
太行山地区栓皮栎不同种源种子萌发及苗期生长特性
2024-09-28 00:00邢学丁任俊杰殷小娟霍巍高卓田康则全张星伟张卫强
林业科技 2024年5期

摘要: 为研究太行山地区栓皮栎种源间的遗传变异,为栓皮栎新品种选育提供种质资源,对6个种源栓皮栎种子形态指标(千粒重、种长、种宽、种形指数)、萌发特性(发芽率、发芽势、发芽指数)及当年生苗期生长性状进行分析。结果表明,种子形态指标和萌发特性在不同种源间均存在显著差异,因此栓皮栎良种选育具有很大地潜力。千粒重、种长、种宽、种形指数分别为:2897.53~5402.60 g、19.38~22.69 mm、15.97~19.51 mm、1.14~1.34;发芽率、发芽势、发芽指数、苗高分别为:44.45~91.11%、16.67~45.55%、1.07~2.86、47.49~63.99 cm。利用模糊隶属函数对千粒重、发芽率、发芽势、发芽指数和苗高综合分析,种源种实品质依次为:河南林州>山西夏县>河南修武>天津蓟州>北京海淀>河北易县。
关键词: 栓皮栎; 种源; 萌发特性; 形态指标; 模糊隶属函数
中图分类号: S 722. 1 + 3 文献标识码: A 文章编号:1001 - 9499(2024)05 - 0001 - 05
Seed Germination and Seedling Growth Characteristics of Different Provenances of Quercus variabilis in Taihang Mountains
Abstract In order to study the genetic variation among seed sources and provide germplasm resources for breeding new varieties of Quercus variabilis sinensis in Taihang Mountains, the seed morphological indexes (1000-grain weight, seed length, seed width, seed shape index), germination characteristics (germination rate, germination potential, germination index) and seedling growth traits of 6 seed sources were analyzed. The results showed that there were significant differences in seed morphological indexes and germination characteristics among different provenances, so there was a great potential for the breeding of Quercus variabilis. The 1000 grain weight, seed length, seed width, and seed form index were 2897.53~5402.60 g, 19.38~22.69 mm, 15.97~19.51 mm and 1.14~1.34; The germination rate, germination potential, germination index and seedling height were 44.45~91.11%, 16.67~45.55%, 1.07~2.86 and 47.49~63.99 cm. The comprehensive analysis of 1000 grain weight, germination rate, germination potential, germination index and seedling height by using fuzzy membership function shows that: Linzhou in Henan>Xia Xian in Shanxi>Xiuwu in Henan>Jizhou in Tianjin>Haidian in Beijing>Yi Xian in Hebei.
Key words Quercus variabilis; provenance; characteristic of germination; index of form; fuzzy membership function
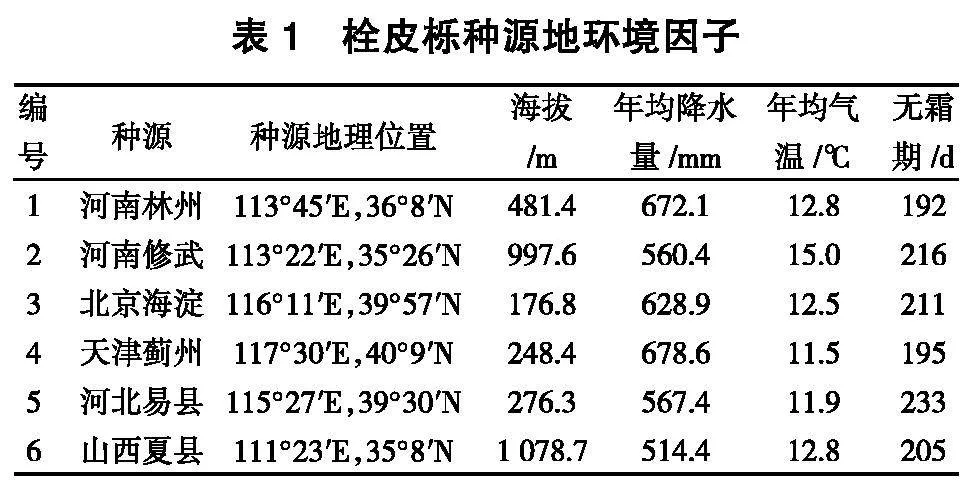
栓皮栎(Quercus variabilis)为壳斗科栎属落叶乔木,在我国广泛分布,北起辽宁省南部,南至云南省中部,分布的海拔上限自北向南不断升高,占全国总面积的44.1%[ 1 , 2 ]。其适应性强,耐干旱、瘠薄土地,在山区、丘陵均可栽植,是荒山和困难立地造林的先锋树种和水土保持树种[ 3 ]。栓皮栎也是重要的软木资源和食用菌、天麻、木炭生产的优良树种,具有比重小、浮力强、弹性好、不透水、不透气等优良特性,是制作高档酒瓶瓶塞的必选原料[ 4 ]。
种子是农林生产的基本生产资料,种子活力是种子重要的播种品质,高活力种子具有明显的生长优势和生产潜力,对林业发展具有极其重要的意义[ 5 ]。国内外学者对栎类种子的贮藏[ 6 ]、萌发[ 7 - 9 ]、育苗技术[ 10 - 11 ]、苗期评价[ 12 - 14 ]及群体遗传多样性[ 15 - 18 ]进行了大量的研究,但对栓皮栎种子种源特征和环境因子的综合研究较少。林木种源因分布区的温度、降水量、光照及海拔、纬度等环境因素的影响,在种子性状、种子营养成分方面存在一定的地理变异[ 20 ]。本研究比较分析了太行山及天津地区栓皮栎不同种源种子形态指标、萌发特性及苗期生长,以期为栓皮栎国家林木种质资源库建设及优良种源的早期筛选提供理论依据。
1 材料与方法
1. 1 试验材料
2021年8~9月份收集栓皮栎太行山及天津地区典型自然分布区[ 21 ]的河南林州、河南修武、北京海淀、天津蓟州(对照)、河北易县、山西夏县5省份6个种源优质种子(表1),经查询当地气象部门发布的官方数据,2021年天气情况与往年无明显差异。每个种源采种20株以上等比例混合种子共计1000粒,选择生长健壮、树形完整的30年以上稳定结实的母树采种,各采种母树间距为树高的5倍以上。将不同源种子在七里亭示范场内育苗,进行正常的田间管理,待生长期结束后各种源随机调查 30 株栓皮栎的苗高、地径及成熟叶叶长、成熟叶叶宽,并分别称取苗木叶片、茎和根的鲜干质量。
1. 2 试验方法
种子质量用电子天平测量,即从待测种子中随机挑选100粒种子称重,3次重复,并计算出千粒重(保留两位小数)。从待测种子中随机挑选100粒种子,用电子游标卡尺测量种子的种长与种宽(保留两位小数),3次重复,并计算种形指数。种子的发芽率采用上海一恒科学仪器有限公司生产的MGC—400B机型光照培养箱测定,每个种源随机挑选50粒饱满无虫害种子,均匀放在垫有两层滤纸的一次性塑料培养皿中,3次重复。光照培养箱设置为黑暗8 h,27 ℃;60%光照16 h,27.5 ℃[ 5 ]。从置床开始每隔24 h统计1次发芽率,统计标准为胚根长度与种子等长时记为发芽[ 21 ]。
当连续5 d发芽粒数不足供试种子总数1%时,结束试验并计算发芽率、发芽势、发芽指数。计算公式如下:
种形指数=种长(纵径)/种宽(横径)
发芽率=总的发芽粒数/供试种子总数×100%
发芽势=规定天数内的发芽粒数/供试种子总数×100%
发芽指数=∑(Gi /Di)
式中:Gi为第i天发芽种子数;Gi为天数。
变异系数CV=标准差SD/平均值Mean×100%。
1. 3 数据分析
用Excel对原始数据进行整理,用SPSS20.0进行方差分析、差异显著性检验及相关性分析。数据用Mean(均值)±SD(标准差)表示。不同种源种子性状的综合评价采用模糊数学隶属函数法,公式为F(X)=(X-Xmin)/(Xmax-Xmin),式中F(X)为隶属函数值,X为指标测定值,Xmax、Xmin为测定指标的最大值和最小值[ 23 ]。
2 结果与分析
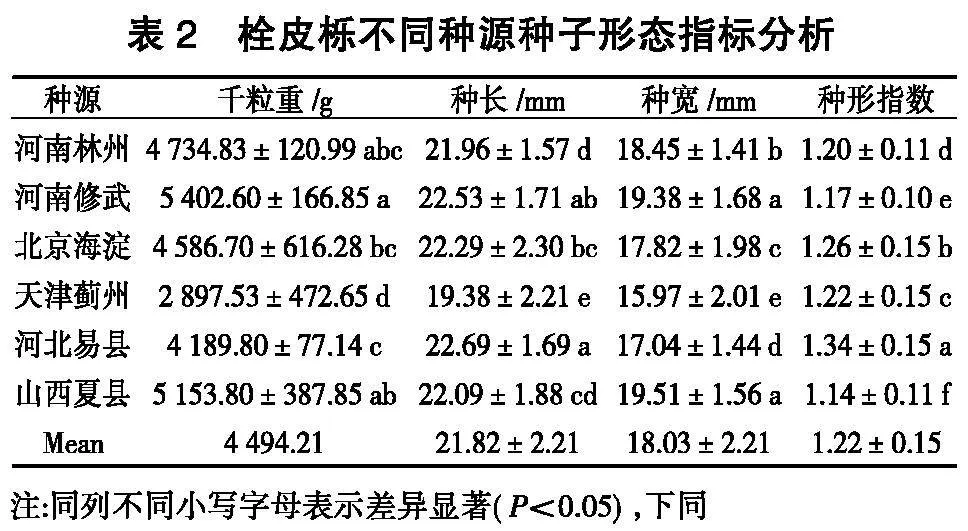
2. 1 栓皮栎不同种源种子形态及变异分析
由表2可知,不同种源种子千粒重、种长、种宽、种形指数存在显著性差异,千粒重为2 897.53~
5 402.60 g,种长为19.38~22.69 mm,种宽为15.97~
19.51mm,种形指数为1.14~1.34。其中河南修武种源种子的千粒重最大,为5 402.60 g;河北易县种源种子的种长最长,为22.69 mm;山西夏县种源种子的种宽最大,为19.51 mm;河北易县种源种子的种形指数最大,为1.34,种子呈长圆形;山西夏县种源种子的种形指数最小,为1.14,种子呈扁圆形。
变异系数越大,说明其适应环境的能力越强,引种越易成功。由表3可知,不同种源种子千粒重、种长、种宽、种形指数存在较大的变异,千粒重变异系数为1.84%~16.31%,种长变异系数为7.13%~
11.39%,种宽变异系数为7.62%~12.58%,种形指数变异系数为8.77%~12.17%。其中天津蓟州和北京海淀种源种子的千粒重、种长、种宽和种形指数变异系数均超过了10%。不同种源种子形态变异大小依次为:天津蓟州>北京海淀>山西夏县>河北易县>河南修武>河南林州。
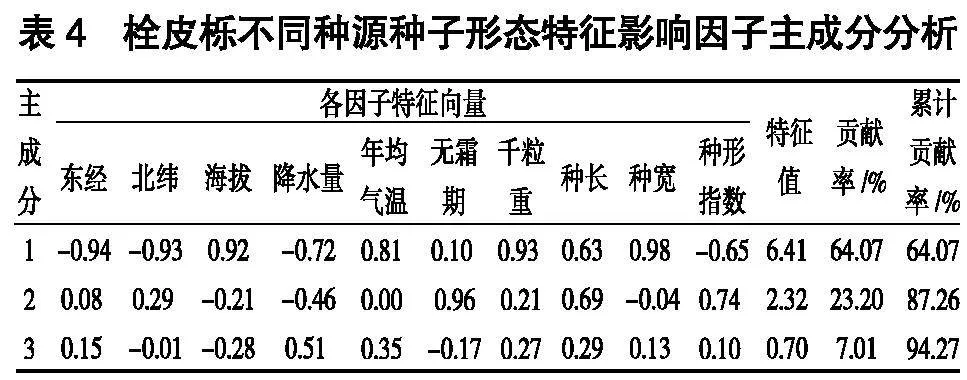
2. 2 种子形态特征影响因子主成分分析
以6个栓皮栎种源混合种子为样本单元,将种子形态(千粒重、种长、种宽、种形指数)、经纬度和海拔进行主成分分析。由表4可知,第1、第2、第3主成分的贡献率分别达到了64.07%、23.20%、7.01%,累计贡献率为94.27%。在第1主成分上特征向量较大的因子是种子的种宽和千粒重,反应种子的质量;第2主成分是无霜期,反应的是环境因子;第3主成分是降水量,反应的是气象因子。由此可知,影响栓皮栎种子形态的首要因素是种子质量,其次是无霜期和降水量。
2. 3 栓皮栎不同种源种子萌发特性分析
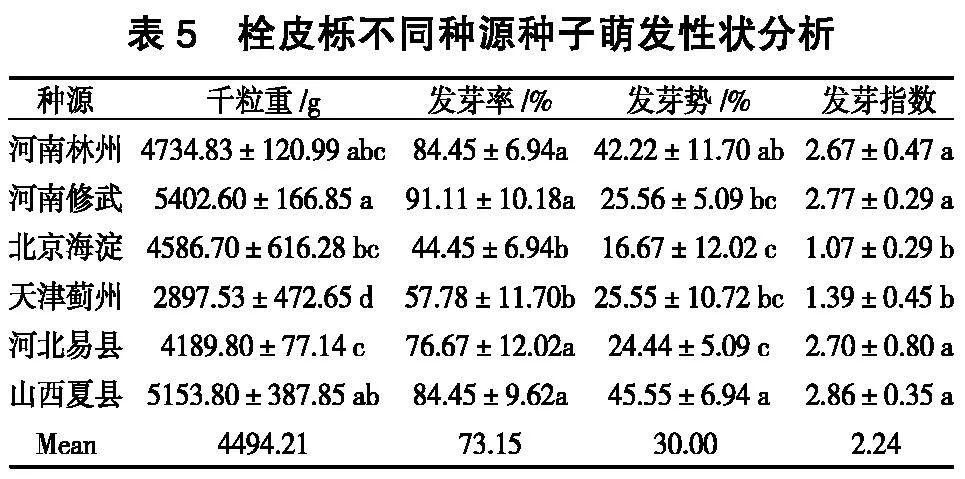
千粒重是评价种子综合品质的一个重要指标,在一定程度上决定着出苗率和植株的生长状况;发芽率越高代表种子播种品质越好;发芽势和发芽指数越高,代表种子萌发越快、出苗越整齐。由表5可知,不同种源间种子的千粒重、发芽率、发芽势、发芽指数间存在显著差异,千粒重最高的河南修武种源发芽率同样也是最高的。种子千粒重范围2 897.53~5 402.60g,千粒重最大的河南修武种源是千粒重最小的天津蓟州种源的1.86倍,高于种源千粒重均值的共有4个,分别是河南修武、山西夏县、河南林州和北京海淀。种子发芽率范围为44.45%~91.11%,发芽率最高的河南修武种源是发芽率最低的北京海淀种源的2.05倍,高于种源发芽率均值的共有4个,分别是河南修武、河南林州、山西夏县和河北易县。种子发芽势范围为16.67%~ 45.55%,发芽势最高的山西夏县种源是发芽势最低的北京海淀种源的2.72倍,高于种源发芽势均值的共有2个,分别是山西夏县和河南林州。种子发芽指数范围为1.07~2.86,发芽指数最高的山西夏县种源是发芽指数最低的北京海淀种源的2.67倍,高于发芽指数均值的共有4个,分别是山西夏县、河南修武、河北易县、河南林州。
2. 4 不同种源苗期生长性状
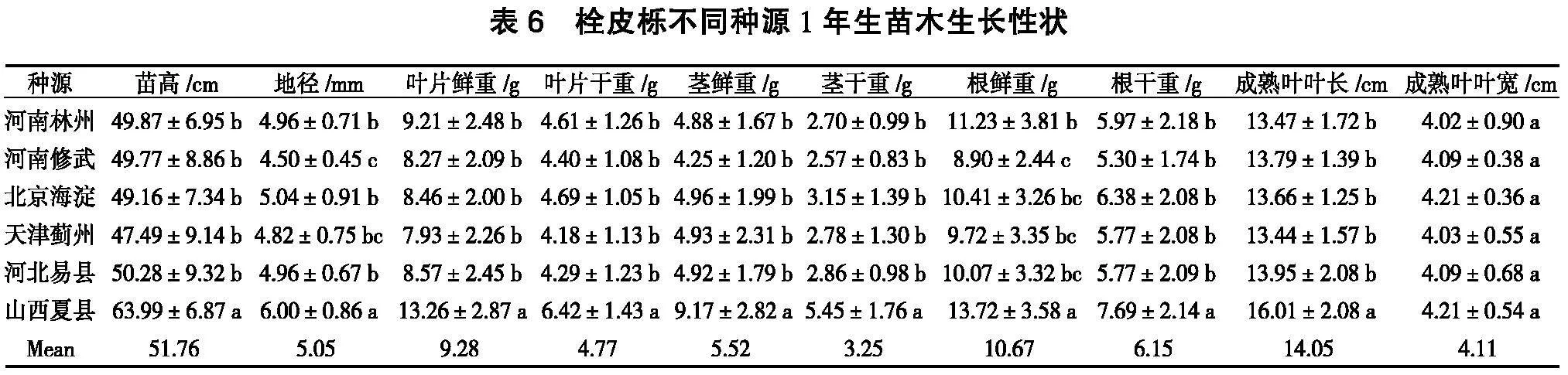
由表6可知,山西夏县种源的苗高、地径、叶片鲜重、叶片干重、茎鲜重、茎干重、根鲜重、根干重、成熟叶叶长与其他种源差异显著,成熟叶叶宽间无显著差异。仅有山西夏县种源的苗高、地径、叶片鲜重、叶片干重、茎鲜重、茎干重、成熟叶叶长、成熟叶叶宽高于均值,其他种源均低于种源均值。
2. 5 栓皮栎不同种源种苗品质分析
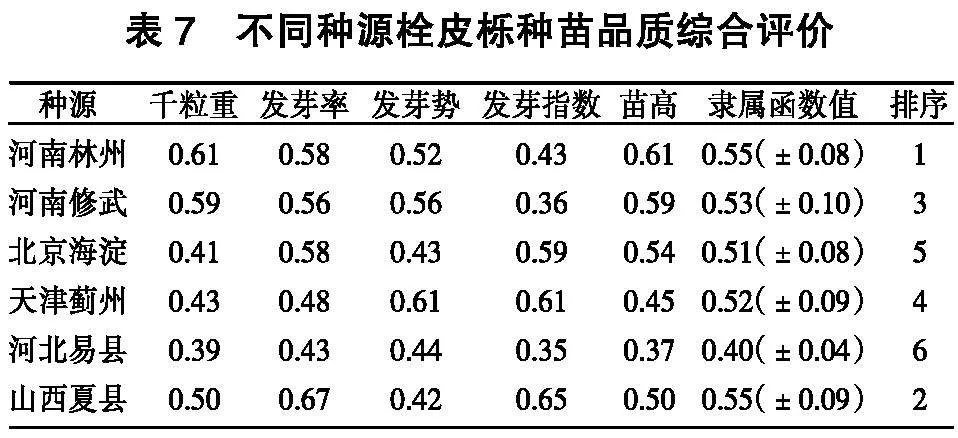
利用模糊隶属函数对千粒重、发芽率、发芽势、发芽指数及苗高、基径综合分析,对栓皮栎种源种苗综合品质进行排序(表7):河南林州>山西夏县>河南修武>天津蓟州>北京海淀>河北易县。
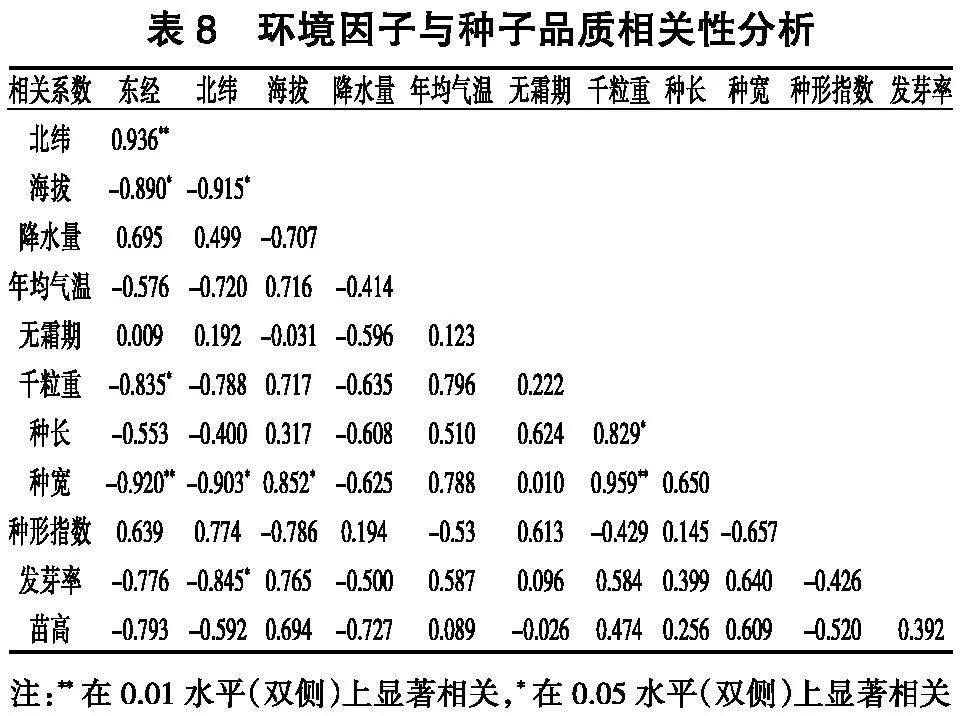
2. 6 环境因子与种苗品质的相关分析
由表8可知,经度和纬度与种宽分别呈极显著负相关和显著负相关,经度与千粒重呈显著负相关,海拔与种宽呈显著正相关,千粒重与种长和种宽呈极显著正相关,发芽率与纬度呈显著负相关。而种形指数与经纬度呈现不显著正相关,说明种子形状主要受到遗传控制。
3 结论与讨论
通过对栓皮栎6个种源种子进行方差分析和相关分析发现,千粒重、发芽率、发芽势、发芽指数均存在显著差异,且种源间变异要大于种源内。这与国内外同类研究得到了类似结论,Servet Caliskan[ 23 ]通过对不同种源青东栎(Quercus ilex)的种实特性和萌发情况分析,发现不同种源间各性状均存在显著差异,在20 ℃的恒温条件下,发芽率最高;陈素传等[ 24 ]通过对栓皮栎28个种源研究发现,不同种源栓皮栎的差异主要是由种子千粒质量、宽度和长度决定,初步选出云南勐海等8个种源为优异种质资源,本研究得到了类似的研究结论,发现千粒重与种长、种宽存在显著正相关。方差分析发现,千粒重、种长、种宽的变异主要存在组内,这与Zhang et al.[ 18 ]研究麻栎种源的遗传变异得到了类似的结论,Zhang et al.利用12对微卫星引物对中国28个麻栎种源地672个家系进行了遗传变异分析,发现84%的变异发生在种源间;而李文文[ 25 ]研究蒙古栎种源变异发现,表型变异主要存在于种间。出现研究结果的差异,可能是因为群体所处的环境不同有关。常恩福等[ 26 ]以铁橡栎4个种源33个家系为研究材料,发现影响铁橡栎种子形态特征的首要因子是种子的单粒重、千粒重和种形指数,种子形状主要受遗传控制。胡玉珠[ 27 ]通过研究蒙古栎不同种源种实形态,发现各种源间的种实千粒重、种长、种宽均存在显著差异。因此可以将种实形态作为区分不同种源的依据。
种子的变异系数越大,遗传多样性越高,说明其适应环境的能力越强,引种越易成功。不同种源种子千粒重、种长、种宽、种形指数存在显著差异,天津蓟州和北京海淀种源种子的千粒重、种长、种宽和种形指数变异系数均超过了10%,这与李贵芬[ 28 ]、方芳[ 5 ]得到的研究结果类似。李贵芬等对蒙古栎7个种源地种子形态指标变异研究发现,山东烟台种源蒙古栎百粒重变异系数为12.88%;方芳对5个种源栓皮栎种子形态研究发现,安徽大别山种源种子长、种子宽变异均超过了10%。
千粒重、萌发特性和苗高是评价种苗综合品质的重要指标,利用模糊隶属函数对千粒重、发芽率、发芽势、发芽指数和苗高综合分析,得到种源种苗品质依次为:河南林州>山西夏县>河南修武>天津蓟州>北京海淀>河北易县。
环境因子与种子品质相关分析发现,经纬度与种宽呈显著负相关,经度与千粒重呈显著负相关,海拔与种宽呈显著正相关,千粒重与种长和种宽呈极显著正相关。不同种源种子形态指标存在显著差异,千粒重变异最为丰富[ 27 - 29 ],究其原因是种子的大小和质量与外界环境密切相关,是长期自然选择的结果,也是基因型与环境相互作用的结果,结合种子形态指标可以为野外采种初步筛选优良种源,选育高产丰产家系提供理论依据。林木育种界普遍认为,林木树龄越大,选择结果越可靠,但由于林木子代测定周期长,赵世杰[ 30 ]等对落叶松的生长情况进行了早期和晚期的研究,试验结果证明了早期筛选的可行性。本试验对栓皮栎不同种源种实特性及当年生长性状进行调查与分析,后续应结合测定林生长情况以及分子生物学鉴别进行进一步的验证和二次选择,以期为栓皮栎国家林木种质资源库优良种源的筛选提供依据。
参考文献
[1] 雷静品, 肖文发, 刘建锋. 我国栓皮栎分布及其生态学研究[J].世界林业研究, 2013, 26(4): 57 - 62.
[2] 吴肖. 栓皮栎软木活性炭的制备及其染料吸附性能研究[D].长沙: 中南林业科技大学, 2022.
[3] 丁伯让. 皖东地区栎类资源及其开发利用[J]. 安徽林业科技, 2006(1): 75 - 76.
[4] 唐晓倩. 北方主要落叶栎类种子形态特征和养分含量的研究[D]. 泰安: 山东农业大学, 2012.
[5] 方芳. 刺槐、 栓皮栎种子活力与萌发特性研究[D]. 北京: 北京林业大学, 2013.
[6] 赵海燕. 不同贮藏处理措施对栓皮栎种子的影响[J]. 山西林业科技, 2022, 51(2): 34 - 36.
[7] 赵娟. 气候变化背景下模拟温度升高和降水变化对蒙古栎种子萌发和幼苗生长的影响[D]. 哈尔滨: 东北林业大学, 2013.
[8] Nicolaza Pariona,Arturo I. Martínez,et al. Effect of magnetite nanoparticles on the germination and early growth of Quercus macdougallii[J]. Science of the Total Environment, 2017, 575.
[9] Lei-Deng,Wen-Hui Zhang,Jin-Hong Guan. Seed rain and community diversity of Liaotung oak (Quercus liaotungensis Koidz) in Shaanxi, northwest China[J]. Ecological Engineering, 2014, 67.
[10] 陆秀君, 李宏祎, 艾万峰, 等. 容器规格、基质配比和缓释肥对蒙古栎容器苗质量的影响[J]. 东北林业大学学报, 2020, 48(7): 17 - 22.
[11] 李亚男, 李东胜, 许中旗, 等. 栎属不同种源树种的出苗及生长过程[J]. 西北林学院学报, 2014, 29(4): 139 - 144.
[12] 李璇, 薛美玲, 胡荣, 等. 麻栎与栓皮栎不同种源苗期生长差异性及评价[J]. 东北林业大学学报, 2021, 49(7): 9 - 15.
[13] 陈素传, 王陆军, 蔡新玲, 等. 栓皮栎不同种源苗期变异与初步选择[J]. 林业科技开发, 2012, 26(4): 22 - 26.
[14] 李文文. 蒙古栎种源变异及无性鳘殖研究[D]. 北京: 中国林业科学研究院, 2010.
[15] María Cristina Romero-Rodríguez, et al.Toward characterizing germination and early growth in the non-orthodox forest tree species Quercus ilex through complementary gel and gel-free proteomic analysis of embryo and seedlings[J]. Journal of Proteomics, 2018, 197.
[16] 徐小林, 徐立安, 黄敏仁, 等. 栓皮栎天然群体SSR遗传多样性研究[J]. 遗传, 2004(5): 683 - 688.
[17] 韩照祥. 陕西地区栓皮栎种群区域变异性与遗传多样性的研究[D]. 杨凌: 西北农林科技大学, 2004.
[18] 李军民, 丁小飞, 陈红林, 等. 长阳栓皮栎天然群体遗传多样性的等位酶分析[J]. 湖北林业科技, 2012(1): 3 - 4 + 12.
[19] Zhang Yuanyan,Fang Yanming ,Yu Mukui ,et al. Molecular characterization and genetic structure of Quercus acutissima germplasm in China using microsatellites[J]. Molecular biology reports, 2013, 40(6).
[20] 颜启传. 种子检验的原理和技术[M]. 北京: 农业出版社.1992, 204 - 209.
[21] 李迎超. 木本淀粉能源植物栓皮栎与麻栎资源调查及地理种源变异分析[D]. 北京: 中国林业科学研究院, 2013.
[22] 张国君, 代波, 孙海军, 等. 华北落叶松25个种源种子萌发及苗期生长特性[J]. 东北林业大学学报, 2015, 43(7): 11 - 14 + 83.
[23] 邢学丁. 华北落叶松2代种子园结实和子代生长双优无性系的筛选[D]. 秦皇岛: 河北科技师范学院, 2020.
[24] Caliskan S. Germination and seedling growth of holm oak (Quercus ilex L.): effects of provenance, temperature, and radicle pruning[J]. iForest- Biogeosciences and Forestry, 2014, 7(1).
[25] 陈素传, 肖正东, 金笑龙, 等. 不同种源栓皮栎种子形态和营养成分差异分析[J]. 林业科技开发, 2012, 26(1): 17 - 21.
[26] 常恩福, 张清, 肖桂英, 等. 铁橡栎不同种源及家系种子的形态特征及变异[J]. 种子, 2020, 39(6): 53 - 58.
[27] 胡玉珠. 4个不同种源蒙古栎种实形态分析[J]. 吉林林业科技, 2020, 49(1): 1 - 3.
[28] 李贵芬, 李昕蔓, 刘朝华, 等. 不同种源蒙古栎种子形态指标比较与分析[J]. 现代农业科技, 2021(11): 114 - 116.
[29] 任俊杰, 庞新博, 刘昭阳, 等. 不同种源蒙古栎种子表型性状的多样性[J]. 浙江农林大学学报, 2022, 39(6): 1221 - 1228.
[30] 赵士杰, 杨俊明, 李文治. 华北落叶松优树子代测定初报[J].河北林业科技, 1988(3): 1 - 5.
