
苦马豆(Sphaerophysa salsula)6个不同居群的染色体核型分析
2024-12-31 00:00:00曲荣举刘玉萍陈金元苏旭张朋辉靳佳瑞余明君杨倩
草地学报 2024年8期



摘要:本研究采用染色体常规压片技术和核型分析方法,对苦马豆(Sphaerophysa salsula)6个自然居群(P81,P89,P96,P102,P137,P142)的染色体核型特征进行了分析,探讨了不同居群间的进化趋势和亲缘关系。结果表明:苦马豆6个参试居群的染色体数目和基数恒定,均为2n=2X=16;主要有中部着丝粒(m)和亚中部着丝粒(sm)染色体两种类型,中部着丝粒染色体数量多于亚中部着丝粒染色体;绝大多数居群的核型公式为2n=2X=16 m,仅居群P137的核型公式为2n=2X=14 m+2 sm;染色体长度介于1.05~2.63之间,属于小型染色体;核型类型包括1A,2A,3A和2B,平均臂比处于1.18~1.43之间,长度比居于1.44~2.05之间,核不对称系数介于54.02~58.98之间;居群P89的核型类型为1A,核不对称系数最小,进化程度最低;当遗传距离为4时,苦马豆6个居群形成三大类,居群P137和P142的亲缘关系较近。本研究首次分析了苦马豆的染色体核型特征,探讨了不同居群间的进化趋势和亲缘关系,为苦马豆属物种系统发育和基因组学研究提供了细胞学的佐证。
关键词:苦马豆;染色体;核型分析;进化趋势;亲缘关系;聚类分析
中图分类号:Q941+.2""" 文献标识码:A"""" 文章编号:1007-0435(2024)08-2469-09
Chromosome Karyotype Analysis from Six Different Populations
of Sphaerophysa salsula (Fabaceae)
QU Rong-ju1,2, LIU Yu-ping1,2,3, CHEN Jin-yuan1,2,3, SU Xu1,2,3*,
ZHANG Peng-hui1,2, JIN Jia-rui1,2, YU Ming-jun1,2, YANG Qian1,2
(1.School of Life Sciences, Qinghai Normal University, Xining, Qinghai Province 810008, China; 2. Key Laboratory of Biodiversity
Formation Mechanism and Comprehensive Utilization of the Qinghai-Xizang Plateau in Qinghai Province, Qinghai Normal
University, Xining, Qinghai Province 810008, China; 3. Academy of Plateau Science and Sustainability, Qinghai Normal
University, Xining, Qinghai Province 810016, China)
Abstract:We analyzed the karyotype features of six natural geographic populations (P81,P89,P96,P102,P137,P142) of Sphaerophysa salsula and discussed their evolutionary trends and relationships by the conventional squash technique of chromosomes and karyotype analysis. The results showed that:(1) The number and basic number of chromosomes was constant across six populations of S. salsula,with 2n=2X=16. There were mainly two types of mid-centromeric and sub-centromeric chromosomes,and the number of mid-centromeric chromosomes was more than that of sub-centromeric chromosomes. (2) The karyotype formula of most populations of S. salsula was 2n=2X=16 m,except that the karyotype formula of P137 was 2n=2X=14 m+2 sm. The chromosome length was between 1.05 and 2.63,which belonged to small chromosomes. (3) Karyotype types of S. salsula included 1A,2A,3A and 2B,the mean arm ratio was from 1.18 to 1.43,the length ratio was between 1.44 and 2.05,and the karyotype asymmetry coefficient was 54.02~58.98. Besides,the karyotype type of P89 was 1A,with the lowest karyotype asymmetry coefficient and the lowest evolutionary degree. (4) When the genetic distance was four,the six populations of S. salsula were classified into three groups. Among them,P137 and P142 had the closest relationship. This study firstly discussed the chromosome karyotype features of S. salsula,and studied the evolutionary trends and relationships of different populations,which could provide the cytological evidence for phylogeny and genomic studies of the species of Sphaerophysa.
Key words:Sphaerophysa salsula;Chromosome;Karyotype analysis;Evolutionary trend;Relationship;Clustering analysis
苦马豆(Sphaerophysa salsula)是豆科(Fabaceae)、蝶形花亚科(Papilionoideae)、苦马豆属(Sphaerophysa)的一种多年生草本植物,是我国西北荒漠区的重要旱生植物,主要分布于海拔960~3180 m的草原、荒漠、沙滩、山坡、荒地、沟渠、盐池。苦马豆具有较强的耐盐碱、耐贫瘠等特性,生态价值较高,是盐碱地改良的一种重要资源植物[1];同时,它具有较高的药用价值,利尿、消炎、止血,可用于治疗肝硬化、浮肿、肾炎等疾病[2]。目前,国内外对苦马豆的研究主要集中于种子萌发特性[3-5]、化学成分[6-8]、药理作用[9-11]、共生微生物[12-14]等方面,而关于苦马豆系统发育的研究相对较少[15],尤其从群体水平探讨苦马豆染色体数目和核型分析的研究未见报道。
染色体是遗传物质的主要载体,在细胞分裂过程中由染色质凝缩而成,其形态和数目是最稳定的细胞学特征之一[16-17]。染色体核型分析是基础的细胞遗传学研究方法,主要通过观察和统计细胞有丝分裂中期的染色体,计算其数目和长度等数量特征,分析着丝点位置、染色体类型和随体有无等结构特征[18],进而探究同种和不同物种之间的亲缘关系[19]。迄今为止,国内外诸多学者[20-22]利用染色体压片技术研究了许多被子植物的染色体数目及核型特征,并阐明了这些植物的系统位置和进化关系,取得了显著成效。譬如,Rane等[20]分析了虎掌藤属(Ipomoea)10种植物的染色体核型,认为该属植物均为二倍体,2n=2X=30,并推断了物种间的进化关系;顾志建等[21]报道了青藏高原地区云雾雀儿豆(Chesneya nubigena)的染色体数目和核型,发现其为二倍体,核型公式为2n=2X=16=10 m+6 sm,属于2B型;白俐等[22]采用常规压片法研究了苦参属(Sophora)6种植物的染色体数目和核型,结果表明该属植物的染色体基数均为9,属内不同物种间染色体核型差异较大;同样,杨萍等[23]采用常规染色体压片技术研究了扇穗茅6个不同自然地理居群的染色体数目和核型,发现参试居群间的亲缘关系与海拔高度存在极大的相关性。此外,研究还表明染色体制片和核型分析可以为植物高质量基因组研究提供细胞学的基础数据[24]。据此,本研究采用常规染色体压片技术和核型分析方法,探究苦马豆6个不同居群的染色体数目和核型,推断其进化趋势和亲缘关系,旨在为苦马豆属物种系统发育和基因组学研究提供细胞学方面的理论佐证。
1 材料与方法
1.1 试验材料
本研究的试验材料为2021年7—9月采自内蒙古、陕西、宁夏和青海地区苦马豆6个自然地理居群的成熟种子,具体采样信息详见表1。凭证标本均保存于中国科学院西北高原生物研究所青藏高原生物标本馆(QTPMB)。
1.2 试验方法
1.2.1 种子萌发 从苦马豆6个参试居群中精选完整饱满且无虫孔的成熟种子,然后从每个居群中分别选取10粒种子进行萌发试验。即先将种子置于—20℃冰箱中处理24 h,随后将其均匀放于预先铺有用蒸馏水湿润的两层滤纸的培养皿中[25],并于22℃恒温培养箱中萌发;待苦马豆种子根长长至0.5~1.0 cm时进行取样预处理。
1.2.2 预处理 剪取的苦马豆根尖置于预先蒸馏水湿润的2.0 mL EP管中,立即将其透入到充满N2O密封罐中处理130 min,设置压强为0.55 MPa。
1.2.3 固定和解离 预处理好的苦马豆根尖立即放入含有卡诺氏固定液(95%乙醇∶冰醋酸=3∶1,现配现用)的2.0 mL EP管中,保证根尖充分浸入固定液,然后迅速将EP管放置于4℃冰箱中至少固定10 min,随后用45%的乙酸解离5 min,待用。
1.2.4 制片、染色和观察 取出解离完成的苦马豆根尖,放置于载玻片中央,用双面刀片切除除根尖顶端分生组织外的其他部分,然后用改良的石炭酸品红染液对其染色,7 min后盖上盖玻片,立即用镊子钝端轻轻敲击盖玻片,使根尖分生组织充分散开,随后迅速将载玻片掠过酒精灯外焰,蒸干组织区域的液体,用滤纸垂直向下按压盖玻片,使染色体均匀分散;用Olympus BX53型相差显微镜观察染色体形态和统计数目,同时选取形态良好、数目清晰的分裂中期染色体视野拍照[26]。
1.2.5 数据分析 从苦马豆每个参试居群中选择30个以上处于分裂中期的细胞,利用Olympus BX53型相差显微镜确定染色体数目和倍性;然后,从每个居群中分别挑选10个分散良好、数目完整、形态清晰的染色体进行核型分析。即采用Photoshop 2021软件统计苦豆子每个个体的所有染色体长臂长和短臂长[27],测量数据导入Excel中进行统计;采用常规方法计算染色体的相对长度和臂比,依据李懋学[28]方法进行染色体配对;根据Kuo等[29]的方法计算相对长度系数、平均臂比、染色体长度比、核不对称系数等参数,并利用Arano[30]和Stebbins[31]方法进行参数统计和核型分类。此外,根据刑世岩[32]方法分析苦马豆染色体核型进化趋势,使用Excel软件绘制核型二维进化图;参照李晓莉等[33]方法计算染色体平均臂比及其方差、相对长度方差、染色体长度比、各类型染色体比例等参数,并采用SPSS 26软件进行苦马豆染色体的聚类分析。
2 结果与分析
2.1 染色体数目与倍性鉴定
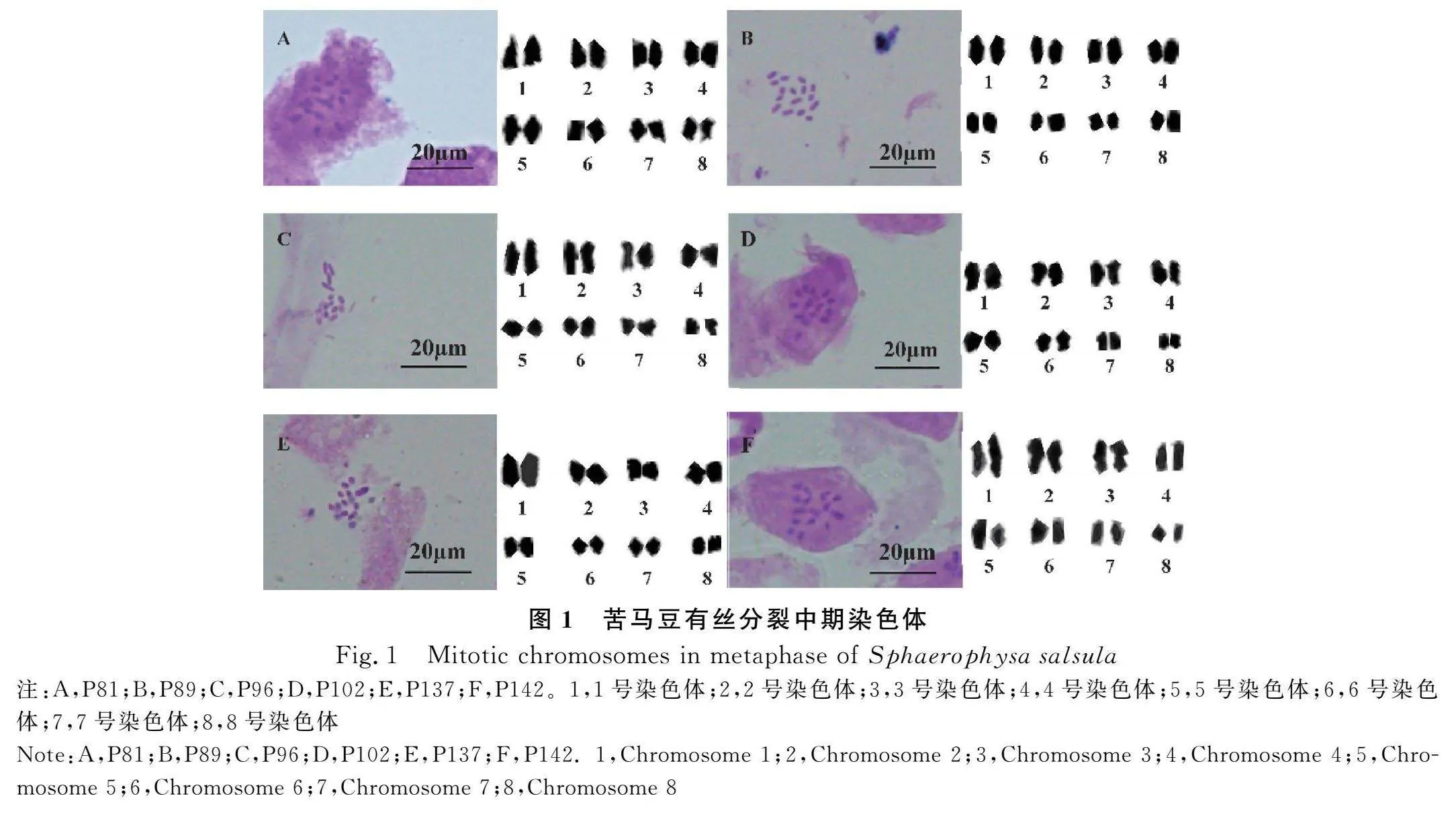
研究结果表明,苦马豆6个居群的染色体均为二倍体,基数为8,且数目恒定,2n=2X=16,不存在染色体数目和形态的异常(图1)。
2.2 染色体形态特征与类型分析
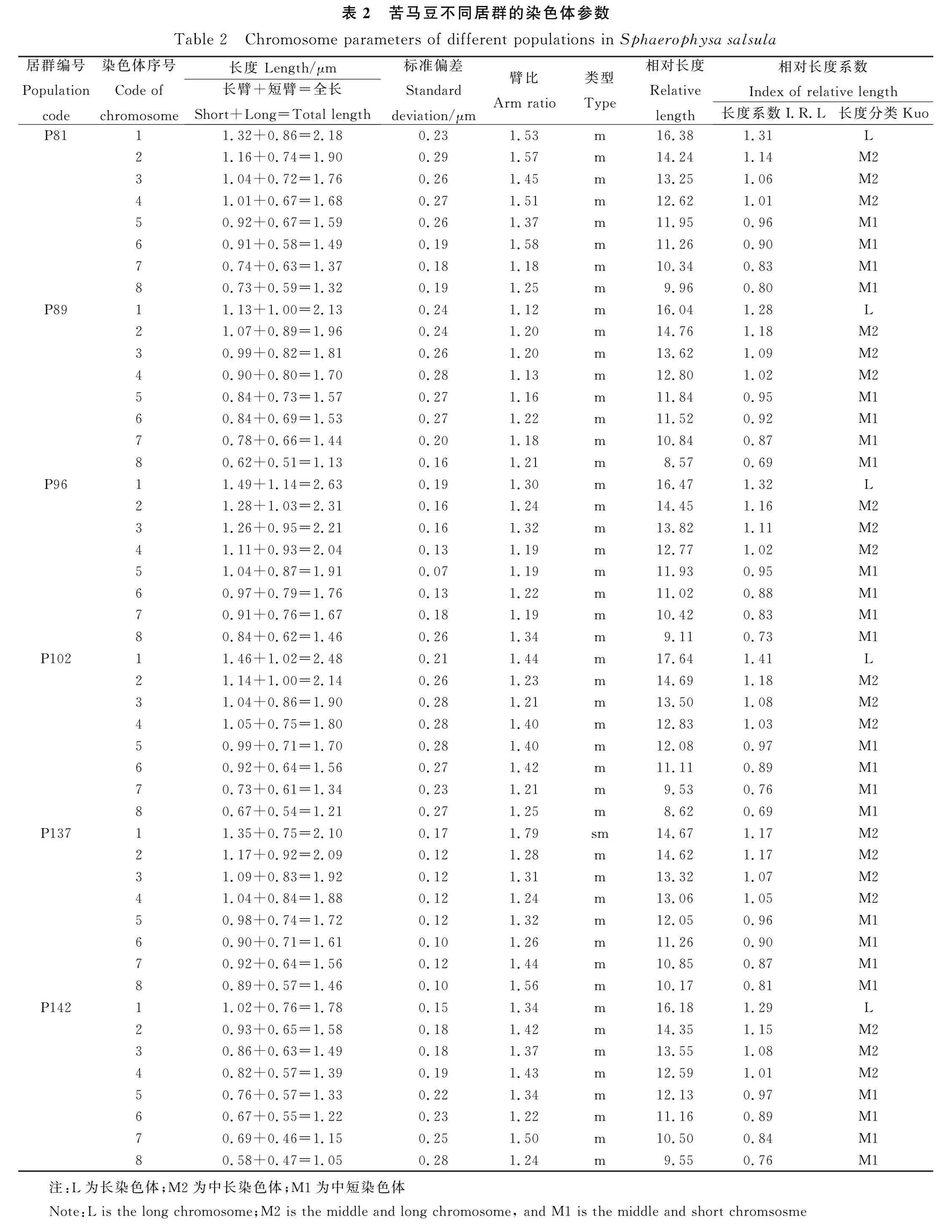
从苦马豆每个参试居群中,分别选取较为清晰的染色体中期分裂图,然后根据着丝点位置以及染色体相对长度、臂比等参数,对其进行同源染色体配对,结果显示每个居群均有8对染色体,参数统计结果详见表2。
从表2可以看出,除居群P137的染色体由中部着丝点染色体(m)和近中部着丝点染色体(sm)组成外,其他所有居群的染色体均为中部着丝点染色体(表2)。同时,根据染色体相对长度系数[34]和Kuo[29]分类标准,苦马豆的染色体可以划分为长染色体L型(≥1.26)、中长染色体M2型(1.01~1.25)和中短染色体M1型(0.76~1.00)三种类型,其中居群P137的染色体类型为M2和M1型,而其他5个居群的染色体类型均为L,M2和M1型(表2)。此外,本研究还根据苦马豆染色体的配对结果和参数数据,绘制了染色体的核型模式图(图2)。
2.3 染色体核型分析
基于苦马豆所有居群染色体长度的统计分析,本研究得到了它们的染色体长度比、臂比大于2的比例、核型公式和类型以及核不对称系数等参数。结果显示,苦马豆所有居群染色体的核型公式均为2n=2X=16,其中居群P137的核型公式为2n=2X=14 m+2 sm,其他参试居群的核型公式为2n=2X=16 m;长度比介于1.44~2.05之间,居群P102的长度比最大(2.05),居群P137的长度比最小(1.44)(表3)。苦马豆染色体核型类型包括1A,2A,2B和3A,其中居群P137和P142中臂比大于2的染色体占比较高,核型类型为3A;而居群P89不存在臂比大于2的染色体,核型类型为1A,说明居群P89的染色体最对称(表3)。此外,苦马豆参试居群染色体平均臂比处于1.18~1.43之间,且平均臂比较高的居群,其核不对称系数也较大;其中,居群P81的核不对称系数相对较大(58.98%),居群P89的核不对称系数较小(54.02%),其他居群的核不对称系数介于两者之间(表3)。
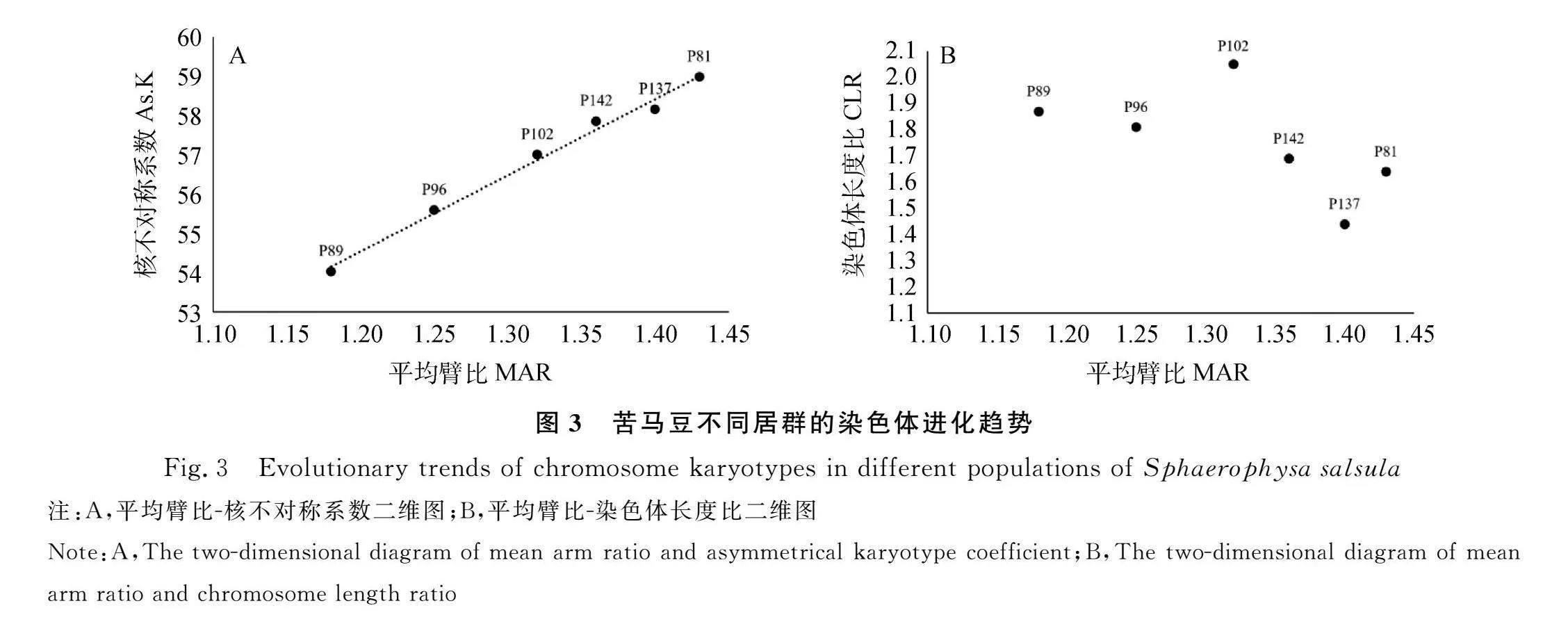
同时,本研究还以平均臂比(MAR)为横轴,以核不对称系数(As.K)和染色体长度比(CLR)为纵轴,分别构建“平均臂比-核不对称系数”和“平均臂比-染色体长度比”的二维图(图3),分析苦马豆参试居群间染色体核型的进化趋势。研究结果表明,苦马豆染色体的平均臂比与核不对称系数呈正相关,6个参试居群沿平均臂比-核不对称系数向右上角进化程度依次增高(图3A)。其中,居群P81的核不对称系数最大,染色体对称性最低;居群P89的核不对称系数最小,染色体对称性最高。此外,从图3B可以明显看出苦马豆的染色体核型呈“双向进化趋势”,其中居群P81和P102的进化程度最高。
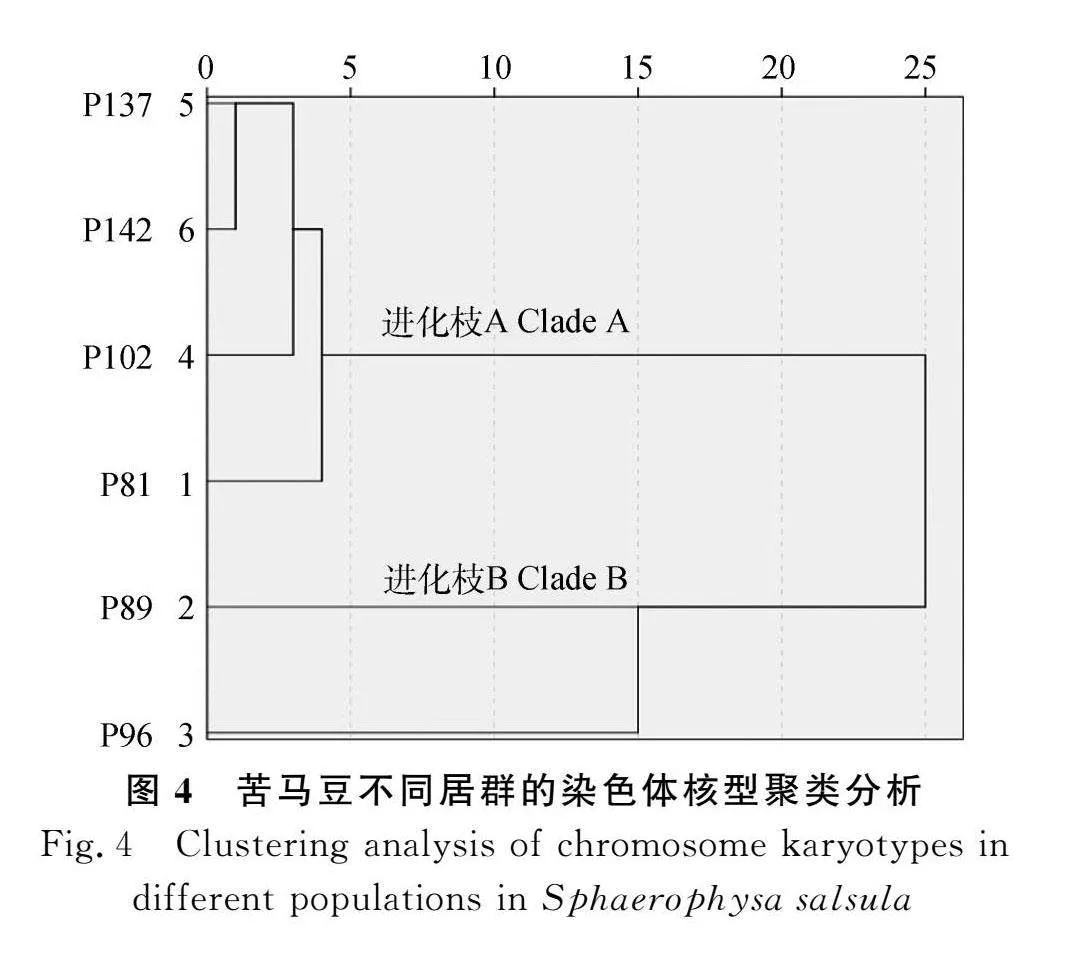
另外,依据参试居群染色体的平均臂比、臂比gt;2的染色体比例、核不对称系数等,本研究还构建了苦马豆参试居群间的核型聚类图,并分析了不同居群间的亲缘关系。结果显示,当遗传距离为25时,参试居群形成A,B两大分支,其中居群P89和居群P96构成B支,居群P137,P142,P102,P81组成A支;遗传距离为15时,参试居群聚为3个分支,其中构成B支的两个居群形成独立的分支;当遗传距离为1时,6个参试居群独立成支(图4)。因此,居群P137和P142以及居群P89和P96的亲缘关系相对较近。
3 讨论
染色体数目、形态、结构和核型作为较稳定的细胞学特征,对研究植物的分类、起源和系统进化关系有着重要作用[35-36];并且,植物染色体数目和基数还可为后续基因组研究提供基础数据[37]。譬如,张力天等[38]通过常规根尖压片法对西藏日喀则市白草染色体数目的统计和核型分析,发现白草的染色体数目为36,基数为9(2n=4X=36=34 m+2 sm)。本研究发现,苦马豆的染色体数目为16条,基数为8(2n=2X=16),这与先前李红等[39]对骆驼刺(Alhagi camelorum)染色体核型特征的研究结果一致,推测它们可能是豆科植物中亲缘关系较近的类群,这也得到苏丹丹等[15]利用叶绿体基因组数据构建的苦马豆和骆驼刺的亲缘关系结果的支持和印证。除居群P137的核型公式为2n=2X=14 m+2 sm外,苦马豆5个居群的核型公式均为2n=2X=16 m,表明苦马豆的染色体具有较高的遗传稳定性[40]。Du等[41]认为额外的DNA主要添加到植物较小染色体的长臂上且受地理位置和海拔高度的影响,因而苦马豆6个参试居群中仅有居群P137的染色体核型由7对中部着丝粒染色体和1对近中部着丝粒染色体构成,这可能是由于该居群主要生长于纬度相对较低的生境中所导致的。据此,本研究认为苦马豆染色体形态变化产生的多样性是其长期适应环境演化的结果[42]。
染色体形态作为重要的核型特征,可被用于探讨不同物种或不同居群间的亲缘关系和进化趋势[43]。譬如,孙立民等[44]通过对古银杏(Ginkgo biloba)半同胞家系染色体核型分析及进化趋势的研究,认为不同的地理区位和漫长的生活史期间的环境变化是导致古银杏各家系间核型组成和类型存在较大差异的主要原因;宛涛等[45]通过观察分析内蒙古不同生态区冷蒿(Artemisia frigida)的染色体核型,发现不同生态区冷蒿的染色体形态和倍性具有一定的差异,推测这可能是由生态环境的影响造成的;王亚男等[46]采用染色体常规压片技术探讨了沙鞭(Psammochloa villosa)不同居群的染色体核型特征和进化关系,认为同种植物染色体形态随着生境不同而发生变化。本研究发现,苦马豆参试居群的染色体长度介于1.05~2.63之间,隶属于小型染色体;染色体核型可被划分为1A,2A,3A和2B,长度比、核不对称系数、平均臂比等均存在明显差异,说明苦马豆不同参试居群间的染色体形态差异较大,这可能由于它们所处的地理环境条件不同造成,进一步验证了先前研究结果[44-46]的正确性与合理性,同样认为同种植物在长期的环境因子作用下将会逐渐演变成适应各自生境的生态居群。
研究表明,植物染色体向着核型不对称的方向进化,即核型越不对称进化程度越高;着丝粒位置和染色体长度差异决定植物染色体的核不对称性[31]。通常,植物染色体具有向核不对称系数增大以及Stebbins标准1A—4C方向演化的趋势[47]。本研究结果表明,苦马豆参试居群的核不对称系数介于54.02%~58.98%之间,居群的进化程度依次为P89lt;P96lt;P102lt;P142lt;P137lt;P81;参试居群的染色体核型类型可被划分为1A,2A,3A和2B,居群染色体的进化程度为P89lt;P81lt;P96lt;P137lt;P142lt;P102。由此看来,居群P89的进化程度最低,即该居群的核型类型为1A,核不对称系数最小,染色体对称性最高,推测宁夏回族自治区石嘴山市可能为苦马豆的生物多样性中心和起源中心[34]。相反,居群P81核不对称系数最大,进化程度最高。研究表明,海拔高度的变化能够引起降水、地表温度和升温速率等环境因子的改变,尤其随着海拔的升高温度会逐渐降低[48]。苦马豆6个参试居群中,居群P81生长的海拔最高,核不对称系数最大,这可能是由于其处于较低的环境温度进而影响染色体着丝粒位置导致的。同样,居群P102的染色体属于2B型染色体,长度比最大,进化程度最高,研究团队推测较高的纬度条件共同促进了居群P102的染色体长度差异变化[41]。因此,本研究认为生态因子对苦马豆染色体的进化方向具有重要影响,不同的生态因子往往使苦马豆的染色体核型向着特定的方向进化,这为以后苦马豆的引种驯化和优良种质资源筛选提供了重要参考。
另外,聚类分析能够有效整合多变量信息来推断不同物种和不同居群间的亲缘关系[49]。本研究结果显示,当遗传距离为25时苦马豆参试居群形成A,B两大分支,其中A支包括居群P137,P142,P102和P81,B支包含居群P89和P96,同一分支内不同居群间具有较近的亲缘关系;当遗传距离为1时,居群P137和P142先聚成1个分支,然后与其他居群形成不同的分支;并且,居群P137和居群P142始终亲缘关系最近。本研究首次报道了苦马豆不同居群的染色体核型特征、进化趋势和亲缘关系,可以为将来解析苦马豆属乃至蝶形花亚科物种系统进化和群体基因组学研究提供细胞学佐证。
4 结论
苦马豆染色体的数目为16条,基数为8,核型公式为2n=2X=16;除居群P137的染色体由近中部着丝粒(sm)和中部着丝粒(m)染色体构成外,其他居群的染色体均为中部着丝粒染色体。苦马豆的染色体长度介于1.05~2.63之间,属于小型染色体;平均臂比处于1.18~1.43之间,长度比居于1.44~2.05之间;染色体核型类型包括1A,2A,3A和2B,核不对称系数介于54.02%~58.98%之间,染色体呈“双向进化趋势”;当遗传距离为4时,苦马豆6个居群形成三大类,居群P137和P142的亲缘关系较近。本研究首次分析了苦马豆不同居群的染色体核型特征与差异,探讨了它们的进化趋势和亲缘关系,可为今后苦马豆属物种系统发育和基因组学研究提供细胞学佐证。
参考文献
[1] 梁湘兰,郭松. 苦马豆叶绿体基因组密码子偏好性分析[J]. 西北林学院学报,2022,37(2):121-126
[2] 中国植物志编辑委员会. 中国植物志:第9(2)卷[M]. 北京:科学出版社,2002:378
[3] 王进,张勇,席亚丽,等. 光照、温度、土壤水分和播种深度对苦马豆种子萌发和幼苗生长的影响[J]. 西北农业学报,2011,20(10):157-161
[4] 黄文娟,张越,梁继业,等. 光-温耦合条件对3种豆科植物种子萌发特性的影响[J]. 江苏农业科学,2014,42(6):234-237
[5] 刘红玲,庄丽,齐晓丽,等. 外界因素对苦马豆种子萌发及幼苗生长的影响[J]. 石河子大学学报(自然科学版),2010,28(6):696-699
[6] 马忠俊,孟大利,王玉霞,等. 苦马豆根和茎中化学成分的研究[J]. 沈阳药科大学学报,2003(2):104-106
[7] 李占林,芦晓燕,孟大利,等. 苦马豆根和茎中化学成分的研究[J]. 中草药,2005,20(1):104-106
[8] 白玛桑姆,杨晓雯,马烽,等. 苦马豆生物碱成分薄层色谱分析[J]. 动物医学进展,2014,35(6):58-61
[9] 杨扶德,罗文蓉,林丽,等. 苦马豆的生药学研究[J]. 中医药学刊,2003(9):1587-1588
[10]周明,陶静仪,许顺尧,等. 苦马豆总黄酮甙的降压作用及其机理[J]. 中国药理学与毒理学杂志,1987(3):258-262
[11]贺莹. 苦马豆杀螨活性成分研究[D]. 杨凌:西北农林科技大学,2014:17-18
[12]周启武,赵宝玉,路浩,等. 苦马豆内生真菌分离鉴定与多样性分析[J]. 畜牧兽医学报,2013,44(3):465-474
[13]邓振山. 苦马豆根瘤中内生细菌遗传多样性分析[J]. 草业科学,2016,33(10):1951-1962
[14]白玛桑姆. 苦马豆内生真菌分离鉴定及其生物碱成分分析[D]. 杨凌:西北农林科技大学,2015:34-37
[15]苏丹丹,刘玉萍,刘涛,等. 苦马豆叶绿体基因组结构及其特征分析[J]. 植物研究,2022,42(3):446-454
[16]吴松权,王立平,孙丽娜,等. 黄芪染色体核型分析[J]. 湖北农业科学,2006(5):631-633
[17]张婕妤,张子琦,周薇,等. 横断山高山冰缘带5种黄耆属植物的核型分析[J]. 植物科学学报,2023,41(1):63-69
[18]华利源,王轩,钱关泽. 苹果属植物的核型分析[J]. 聊城大学学报(自然科学版),2019,32(6):97-110
[19]左青青,钱露,高瑜,等. 全缘千里光的染色体核型分析及其系统植物学特征[J]. 遵义医科大学学报,2020,43(3):300-304
[20]RANE V A,PATEL B B,GEORGE J. Karyotype analysis of ten species of Ipomoea jacq[J]. Cytologia,2012,77(2):239-249
[21]顾志建,王丽,孙航,等. 青藏高原一些种子植物的核型研究[J]. 云南植物研究,1993(4):377-384,437-439
[22]白俐,刘兵兵,汝大福. 苦参属6种植物的核型分析[J]. 陕西大学学报(自然科学版),2023,46(2):478-486
[23]杨萍,苏旭,刘玉萍,等. 扇穗茅不同居群染色体数目及核型分析[J]. 草地学报,2022,30(7):1712-1720
[24]孙祥岭,马艳红,姚立蓉,等. 盐生草和白茎盐生草染色体核型分析及盐生草基因组大小预测[J]. 甘肃农业大学学报,2023,58(5):191-198
[25]徐波,孙文光,李志敏. 青藏高原5种石竹科垫状植物的核型研究[J]. 西北植物学报,2020,40(7):1157-1163
[26]郑慧敏,毛培胜,黄莺,等. 牧草染色体核型和带型分析研究进展[J]. 草地学报,2015,23(1):14-20
[27]李国泰. 百合科4种植物染色体的核型比较[J]. 中国林副特产,2017(6):21-25
[28]李懋学. 植物的染色体组和组型分析[J]. 生物学通报,1981(4):18-21
[29]KOU S R,WANG T T,HUANG T C. Karyotype analysis of some formosan gymnosperms[J]. Taiwania,1972,17(1):66-80
[30]ARANO H. Cytological studies in subfamily carduoideae (compositae) of Japan IX. The karyotype analysis and phylogenic considerations on Pertya and Ainsliaea[J]. Shokubutsugaku Zasshi,1963,76(895):32-39
[31]STEBBINS G L. Chromosomal evolution in higher plants[M]. London:Edward Arnold,1971(48):85
[32]邢世岩,高进红,姜岳忠,等. 银杏特异种质核型进化趋势[J]. 林业科学,2007,43(1):21-27
[33]李晓莉,贺新桠,肖鑫辉,等. 5个木薯品种染色体核型与聚类分析[J]. 热带作物学报,2019,40(1):79-86
[34]杨慧芳. 冰草居群核型变异研究[D]. 雅安:四川农业大学,2016:28-31
[35]雷海英,侯沁文,白凤麟,等. 八种不同产地苦参的染色体数目及核型分析[J]. 植物生理学报,2019,55(7):967-974
[36]杨光穗,冷青云,王呈丹,等. 16个红掌品种的核型分析[J]. 热带作物学报,2016,37(12):2283-2287
[37]李明阳,刘彦泽,王鑫,等. 铁线莲属21个类群的染色体核型分析[J]. 广西植物,2022,42(1):78-89
[38]张力天,刘德梅,刘博,等. 西藏自治区日喀则市白草核型分析(英文)[J]. 草地学报,2021,29(7):1589-1593
[39]李红,赵淑文,乌都苏布德. 骆驼刺染色体数目及核型分析[J]. 干旱区资源与环境,2012,26(8):176-179
[40]牛牧歌,郑芹,李际红,等. 流苏树山东不同居群的染色体核型分析[J]. 西北植物学报,2023,43(4):582-591
[41]DU Y,BI Y,ZHANG M,et al. Genome size diversity in Lilium (Liliaceae) is correlated with karyotype and environmental traits[J]. Frontiers in Plant Science,2017(8):1303
[42]张建波,白史且,张新全,等. 川西北高原12个垂穗披碱草居群的核型研究[J]. 西北植物学报,2008,28(5):946-955
[43]孙勃,辜金花,童袁桃,等. 不同产地木耳菜的染色体核型分析[J]. 西北植物学报,2017,37(8):1525-1532
[44]孙立民,王文英,邢世岩,等. 古银杏半同胞家系染色体核型分析及进化趋势研究[J]. 西南林业大学学报(自然科学),2017,37(5):15-21
[45]宛涛,孙启忠,蔡萍,等. 内蒙古不同生态区冷蒿染色体核型观察[J]. 西北植物学报,2011,31(3):456-461
[46]王亚男,刘玉萍,刘雪利,等.沙鞭不同居群染色体数目及核型分析[J]. 西北植物学报,2021,41(9):1489-1499
[47]PASZKO B. A critical review and a new proposal of karyotype asymmetry indices[J]. Plant Systematics and Evolution,2006,258(1-2):39-48
[48]秦进. 基于树木年轮的秦岭林线典型树种对气候的响应与区域气温重建[D]. 西安:西北大学,2018:1
[49]胡延萍,谢小龙,冯海生,等. 唐古特大黄染色体的核型分析[J]. 中国药学杂志,2007,42(4):258-260
(责任编辑 闵芝智)
猜你喜欢
科学之谜(2019年3期)2019-03-28 10:29:44
科学之谜(2018年8期)2018-09-29 11:06:46
湖北农业科学(2016年21期)2017-03-18 15:09:28
江苏农业科学(2016年8期)2017-02-15 20:03:31
安徽农学通报(2016年24期)2017-01-12 20:34:23
大经贸(2016年9期)2016-11-16 16:16:46
中国市场(2016年33期)2016-10-18 12:16:58
科技视界(2016年20期)2016-09-29 12:32:48
恋爱婚姻家庭·养生版(2016年9期)2016-09-07 11:25:01
企业导报(2016年9期)2016-05-26 21:08:14
