
不同品种马铃薯幼苗对NaHCO3 胁迫的响应及耐碱能力指标筛选
2024-07-09 09:41:05逯春杏刘玉龙吴娟苑志强胡戎朔张胜王晓娇曹春梅张必周逯海林
植物营养与肥料学报 2024年4期
逯春杏 刘玉龙 吴娟 苑志强 胡戎朔 张胜 王晓娇 曹春梅 张必周 逯海林


摘要: 【目的】探究不同马铃薯品种对NaHCO3 胁迫的响应机制,为选育耐碱性较强的马铃薯品种提供理论依据。【方法】以 V7、后旗红、克新1 号、兴佳2 号、荷兰15 号、康尼贝克6 个品种马铃薯为研究对象,以1/2霍格兰(Hoagland) 营养液为母液进行了水培试验。营养液设置6 个NaHCO3 浓度,分别为0、10、20、30、40、50 mmol/L,选择出苗后7 天的马铃薯幼苗,缓苗3 天后进行碱胁迫处理。胁迫处理7 天后,调查马铃薯幼苗株高、茎粗、主根长,分析叶片含氮量和叶绿素相对含量,分析根系脯氨酸含量、过氧化物酶(POD)活性、过氧化物歧化酶(SOD)活性以及过氧化氢酶(CAT)活性等11 项生理指标。【结果】随着碱胁迫浓度的增加,各品种马铃薯幼苗生长受到不同程度的影响,在碱浓度30、40、50 mmol/L 胁迫处理下,V7 根长较0 mmol/L 处理分别增加了54.37%、46.04%、25.58%,其余5 个品种根长均显著低于0 mmol/L。50 mmol/L 胁迫处理下,各品种株高、茎粗均显著低于碱浓度0 mmol/L 处理。各品种叶片含氮量、叶绿素相对含量随胁迫浓度增加呈现不同的变化趋势,V7、后旗红、克新1 号叶片含氮量均在40 mmol/L 处理下达到峰值,兴佳2 号叶绿素相对含量在50mmol/L 处理下达到峰值,而荷兰15 号、康尼贝克叶片含氮量、叶绿素相对含量均在50mmol/L 处理达到最小值。各品种幼苗根系活力呈先增后降的趋势,碱浓度40 mmol/L 胁迫处理下,V7、后旗红根系活力显著高于0mmol/L,而荷兰15 号的降幅达80% 以上。各品种幼苗根系中丙二醛含量随胁迫程度加剧而增加,但V7 的丙二醛含量始终低于其他品种。各品种幼苗根系CAT 活性、SOD 活性均呈先升后降变化趋势,其中V7、后旗红、克新1 号、兴佳2 号的CAT 活性、SOD 活性高于康尼贝克和荷兰15 号。利用主成分分析计算得到5 个特征值>1 的归一化指标,其对碱胁迫抗性的累积贡献率达80.07%,符合主成分分析要求,因此将11 个指标转化为7 项生理指标,包括:株高、茎粗、叶片含氮量、叶绿素相对含量、脯氨酸含量、POD活性、CAT 活性。【结论】株高、茎粗、叶片含氮量、叶绿素相对含量、脯氨酸含量、POD 活性、CAT 活性这7 项生理指标可作为马铃薯耐碱性能力强弱的评价指标。综合各项指标筛选出耐碱能力较强的品种为V7,后旗红、克新1 号、兴佳2 号耐碱能力中等,康尼贝克与荷兰15 号耐碱性较弱。
关键词: 马铃薯; NaHCO3 胁迫; 耐碱性; 耐碱能力指标
土壤盐渍化已成为当今世界面临的重要环境问题[1] 。中国盐碱地面积居世界第三位[2]。据统计,我国盐碱地面积已超过0.3 亿hm2,其中碱性土壤为87 万hm2[2],涉及 17 个省(市、自治区),主要分布在内蒙古、宁夏、黑龙江、甘肃、青海以及沿海等地[ 3 ]。盐碱化的土壤中含盐量高、碱性大[ 4 ],严重阻碍农作物的正常生长,特别是对适宜生长在弱酸性或中性土壤环境中的作物,导致其种植面积、产量以及品质都受到极大影响,制约农业生产力的发展[5]。研究作物对盐碱胁迫的响应机制,提高作物耐盐碱性,对推动我国盐碱地可持续利用具有重要的科学意义。
马铃薯(Solanum tuberosum L.) 喜酸性土壤,是世界上广为种植的粮菜兼用作物。2015 年,农业农村部将马铃薯确定为第四大主粮作物,提出马铃薯主粮化[6]。有学者认为,马铃薯属于盐碱敏感型作物[7],当土壤中pH>7.0 时,马铃薯产量会降低;有些品种种植在强碱性土壤中,会出现生长缓慢,甚至无法正常出苗,或发生霉变等现象,严重影响其商品价值[8]。刘婕等[7]研究表明,随着土壤中盐分含量的增加,马铃薯克新1 号、青薯9 号、冀张薯12 号等品种的株高均受到不同程度抑制,当盐胁迫浓度达到100 mmol/L 时,马铃薯株高停止增长并伴随茎段褐化现象发生。相关研究表明,东农308、费乌瑞它两个马铃薯品种在含有NaCl、 NaHCO3 混合盐的MS 培养基中胁迫处理,二者叶片中叶绿素含量、过氧化氢酶(CAT) 活性均降低,丙二醛(MDA)含量、超氧化物歧化酶(SOD) 活性、过氧化物酶(POD) 活性均升高[9];随着土壤中盐碱含量的增加,马铃薯叶片中脯氨酸含量、可溶性糖含量较对照升高[8],而可溶性蛋白含量降低[10]。
碱胁迫条件下,由于pH 升高会直接破坏植物的根系,导致其对养分、矿物质的吸收与利用率降低,破坏植株内部离子平衡,因此会给农作物带来更大的危害[11]。现阶段有关马铃薯响应盐碱胁迫的研究主要集中在以NaCl 中性盐、NaCl 与Na2SO4、Na2CO3 以及NaHCO3 混合盐等方面,而针对幼苗期马铃薯单独响应某种碱性盐胁迫以及不同品种间生理响应差异的研究鲜见报道。本研究以预试验筛选出的6 个马铃薯品种为试验材料,用1/2 Hoagland营养液为对照(CK),采用水培试验法,探究不同程度NaHCO3 碱性胁迫对马铃薯幼苗生长及相关生理指标的影响,旨在阐释马铃薯耐碱机理,为选育耐碱性较强的马铃薯品种及生产应用提供理论依据。
1 材料与方法
1.1 试验地点与材料
试验于内蒙古自治区农牧业科学院人工智能温室内进行,供试马铃薯品种为V7、后旗红、克新1 号、兴佳2 号、荷兰15 号以及康尼贝克,均由内蒙古自治区农牧业科学院特色作物研究所马铃薯研究中心提供。
1.2 试验设计与方法
试验处理:试验以1/2 Hoagland 营养液为母液,分别配制成含NaHCO3 0、10、20、30、40、50 mmol/L 6 种不同浓度胁迫处理溶液,其对应的pH 分别为6.20、7.39、8.01、8.22、8.43 和8.57。
试验采用盆栽种植与营养液胁迫相结合的方式。盆栽种植选用上口径20.5 cm,下口径18.5 cm,高18 cm 的塑料花盆,盆内装有由蛭石∶草炭∶珍珠岩以4∶1∶1 体积比混合而成的基质至盆上沿以下5 cm 处。每盆播种不同马铃薯品种微型薯5 粒,每个品种栽种30 盆,共计180 盆。置于温度25℃±2℃、光照/黑暗时间为16 h/8 h 的条件下培养发芽。从出苗开始,每天上午8:00—9:00,用200 mL1/2Hoagland 营养液浇灌幼苗,以提高其适应性。待出苗7 天后,各品种选取生长整齐一致幼苗每6 株为1 次重复,固定在已打孔的泡沫板上,置于装有300 mL 1/2 Hoagland 营养液的水培盆内,使其根系完全浸没在营养液中缓苗3 天。缓苗后,将水培盆内的营养液更换为等体积(300 mL) 的不同浓度NaHCO3 胁迫液进行胁迫培养。水培胁迫管理条件:温度保持在23℃~25℃,利用增氧泵每天保证根系通气6 h,光照/黑暗时间为14 h/10 h,各处理每24 h更换1 次相应浓度NaHCO3 胁迫液。每个处理6 株幼苗,3 次重复,共计108 盆,648 株。胁迫处理7天后,取样测定各品种、各处理马铃薯的形态指标、生物量、光合指标等,同时采集各品种幼苗根系及叶片,用蒸馏水冲洗3 次,滤纸吸干表面多余水分,用锡箔纸包好迅速放入液氮中速冻,所有样品做好标记后置于–80℃ 超低温冰箱中保存,用于生理活性指标测定。
1.3 测定项目及方法
1.3.1 幼苗叶片光合指标测定
利用便携式植株养分速测仪(TYS-3N) 活体测定胁迫7 天后各品种幼苗叶片中叶绿素相对含量(SPAD 值)、氮素含量。测量时间为上午8:00—10:00。具体测定方法:每处理随机选取生长正常且长势一致幼苗5 株,逐株从顶芽向下数第3、4 片完全展开叶,用TYS-3N 快速轻轻夹住叶片,3 秒后同时读取SPAD 值、氮素含量。每个叶片重复测量3 次,取均值记录。
1.3.2 幼苗形态指标测定
将各品种、各处理测定叶绿素及氮素含量后的5 株幼苗从泡沫板上取出,用蒸馏水冲洗干净并吸干表面水分,采用直尺逐株快速测定幼苗株高、主根长度,利用游标卡尺测量茎基部最粗处直径,读取直径平均值。
1.3.3 幼苗生理活性指标测定
选取置于?80℃ 超低温冰箱保存的根系、叶片样品,用于各项生理活性指标测定。根系活力采用氯化三苯基四氮唑(TTC)法[12]测定;丙二醛(MDA)含量采用硫代巴比妥酸法[13]测定;超过氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT) 光还原法[14]测定;过氧化氢酶(CAT)活性采用H2O2 法[15]测定;过氧化物酶(POD)活性采用愈创木酚比色法[13]测定;脯氨酸含量采用磺基水杨酸法[16]测定,以上所有测定指标均为3 次重复。
1.4 数据处理
使用 Microsoft Excel 2019 软件对数据进行整理作图,使用 SPSS 22.0 软件进行主成分分析和单因素方差分析(ANOVA),显著水平为0.05。
2 结果与分析
2.1 NaHCO3 胁迫对不同马铃薯品种幼苗生长的影响
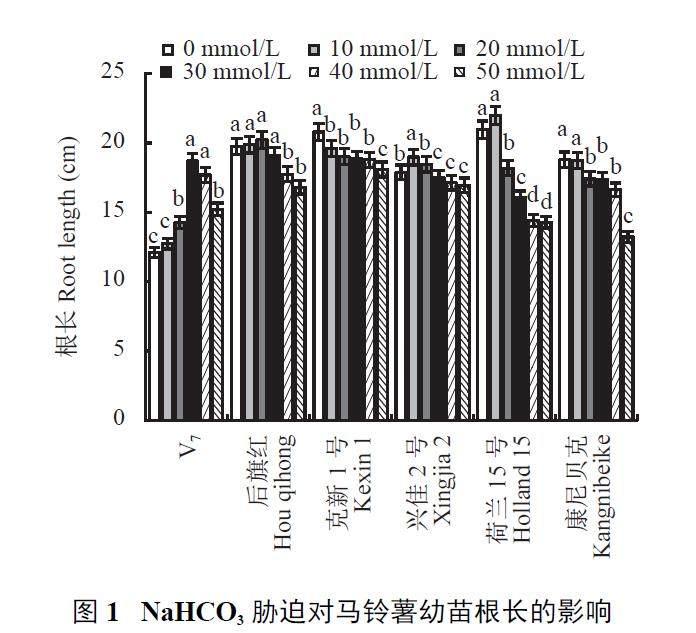
2.1.1 对幼苗根系长度的影响
由图1 可见,随着NaHCO3 胁迫浓度的增加,V7 主根长呈明显的先升后降趋势,后旗红、兴佳2 号、荷兰15 号主根长仅在10 mmol/L 胁迫下有小幅增长,随后逐渐下降,而克新1 号、康尼贝克则随胁迫浓度增加呈下降趋势。在NaHCO3 30、40、50 mmol/L 胁迫浓度处理下,V7 根系长度较0 mmol/L 差异显著,分别增加54.37%、46.04%、25.58%。而后旗红、康尼贝克、克新1 号、兴佳2 号和荷兰15 号在NaHCO3 20 mmol/L及以上浓度胁迫下均显著低于0 mmol/L,当NaHCO3胁迫浓度达到50 mmol/L 时,后旗红、克新1 号、兴佳2 号、荷兰15 号、康尼贝克的主根长分别比0mmol/L 降低14.87%、13.06%、5.14%、31.80%、29.68%,差异显著。由此可见,中、高浓度碱胁迫下,后旗红、克新1 号、兴佳2 号、荷兰15 号、康尼贝克的幼苗根系生长均受到显著抑制,而对V7 根系的生长具有一定促进作用。
2.1.2 对幼苗茎粗的影响
图2 表明,随着NaHCO3胁迫浓度增加,兴佳2 号、康尼贝克幼苗茎粗呈显著下降趋势,10 mmol/L 浓度处理分别较0 mmol/L降低11.97%、15.92%,各处理间差异显著;后旗红、克新1 号、荷兰15 号在低浓度胁迫下无显著差异,而中、高浓度下则显著下降,且处理间差异显著;V7 呈先升后降趋势,在20 mmol/L 浓度处理下茎粗最大, 3 0 m m o l / L 浓度处理次之, 分别比0 mmol/L 增加12.23%、9.21%,二者差异不显著,但显著高于其他各处理。当胁迫浓度达到50 mmol/L时,后旗红、克新1 号、荷兰15 号、兴佳2 号及康尼贝克茎粗达到最小值,分别比0 mmol/L 降低28.60%、22.69%、28.60%、37.24% 和33.79%,差异显著;而V7 茎粗较0 mmol/L 仅减小0.21 mm,降低5.13%,同比其他品种降幅最小。结果表明,V7在低、中碱胁迫条件下,对茎粗生长有明显促进作用,而在高浓度胁迫下所受影响也远小于其他5 个供试品种。
2.1.3 对幼苗株高的影响
从图3 可以看出,各品种仅有V7 的株高随胁迫浓度增加呈先增后降变化趋势,其余5 个品种均呈下降趋势。NaHCO3 10~50 mmol/L 胁迫下,后旗红、克新1 号、兴佳2 号、荷兰15 号、康尼贝克的株高较0 mmol/L 降幅范围分别为4.0%~19.2%、8.4%~11.1%、4.7%~10.5%、8.5%~20.4%、2.1%~7.3%,中、高浓度各处理均显著低于0 mmol/L。表明,上述5 个品种幼苗株高受碱胁迫影响大,耐碱性差。而V7 在NaHCO3 10、20、30 mmol/L 浓度胁迫下,其株高较0 mmol/L 分别增加21%、12.9%、8.9%,差异显著;40、50mmol/L 浓度胁迫下,株高比0 mmol/L 略有降低,但差异均不显著。结果表明,V7 具有较强的耐碱性。
2.2 NaHCO3 胁迫对马铃薯幼苗光合性能的影响
2.2.1 对幼苗叶片含氮量的影响
如图4 所示,V7、后旗红、克新1 号、荷兰15 号、康尼贝克5 个品种幼苗叶片中含氮量均随胁迫浓度增加呈先增后降趋势,V7、后旗红、克新1 号幼苗叶片中含氮量峰值出现在40 mmol/L 胁迫浓度下,分别为3.71、2.76、3.55 mg/g,荷兰15 号、康尼贝克出现在20mmol/L 胁迫浓度下,分别为3.14、2.84 mg/g,各品种峰值均显著高于0 mmol/L,而在50 mmol/L 胁迫浓度下,均显著低于0 mmol/L。整体上来看,后旗红、克新1 号、康尼贝克各处理变幅相对较小,而V7、荷兰15 号变幅较大;兴佳2 号随胁迫浓度增加上升幅度更大,且胁迫处理叶片含氮量均高于0 mmol/L。结果表明,V7、后旗红、克新1 号、兴佳2 号叶片氮含量受碱胁迫的影响小于荷兰15 号、康尼贝克。
2.2.2 对幼苗叶片叶绿素相对含量的影响
如图5所示,NaHCO3 胁迫对各品种叶片中叶绿素相对含量变化趋势与含氮量变化趋势基本一致。V7、兴佳2 号叶片叶绿素相对含量随胁迫浓度增加其增幅较大,当胁迫浓度达到40 mmol/L 或50 mmol/L 时,分别较0 mmol/L 增加64.75%、31.49%,显著高于其他品种。克新1 号各浓度处理叶绿素相对含量变幅较小。荷兰15 号、康尼贝克在中、高浓度下,其叶绿素含量下降。整体而言,V7、兴佳2 号叶片叶绿素合成受碱胁迫影响小,表现出耐碱性较强,而荷兰15 号、康尼贝克耐碱性较差。
2.3 NaHCO3 胁迫对马铃薯幼苗生理特性的影响
2.3.1 对幼苗根系活力的影响
由图6 可知,不同程度碱胁迫对6 个供试品种马铃薯的根系活力均产生了影响。随碱胁迫程度的加剧,各品种根系活力均呈单峰曲线变化趋势。V7、后旗红在各处理下根系活力峰值出现在胁迫浓度40 mmol/L 时,分别较0 mmol/L 增长183.61%、154.55%,差异显著,体现出较强耐碱性。克新1 号、兴佳2 号、康尼贝克峰值出现在20 mmol/L 处理下,较0 mmol/L 分别增长17.0%、451.0%、34.1%,差异显著;荷兰15 号峰值出现在10 mmol/L 处理下,与0 mmol/L 差异不显著。整体而言,V7、后旗红幼苗根系活力在中、高浓度碱胁迫下仍有较大增幅,表现出较强耐碱能力。克新1 号、兴佳2 号、康尼贝克在轻度碱胁迫下,根系活力有所提升,而中、高浓度碱胁迫下,根系活力显著下降,耐碱能力较差;荷兰15 号在中、高浓度碱胁迫下,较0 mmol/L 降幅达80% 以上,差异显著,耐碱性最差。
2.3.2 对幼苗脯氨酸含量的影响
由图7 可知,随着NaHCO3 胁迫浓度增加,V7、兴佳2 号、荷兰15 号、康尼贝克4 个品种植株体内脯氨酸含量均呈逐渐上升趋势,且以50 mmol/L 胁迫浓度下脯氨酸含量最高,其中V7、荷兰15 号显著高于0 mmol/L,兴佳2 号、康尼贝克各处理与0 mmol/L 无显著差异;后旗红、克新1 号随碱胁迫浓度增加脯氨酸含量呈先增后降趋势,后旗红各处理均显著高于0 mmol/L,克新1 号与0 mmol/L 无显著差异。脯氨酸作为一种渗透保护剂,通过提高其含量来抵抗高碱环境。因此,V7、后旗红、荷兰15 号在碱胁迫下幼苗体内脯氨酸提升,表现出较强耐碱能力,而克新1 号、兴佳2 号、康尼贝克各处理脯氨酸含量无显著差异,表明其耐碱能力较差。
2.3.3 对幼苗丙二醛(MDA) 含量影响
由图8 所示,随着碱胁迫浓度增加,除兴佳2 号外,各品种根系中MDA 含量均呈逐渐上升趋势,且于50 mmol/L处理下达到最高值,均显著高于0 mmol/L。低浓度胁迫下,各品种MDA 含量与0 mmol/L 差异不显著, 高浓度胁迫下, 后旗红、克新1 号、荷兰15 号、康尼贝克根系MDA 含量较0 mmol/L 增幅显著,表明受碱胁迫影响其细胞膜破坏程度大,耐碱能力较差。V7、兴佳2 号马铃薯根系丙二醛含量与0 mmol/L 相比虽有增加,但其含量远小于其他品种,表明其耐碱性相对较强。
2.4 NaHCO3 胁迫对马铃薯幼苗根系抗氧化酶系统的影响
2.4.1 对超氧化物歧化酶(SOD) 活性的影响
如图9 所示,NaHCO3 胁迫对各供试品种根系中SOD活性的影响存在差异。随胁迫浓度增加,V7 根系SOD 活性呈逐渐上升趋势,各处理均显著高于0mmol/L;兴佳2 号在40 mmol/L 胁迫下SOD 活性最高,但各处理与0 mmol/L 相比均未达到显著水平。后旗红、克新1 号、荷兰15 号、康尼贝克呈先上升后下降趋势,且均在30 mmol/L 胁迫下SOD 活性达到最大值,其峰值均显著高于0 mmol/L,但随胁迫浓度进一步提高,SOD 活性又逐渐下降。结果表明,V7 和兴佳2 号在碱胁迫下能够诱导根系SOD 活性提高,有利于消除体内O-2,降低对细胞造成伤害,表现出较强耐碱性;其他4 个品种在中、高浓度胁迫下,根系SOD 活性降低,耐碱能力相对较差。
2.4.2 对过氧化物酶(POD) 活性的影响
由图10所示,随着碱胁迫浓度的增加,各品种幼苗根系POD活性均呈先升后降趋势。
从整体来看,V7、后旗红、兴佳2 号根系POD活性相对高于荷兰15 号、康尼贝克及克新1 号。10、20 mmol/L 胁迫处理,V7 根系POD 活性显著高于其他供试品种,较0 mmol/L 分别增加40.01%、50.12%,但差异不显著;30 mmol/L 胁迫处理,后旗红根系中POD 活性为79.25 μg/(g·min),达到峰值,且高于其他5 个品种,与0 mmol/L 差异显著。高浓度NaHCO3 胁迫下,克新1 号根系中POD 活性为9.43 μg/(g·min),为6 个供试品种中最低,比0 mmol/L低62.50%,这表明该品种耐碱性较差。
2.4.3 对过氧化氢酶(CAT) 活性的影响
由图11所示,NaHCO3 胁迫下,6 个供试品种幼苗根系CAT活性变化与POD 活性变化趋势一致,均呈先升后降变化。V7、克新1 号峰值分别出现在30、20 mmol/L浓度下,均显著高于0 mmol/L,在50 mmol/L 浓度下较0 mmol/L 有所下降,但无显著差异;后旗红、兴佳2 号、荷兰15 号、康尼贝克峰值均出现在10 mmol/L 胁迫浓度下,均显著高于0 mmol/L,但50 mmol/L 浓度下均较0 mmol/L 降低。结果表明,一定浓度的碱胁迫能诱导增强马铃薯根系CAT 活性,进而缓解体内H2O2 带来的伤害。V7、克新1 号在中等浓度碱胁迫下仍显著提高了CAT 活性,表现出较强的耐碱能力, 后旗红、兴佳2 号、荷兰15 号、康尼贝克仅在低浓度碱胁迫下根系CAT 活性有所增加,因此耐碱能力表现较差。
2.5 NaHCO3 对不同品种马铃薯幼苗胁迫的主成分分析
为筛选确定马铃薯幼苗耐NaHCO3 碱性胁迫的主要指标,针对6 个供试品种11 项性状指标进行主成分分析(表1),经标准化处理,分析不同马铃薯品种经NaHCO3 胁迫处理后生理生化指标主成分贡献率及其特征向量。以主成分分析得到特征值的累积方差贡献率大于80% 为依据。
由表1 可知,前5 个主成分的特征值分别为3.133、2.055、1.515、1.156、1.038,均大于1.0,贡献率分别为28.48%、18.69%、13.77%、10.51% 和8.62%,特征值前5 位的主成分累积贡献率80.07%,符合主成分分析方法的要求。因此,可以将11 项生理指标转化为7 项新的互相独立的综合指标,用来代替6 个不同马铃薯品种耐碱性盐原始所有指标的绝大部分信息。由表1 可以看出,叶片含氮量、叶绿素相对含量在主成分F1 上表现出较高的载荷量,株高、CAT 活性在主成分F2 上表现出高载荷量,脯氨酸含量、茎粗、POD 活性分别在F3、F4、F5 上具有高载荷量。
由此可知,不同马铃薯品种幼苗叶片中含氮量、叶绿素相对含量、脯氨酸含量、株高、茎粗及根系CAT 活性、POD 活性这7 项指标与其耐碱能力密切相关,可作为马铃薯幼苗期耐碱性鉴定的重要指标。
3 讨论
植株的外部形态和生长状况能够真实直观地反映作物受盐碱胁迫的程度[ 1 5 ],根系主要起到吸收水分、营养运输,保障植株正常生长发育的关键作用[16]。研究表明,当作物在遭受盐碱胁迫时,植株根系会立即感受到外界环境变化,从而减缓水分、养分的运送,因此生物量的积累受到抑制[17],当胁迫强度达到植株自身无法调节时终会导致早衰、死亡等现象发生[18]。本研究发现,6 个供试品种在不同程度碱胁迫下,其主根长、茎粗、株高变化存在差异性。低浓度碱胁迫下,后旗红、荷兰15 号幼苗的根长、克新1 号的茎粗均高于无碱胁迫;而在中、高浓度碱胁迫下,后旗红、荷兰15 号、克新1 号、兴佳2 号以及康尼贝克5 个品种的幼苗根长、茎粗、株高均低于无碱胁迫。这与前人在小麦[19]、大麦[ 2 0 ? 2 1 ]、水稻[ 2 2 ? 2 3 ]、菊芋[ 2 4 ]等作物上的研究结果相同。说明低浓度碱胁迫对有些马铃薯品种生长具有促进作用,而高浓度碱胁迫下,多数马铃薯品种的幼苗根系无法正常生长,这可能是破坏了马铃薯根系内的细胞结构,从而影响生物量的积累。V7 幼苗的根长、茎粗、株高在各胁迫浓度下有所增加,这表明碱胁迫促进了V7 幼苗的生长,这一研究结果与梁晓艳等[16]、修妤等[25]的研究结果不一致,造成此差异的原因可能是作物种类不同以及与本试验所设置的盐碱成分、胁迫浓度有关。
叶绿素作为光合作用的主要色素,其含量可用来衡量植物是否进行正常生长[26]。作物在吸收光能的过程中,叶绿素含量的高低与光合作用密不可分。研究表明,叶绿素含量高有助于促进作物光合作用的进行,提高生物量的积累[27?29]。有学者在苜蓿[30]研究中发现,随着碱性盐浓度提高,pH 值伴随升高,破坏了苜蓿体内叶绿体结构,从而严重阻碍叶绿素形成。本试验中部分马铃薯品种研究结果与上述研究结论相同。即V7、兴佳2 号叶片中叶绿素相对含量随碱胁迫浓度增加呈上升趋势,当胁迫浓度达到40 mmol/L 或50 mmol/L 时,V7、兴佳2 号叶片中叶绿素相对含量在所有供试品种处理中达到最高值;而后旗红、克新1 号、荷兰15 号、康尼贝克4 个品种叶片中叶绿素相对含量呈下降趋势,且均在50 mmol/L 胁迫下低于0 mmol/L。说明在碱胁迫程度不断增加的情况下,V7、兴佳2 号可通过自身的调节机制抵御碱胁迫,叶片中叶绿素的合成不受胁迫影响,可保障叶片进行正常的光合作用;而后旗红、克新1 号、荷兰15 号、康尼贝克4 个品种伴随碱胁迫程度的加剧,幼苗承受压力超过了品种自身可调控的范围,体内叶绿素合成与分解受到影响,进而表现出叶绿素含量降低的现象。
有学者[31?33]在研究草莓、甜菜、小麦等作物时发现,叶片中SPAD 值与含氮量的变化规律相似,本试验研究结果与前人研究结果相一致。对处于生长期的叶片来说,叶片中含氮量和SPAD 值呈正相关,而对于已经皱缩褪绿、衰老的叶片来说,其叶片含氮量与SPAD 值呈负相关[34?35]。本试验研究发现,V7、兴佳2 号叶片含氮量随碱胁迫浓度提高而增加,说明该品种叶片正处于生长期,尚未衰老;其余供试品种叶片中含氮量随碱胁迫加剧呈下降趋势,表明这些品种叶片受到碱害影响较大,抑制了叶片正常生长发育或可能导致其衰老,表现出较差的耐碱性。
当作物受到盐碱胁迫时,地下部根系最先感受到盐害并做出应激反应,因此根系活力也是衡量作物耐盐碱能力的重要指标[36]。前人[37?39]研究表明,低浓度胁迫会提高小麦、水稻、燕麦等幼苗的根系活力,高浓度胁迫下,根系活力显著降低,本试验中部分马铃薯品种研究结果与前人研究结果相一致。20 mmol/L 碱胁迫处理,克新1 号、兴佳2 号、康尼贝克根系活力较0 mmol/L 显著提高,NaHCO3 胁迫呈正效应;而中、高浓度碱胁迫下,克新1 号、兴佳2 号、康尼贝克和荷兰15 号4 个品种根系活力均呈下降趋势,且在50 mmol/L 胁迫下根系活力均低于0 mmol/L。而V7 和后旗红这两个品种则随着碱胁迫浓度增加其根系活力也得到提高,且高浓度胁迫下高于0 mmol/L,这表明不同马铃薯品种其根系对碱胁迫的响应存在差异。碱胁迫条件下,V7 和后旗红二者根系可维持植株正常代谢需求,根系新陈代谢活动强于其他品种。
脯氨酸是作物体内适应逆境胁迫的一种重要渗透调节物质,体内含量越高说明该作物细胞渗透调节能力越强,同时适应逆境胁迫能力也越强。前人[40]研究中发现,叶片中脯氨酸含量随胁迫程度加剧呈显著上升趋势,本研究结果与前人研究结果相一致。6个马铃薯品种幼苗根系中脯氨酸含量均在高碱浓度胁迫下达到峰值。这表明,马铃薯在应对碱胁迫时,可通过积累渗透调节物质来抵御其伤害,从而起到积极作用。
丙二醛含量作为一项生理指标通常被用来评价鉴定细胞膜是否具备完整性,是鉴定体内细胞因碱性盐胁迫造成细胞膜受损程度的重要生理指标[ 4 1 ]。Mandhania 等[41]研究小麦耐盐碱性中发现,随着胁迫浓度增加丙二醛含量显著升高,但抗性好的品种丙二醛含量较低。本研究结果与上述研究相一致。结果表明,随着NaHCO3 胁迫浓度升高,6 个马铃薯品种体内丙二醛含量均呈上升趋势,说明各品种的细胞膜均遭受到不同程度的破坏。值得一提的是,尽管V7、兴佳2 号幼苗中丙二醛含量也伴随胁迫浓度增加而增加,但50 mmol/L 胁迫下,丙二醛含量要低于其他4 个品种,表明V7、兴佳2 号幼苗体内细胞膜相较其他品种来说被破坏的程度轻,耐碱能力强。
SOD、POD、CAT 统称为保护酶系统,是保护植物免受自由氧化伤害最重要的酶系统[42],其作用是清除代谢中产生的H2O2,避免H2O2 积累对细胞的氧化破坏作用。因此SOD、POD、CAT 活性高低与作物耐盐碱能力密切相关。当胁迫超过自身可调控机制时,作物体内 SOD、POD、CAT 的活性降低,使作物生长受到阻碍[42]。有研究表明,随着盐碱胁迫浓度增加,油菜苗期SOD、POD 活性呈先升后降趋势[ 4 3 ];藜麦幼苗叶片中3 种酶活性呈先升后降趋势[42]。本研究发现,随着碱胁迫浓度增加,后旗红、克新1 号、荷兰15 号、康尼贝克4 个马铃薯品种幼苗根系中SOD、POD、CAT 活性均呈先增后降变化趋势,这与前人[42?43]研究结果相一致。表明,马铃薯在响应碱胁迫过程中,SOD、POD、CAT 活性调节具有一定协同作用。当然,作物体内活性氧的清除能力也不是无限的,当胁迫浓度超过一定范围时,体内酶的保护能力也会急剧下降。当碱胁迫浓度增大时,马铃薯植株受到碱害程度加剧,从而导致植株细胞内产生抗氧化酶能力下降,最终抑制其生长。另外,本研究还发现即使在高浓度碱胁迫下,V7 根系中SOD 活性仍高于其他品种,这表明该品种的自我调节和抗氧化能力强于其他供试品种,这一试验结果与前人研究结果[42?43]不一致,导致该结果的原因可能是作物不同、盐碱胁迫成分差异或耐碱能力不同所引起的。
本试验虽以模拟碱胁迫来设计浓度梯度,相比大田试验受外界影响因素较少,但在实际生产中,马铃薯的耐碱性是由多个基因联合代谢组学共同调控表达的结果,其机理尚未进一步明确。今后需从多方面进一步深入研究马铃薯V7 耐碱机理,以便在马铃薯耐碱性应用研究中提供更有力的理论依据。
4 结论
株高、茎粗、含氮量、叶绿素相对含量、脯氨酸含量、POD 活性、CAT 活性可作为马铃薯耐碱能力强弱的评价指标。综上来看,马铃薯品种 V7 耐碱性最强,后旗红、克新1 号、兴佳2 号次之,而荷兰15 号、康尼贝克相对较弱。
参 考 文 献:
[ 1 ]吴运荣, 林宏伟, 莫肖蓉. 植物抗盐分子机制及作物遗传改良耐盐性的研究进展[J]. 植物生理学报, 2014, 50(11): 1621?1629.
Wu Y R, Lin H W, Mo X R. Research progress in the mechanism ofplant salt tolerance and genetic engineering of salt resistant crops[J].Plant Physiology Journal, 2014, 50(11): 1621?1629.
[ 2 ]邓绍云, 邱清华. 中国盐碱土壤修复研究综述[J]. 北方园艺, 2011,(22): 171?174.
Deng S Y, Qiu Q H. Synthetic study on restoration to alkali-salinesoil in China[J]. Northern Horticulture, 2011, (22): 171?174.
[ 3 ]云雪雪, 陈雨生. 国际盐碱地开发动态及其对我国的启示[J]. 国土与自然资源研究, 2020, (1): 84?87.
Yun X X, Chen Y S. International development of saline-alkali land and its enlightenment to China[J]. Territory & Natural ResourcesStudy, 2020, (1): 84?87.
[ 4 ]李聪聪, 宋勇, 孙东泉. 不同品种玉米在盐碱复合胁迫下的幼苗生长差异及其生理机制[J]. 生态学杂志, 2023, 42(10): 2486?2493.
Li C C, Song Y, Sun D Q. Seedling growth difference andphysiological mechanism of different maize cultivars under mixedsaline alkali stress[J]. Chinese Journal of Ecology, 2023, 42(10):2486?2493.
[ 5 ]Correa-Ferreira M L, Viudes E B, De Magalhaes P M, et al. Changesin the composition and structure of cell wall polysaccharides fromArtemisia annua in response to salt stress[J]. Carbohydrate Research,2019, 483: 107753.
[ 6 ]Andrivon D. Potato facing global challenges[J]. Potato Research,2018, 60(3?4): 389?400.
[ 7 ]刘婕, 梅超, 王慧杰, 等. 盐胁迫对4种马铃薯脱毒苗生长及生理特性的影响[J]. 山西农业科学, 2021, 49(5): 550?553.
Liu J, Mei C, Wang H J, et al. Effects of salt stress on the growth andphysiological characteristics of four virus-free potato seedlings[J].Journal of Shanxi Agricultural Science, 2021, 49(5): 550?553.
[ 8 ]刘永强, 马廷蕊, 王方, 等. 马铃薯对盐碱土壤的反应和适应性研究[J]. 土壤通报, 2011, 42(6): 1388?1392.
Liu Y Q, Ma T R, Wang F, et al. Physiological response and adaptivecapacity of potato to saline-alkali soil[J]. Chinese Journal of SoilScience, 2011, 42(6): 1388?1392.
[ 9 ]祁雪, 张丽莉, 石瑛, 崔琳. 盐碱胁迫对马铃薯生理和叶片超微结构的影响[J]. 作物杂志, 2014, (4): 125?129.
Qi X, Zhang L L, Shi Y, Cui L. Influence of salt-alkali stress onphysiological index and leaf ultrastructure of potato[J]. Crops, 2014,(4): 125?129.
[10]孙晓光, 何青云, 李长青, 等. 混合盐胁迫下马铃薯渗透调节物质含量的变化[J]. 中国马铃薯, 2009, 23(3): 129?132.
Sun X G, He Q Y, Li C Q, et al. Changes of osmoregulatorysubstances in potato under mixed salt stress[J]. Chinese PotatoJournal, 2009, 23(3): 129?132.
[11]康益晨. 马铃薯响应碱性盐胁迫的生理及分子机制研究[D]. 甘肃兰州: 甘肃农业大学博士学位论文, 2021.
Kang Y C. Physiological and molecular mechanisms of potatoresponse to alkaline salt stress[D]. Lanzhou, Gansu: PhD Dissertationof Gansu Agricultural University, 2021.
[12]张永成, 田丰. 马铃薯试验研究方法[M]. 北京: 中国农业科学技术出版社, 2007.
Zhang Y C, Tian F. Experimental research methods of potato[M].Beijing: China Agricultural Science and Technology Press, 2007.
[13]Murty A S, Misra P N, Haider. Effect of different salt concentrationson seed germination and seedling development in a few oat cultivars[J]. Indian Agricultural Research Journal, 1984, 18(3): 129?132.
[14]白宝璋, 史安国, 赵景阳, 等. 植物生理学[M]. 北京: 中国农业出版社, 2001.
Bai B Z, Shi A G, Zhao J Y, et al. Plant physiology[M]. Beijing:China Agriculture Press, 2001.
[15]Athah U R, Khan A, Ashraf M. Exogenously applied ascorbic acidalleviates salt-induced oxidative stress in wheat[J]. Environmentaland Experimental Botany, 2008, 63(3): 224?231.
[16]梁晓艳, 顾寅钰, 李萌, 等. 海水胁迫下藜麦根系形态发育及生理响应[J]. 山东农业科学, 2019, 51(11): 28?34.
Liang X Y, Gu Y Y, Li M, et al. Morphological development andphysiological response of quinoa roots under seawater stress[J].Shandong Agricultural Sciences, 2019, 51(11): 28?34.
[17]刘少华, 朱学伸, 闫敏, 王晗. NaCl浸种对盐胁迫下杂交稻幼苗根系生长特性的影响[J]. 西南大学学报(自然科学版), 2020, 42(8):59?65.
Liu S H, Zhu X S, Yan M, Wang H. Effect of NaCl seed soaking onthe growth characteristics of hybrid rice seedling roots under salt stress[J]. Journal of Southwest University (Natural Science Edition), 2020,42(8): 59?65.
[18]Ren P F, Shang L X, Cai Q A, et al. Research progress of plant alkalitolerance and its application prospect in soybean[J]. Soybean Science,2019, 38(6): 977?985.
[19]谷艳芳, 丁圣彦, 李婷, 等. 盐胁迫对冬小麦幼苗干物质分配和生理生态特性的影响[J]. 生态学报, 2009, 29(2): 840?845.
Gu Y F, Ding S Y, Li T, et al. Effects of saline stress on dry matterpartitioning and ecophysiological characteristics of winter wheatseedlings[J]. Acta Ecologica Sinica, 2009, 29(2): 840?845.
[20]王燕, 赵哈林, 赵学勇, 潘成臣. 干旱绿洲农田盐渍化对大麦和苜蓿干物质分配的影响[J]. 中国农业大学学报, 2013, 18(3): 61?67.
Wang Y, Zhao H L, Zhao X Y, Pan C C. Influence of salinization ondry matter partitioning of Medicago sativa and Hordeum vulgare inarid oasis[J]. Journal of China Agricultural University, 2013, 18(3):61?67.
[21]刘志萍, 徐寿军, 张凤英, 等. NaCl胁迫对大麦幼苗生长及光合特性的影响[J]. 大麦与谷类科学, 2014, (4): 1?7.
Liu Z P, Xu S J, Zhang F Y, et al. Effect of NaCl stress on seedlingbiomass and photosynthetic characteristics of barleys[J]. Barley andCereal Sciences, 2014, (4): 1?7.
[22]张素红, 刘忠卓, 李询, 等. 盐胁迫对水稻幼苗期的影响研究: 盐胁迫对水稻干物质的影响[J]. 北方水稻, 2010, 40(2): 21?24.
Zhang S H, Liu Z Z, Li X, et al. Effect of salt stress on rice seedlingstage: Effect of salt stress on dry matter of rice[J]. Northern Rice,2010, 40(2): 21?24.
[23]赵海新, 徐正进, 潘国君, 等. 寒地水稻苗期耐碱研究及形态鉴定指标评价[J]. 黑龙江农业科学, 2014, (11): 21?27.
Zhao H X, Xu Z J, Pan G J, et al. Research on seedling alkaliresistantof rice in cold region and evaluation of its morphologicalidentification indexes[J]. Heilongjiang Agricultural Sciences, 2014,(11): 21?27.
[24]吴成龙, 周春霖, 尹金来, 等. 碱胁迫对不同品种菊芋幼苗生物量分配和可溶性渗透物质量的影响[J]. 中国农业科学, 2008, 41(3):901?909.
Wu C L, Zhou C L, Yin J L, et al. Effects of alkaline stress onbiomass allocation and the contents of soluble osmoticum in differentorgans of two Helianthus tuberosus L. genotypes[J]. ScientiaAgricultura Sinica, 2008, 41(3): 901?909.
[25]修妤, 梁晓艳, 石瑞常, 等. 混合盐碱胁迫对藜麦苗期植株及根系生长特征的影响[J]. 江苏农业科学, 2020, 48(4): 89?94.
Xiu Y, Liang X Y, Shi R C, et al. Effects of complex salt-alkalistresses on the plant and root growth of Chenopodium quinoa[J].Jiangsu Agricultural Sciences, 2020, 48(4): 89?94.
[26]李玉梅, 郭修武, 代汉萍, 姜云天. 盐碱胁迫对牛叠肚幼苗渗透调节物质及叶绿素含量的影响[J]. 经济林研究, 2015, 33(1): 9?16.
Li Y M, Guo X W, Dai H P, Jiang Y T. Effects of salt-saline stress oncontents of osmotic adjustment substances and chlorophyll in Rubuscrataegifolius seedlings[J]. Non-wood Forest Research, 2015, 33(1):9?16.
[27]金稚琴, 李冬林, 丁雨龙, 王磊. 盐胁迫对乌桕幼苗光合特性及叶绿素含量的影响[J]. 南京林业大学学报(自然科学版), 2011, 35(1):29?33.
Jin Z Q, Li D L, Ding Y L, Wang L. Effects of salt stress onphotosynthetic characteristics and chlorophyll content of Sapiumsebiferum seedlings[J]. Journal of Nanjing Forestry University(Natural Sciences Edition), 2011, 35(1): 29?33.
[28]王玉祥, 张博, 王涛. 盐胁迫对苜蓿叶绿素、 甜菜碱含量和细胞膜透性的影响[J]. 草业科学, 2009, 26(3): 53?56.
Wang Y Q, Zhang B, Wang T. Effect of salt stress on the contents ofchlorophyll and betaine and its membrane permeability of Medicagosativa[J]. Pratacultural Science, 2009, 26(3): 53?56.
[29]程丽萍, 刘晋秀, 胡青平. 外源NO对盐胁迫下小麦幼苗叶片丙二醛、 叶绿素及氧化酶的影响[J]. 麦类作物学报, 2013, 33(6):1222?1225.
Cheng L P, Liu J X, Hu Q P. Effects of exogenous nitric oxide on thecontents of MDA and chlorophyll and the activities of oxidases inleaves of wheat seedlings under salt[J]. Journal of Triticeae Crops,2013, 33(6): 1222?1225.
[30]郭鹏, 张万筠, 杨宝灵, 姜健. 盐碱胁迫下紫花苜蓿的生理学及解剖学特征比较[J]. 大连民族学院学报, 2014, 16(5): 486?489.
Guo P, Zhang W Y, Yang B L, Jiang J. Features comparison onphysiology and anatomy in Alfalfa under salt and alkali stress[J].Journal of Dalian Nationalities University, 2014, 16(5): 486?489.
[31]雷泽湘, 艾天成, 李方敏, 董维. 草莓叶片叶绿素含量、含氮量与SPAD值间的关系[J]. 湖北农学院学报, 2001, 21(2): 138?140.
Lei Z X, Ai T C, Li F M, Dong W. The relationships between SPADreadings and the contents of chlorophyll and nitrogen in strawberryleaves[J]. Journal of Hubei Agricultural College, 2001, 21(2): 138?140.
[32]艾天成, 李方敏, 周治安, 等. 作物叶片叶绿素含量与SPAD值相关性研究[J]. 湖北农学院学报, 2000, 20(1): 6?8.
Ai T C, Li F M, Zhou Z A, et al. Relationship between chlorophyllmeter readings (SPAD readings) and chlorophyll content of cropleaves[J]. Journal of Hubei Agricultural College, 2000, 20(1): 6?8.
[33]朱佳, 梁永超, 丁燕芳, 李兆君. 硅对低温胁迫下冬小麦幼苗光合作用及相关生理特性的影响[J]. 中国农业科学, 2006, 39(9): 1780?1788.
Zhu J, Liang Y C, Ding Y F, Li Z J. Effect of silicon on photosynthesisand its related physiological parameters in two winter wheat cultivarsunder cold stress[J]. Scientia Agricultura Sinica, 2006, 39(9): 1780?1788.
[34]张相锋, 杨晓绒, 焦子伟. 植物耐盐性评价研究进展及评价策略[J].生物学杂志, 2018, 35(6): 91?94.
Zhang X F, Yang X R, Jiao Z W. Research progress of salt toleranceevaluation in plants and tolerance evaluation strategy[J]. Journal of Biology, 2018, 35(6): 91?94.
[35]谷俊, 耿贵, 李冬雪, 於丽华. 盐胁迫对植物各营养器官形态结构影响的研究进展[J]. 中国农学通报, 2017, 33(24): 62?67.
Gu J, Geng G, Li D X, Yu L H. Effect of salt stress on plantnutritional organs' morphology and structure: Research progress[J].Chinese Agricultural Science Bulletin, 2017, 33(24): 62?67.
[36]王佺珍, 刘倩, 高娅妮, 柳旭. 植物对盐碱胁迫的响应机制研究进展[J]. 生态学报, 2017, 37(16): 5565?5577.
Wang Q Z, Liu Q, Gao Y N, Liu X. Review on the mechanisms ofthe response to salinity alkalinity stress in plants[J]. Acta EcologicaSinica, 2017, 37(16): 5565?5577.
[37]Amirjani M R. Effect of salinity stress on growth, sugar content,pigments and enzyme activity of rice[J]. International Journal ofBotany, 2011, 7(1): 73?81.
[38]焦德志, 赵泽龙. 盐碱胁迫对植物形态和生理生化影响及植物响应的研究进展[J]. 江苏农业科学, 2019, 47(20): 1?4.
Jiao D Z, Zhao Z L. Research progress on influences of saline-alkalistress on plant morphology, physiology and biochemistry andresponse of plants to saline-alkali stress[J]. Jiangsu AgriculturalSciences, 2019, 47(20): 1?4.
[39] Shevyakova N I, Musatenko L I, Stetsenko L A, et al. Effects of abscisic acid on the contents of polyamines and proline in commonbean plants under salt stress[J]. Russian Journal of Plant Physiology,2013, 60(2): 200?211.
[40]Wang W C, Pang J Y, Zhang F H, et al. Integrated transcriptomicsand metabolomics analysis to characterize alkali stress responses incanola (Brassica napus L. )[J]. Plant Physiology and Biochemistry,2021, 166: 605?620.
[41]Mandhania S, Madam S, Sawhney V. Antioxidant defense mechanismunder salt stress in wheat seedlings[J]. Biologia Plantarum, 2006,50(2): 227?231.
[42]刘文瑜, 杨发荣, 黄杰, 等. NaCl胁迫对藜麦幼苗生长和抗氧化酶活性的影响[J]. 西北植物学报, 2017, 37(9): 1797?1804.
Liu W Y, Yang F R, Huang J, et al. Response of seedling growth andthe activities of antioxidant enzymes of chenopodium quinoato saltstress[J]. Acta Botanica Boreali-Occidentalia Sinica, 2017, 37(9):1797?1804.
[43]杨洋. 不同程度复合盐碱胁迫对油菜苗期生理生化特性的影响[D]. 新疆石河子: 石河子大学硕士学位论文, 2020.
Yang Y. Effects of different levels of combined salt and alkali stresson physiological and biochemical characteristics of rape seedlingstage[D]. Shihezi, Xinjiang: MS Thesis of Shihezi University, 2020.
基金项目:内蒙古自然科学基金项目(2020MS03087);内蒙古农牧业青年创新基金项目(2021QNJJN14)。
猜你喜欢
纺织标准与质量(2022年3期)2022-08-10 09:11:28
少儿科学周刊·儿童版(2021年21期)2021-12-11 01:45:00
安徽农学通报(2021年14期)2021-08-26 02:37:31
粘接(2020年6期)2020-07-09 09:03:26
天津农业科学(2016年12期)2017-01-11 19:50:58
陕西画报(2016年1期)2016-12-01 05:35:30
商(2016年20期)2016-07-04 01:05:25
创新作文(小学版)(2016年31期)2016-03-11 19:08:09
绿色科技(2015年6期)2015-08-05 19:10:52
安徽农学通报(2015年12期)2015-07-09 11:02:20
