此研究二聚体之间的相互作用能对弄清楚微管振动的能量来源有非常重要的参考价值.
参考文献:
[1]" HAMEROFF S R,WATT R C.Information processing in microtubules[J].J Theor Biol,1982,98:549.
[2]" AMOS L,KLUG A.Arrangement of subunits in flagellar microtubules[J].J Cell Sci,1974,14: 523.
[3]" NOGALES E,WOLF S G,DOWNING K H.Structure of the"" tubulin dimer by electron crystallography[J].Nature,1998,391:199.
[4]" POKOMY J,JELINEK F,TRKAL V.Electric field around microtubules[J].Bioelectrochem Bioenerg,1998,45:239.
[5]" MAVROMATOS N E,MERSHIN A,NANOPOULOS D V.QED-cavity model of microtubules implies dissipationless energy transfer and biological quantum teleportation[J].Int J Mod Phys B,2002,16:3623.
[6]" JOBS E,WOLF D E,FLYVBJERG H.Modeling microtubule oscillations[J].Phys Rev Len,1997,79:519.
[7]" POKOMY J,JELINEK F,TRKAL V,et al.Vibrations in microtubules[J].J Biol Phys,1997,23:171.
[8]" PORTER G S,TUSZYNSKI J A,HOGUE C,et al.Elastic vibrations in seamless microtubules[J].Eur Biophys J,2005,34:912.
[9]" 胡健,邱锡钧.微管壁上电子量子行为的研究[J].西北师范大学学报(自然科学版),2010,46:32.
[10]" SHIRMOVSKY S E,SHULGA D V.Quantum relaxation effects in microtubules[J].Phys A,2021,582:126254.
[11]" SHIRMOVSKY S E,SHULGA D V.Quantum relaxation processes in microtubule tryptophan system[J].Phys A,2023,617:128217.
[12]" CHEN Y,QIU X J,LI R X.Pseudo-spin model for the cytoskeletal microtubule surface[J].Chin Phys Lett,2004,21:2313.
[13]" LUCHNIAK A,KUO Y W,MCGUINNESS C,et al.Dynamic microtubules slow down during their shrinkage phase[J].Biophys J,2023,122:616.
(责任编辑" 孙对兄)
猜你喜欢
童话王国·奇妙逻辑推理(2024年5期)2024-06-19 16:03:38
网络安全与数据管理(2022年1期)2022-08-29 03:15:20
导航定位学报(2022年4期)2022-08-15 08:27:00
中学生数理化·中考版(2022年8期)2022-06-14 06:55:24
新世纪智能(数学备考)(2021年9期)2021-11-24 01:14:36
成都医学院学报(2021年2期)2021-07-19 08:35:14
新世纪智能(数学备考)(2020年9期)2021-01-04 00:25:14
中学生数理化·七年级数学人教版(2020年10期)2020-11-26 08:24:50
数学物理学报(2020年2期)2020-06-02 11:29:24
光学精密工程(2016年6期)2016-11-07 09:07:19
