
不同山葡萄品种CO2响应模型拟合及评价
2024-06-28 19:36:31潘越王宝庆王季姣马勇李亚兰
中国农业科技导报 2024年4期
潘越 王宝庆 王季姣 马勇 李亚兰
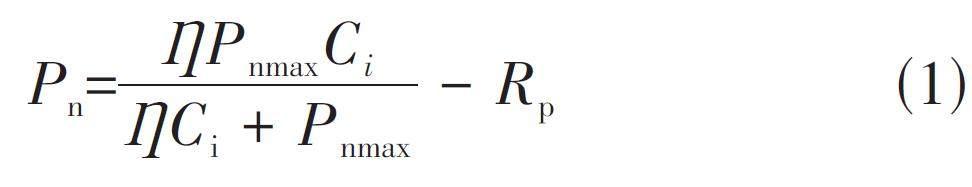
摘要:为探索不同山葡萄品种叶片CO2响应特征差异,以5 a生山葡萄‘北冰红‘北国红‘双红和‘雪兰红为试材,采用Li6400便携式光合仪,测定果实膨大期山葡萄叶片光合-二氧化碳响应曲线(photosynthetic CO2response curve,Pn-Ci)以及胞间CO2 浓度(intercellular CO2 concentration,Ci)、气孔导度(stomatal conductance,Gs)、水分利用率(water use efficiency,WUE)和蒸腾速率(transpiration rate,Tr)等气体交换参数,基于直角双曲线模型、MichaelisMenten模型和直角双曲线修正模型3种模型拟合山葡萄叶片PnCi响应曲线。结果表明,直角双曲线修正模型拟合的山葡萄PnCi响应曲线,其拟合参数与实测值最为接近,可直接计算CO2饱和点(CO2saturation point,CSP)。随大气CO2浓度(atmospheric CO2 concentration,Ca)的增加,4个山葡萄品种Ci呈线性递增趋势;Gs和Tr总体呈先升后降趋势;WUE先降后升,呈“U”型变化趋势。主成分分析提取出2个主成分,累计贡献率达84.613%。综合评价‘雪兰红得分最高,光能转化利用率最高,在低Ca环境下的适应性最佳;‘双红在不同Ca水平下均可保持较高光合效率,排名第2。综上所述,直角双曲线修正模型拟合山葡萄叶片PnCi响应曲线效果最优。
关键词:山葡萄;CO2响应模型;隶属函数;主成分分析
doi:10.13304/j.nykjdb.2022.0915
中图分类号:S663.1 文献标志码:A 文章编号:10080864(2024)04005809
光合作用作为碳循环的重要环节,是植物物质循环及能量流动的基础,植物对环境的敏感程度主要通过光合作用来反映[1]。近年来,由于煤炭、天然气等化石能源的大规模使用,造成温室气体大量排放,导致温室效应及全球气候变暖等一系列环境问题的产生[2-4]。CO2作为主要的温室排放气体,其含量的增加直接影响植物的光合作用[1,5],同时,CO2作为光合作用的基本原料,其含量的变化可直接影响植物光合特性[6]。目前,通过CO2 响应曲线模型建立植物净光合速率(netphotosynthetic rate,Pn)与胞间CO2浓度(intercellularCO2 concentration,Ci )之间的表征关系,并模拟出一系列的CO2 响应参数,如饱和CO2 浓度(CO2saturation point,CSP)、最大净光合速率(maximumnet photosynthetic rate,Pnmax)、CO2 补偿点(CO2compensation point,Γ)、初始羧化效率(initialcarboxylation efficiency,?)、光呼吸速率(rate ofphotorespiration,Rp)等[7]。国内外学者已构建多个光合- 二氧化碳响应模型(photosyntheticCO2response model,PnCi),其中包括直角双曲线模型[8]、直角双曲线修正模型[9]、Michaelis-Menten模型[10]等。据前人研究,经验模型中除二次多项式外,其余模型本质属于单调递增函数,故无法估算Pnmax 和CSP[11];二次多项式虽通过CO2饱和抑制现象计算出CSP,但拟合所得Γ 的参数误差较大[12]。由于不同植物对CO2含量变化的敏感度不一,故不同植物的最佳PnCi响应模型有所差异。因此,针对特定植物,选择与之相适应的PnCi响应模型有助于反映该物种的叶片光合特征。
山葡萄(Vitis amurensis Rupr. )是葡萄科葡萄属中比较抗寒的种之一,其枝蔓能耐-40 ℃低温,根系耐-14~-16 ℃低温,是我国珍贵的抗寒育种资源[1314];同时,山葡萄抗逆性极强[14],在防风固沙、水源涵养、地力维持、环境美化等方面具有显著的生态效益,是典型的生态经济兼用树种。目前,山葡萄的研究多集中于营养成分评价[15]、逆境生理[1617]和葡萄酒理化性质[1819]等方面,尚未有关干旱区山葡萄PnCi响应模型方面的报道。基于此,本研究选取4个南疆地区适应性良好的山葡萄品种[20],通过主成分分析筛选最佳品种和最佳PnCi响应模型,以期为进一步探讨气候变化对山葡萄种群的影响和为山葡萄在南疆地区的推广提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于新疆阿克苏地区温宿县境内的佳木果树学长期科研基地(41°15′N,80°32′E),属温带干旱气候,年均降水量65.4 mm;年均气温10.1 ℃;年均无霜期185 d。
1.2 试验材料
供试山葡萄品种分别为‘北冰红‘ 双红‘ 左优红和‘雪兰红,其中‘北冰红和‘双红为嫁接苗,砧木为‘贝达;‘左优红和‘雪兰红为自根苗。均于2020 年3 月定植,株行距为1.0 m×3.5 m,沿南北行向定植,“厂”字树形,直立叶幕,田间水肥正常管理。
1.3 试验方法
于2022年8月山葡萄转色期,每个品种筛选3株长势一致的植株,于北京时间9:00—11:00,采用美国LI-COR 公司生产的Li-6400 便携式光合仪,选取当年生新梢基部向上的第4~6片成熟、完整的功能叶进行测定。测定Pn和Ci时,大气湿度为45%~60%,光强为1 800 μmol·m-2·s-1,气流速率为152 μmol·s-1,叶面温度为32 ℃,CO2 含量分别为50、100、150、200、300、400、600、800、1 000、1 200 μmol·m-2·s-1,共10个梯度。通过测定结果绘制CO2响应曲线,得出叶片的最大净光合速率(Pnmax)、CO2饱和点(CSP)、CO2补偿点(Γ)和光呼吸速率(Rp),重复3次,以3次重复的平均值作为实测值。测定时,叶室面积设置为25 mm×7 mm,采用CO2钢瓶进行供气,叶室温度设置为27 ℃。
1.4 PnCi响应模型拟合
1.4.1 直角双曲线模型
式中,Pn 为净光合速率;Ci 为胞间CO2浓度;? 为初始羧化效率,是植物光合作用对CO2 响应曲线在Ci =0时的初始斜率;Pnmax为最大净光合速率;Rp为光呼吸速率,在叶片状态稳定后,迅速停止CO2供给,测定叶片光合速率即为Rp。
1.4.2 Michaelis-Menten模型
式中,K为Michaelis-Menten常数。
1.4.3 直角双曲线修正模型
式中,b、c为常数。
饱和胞间CO2浓度(CSP)计算公式如下。
根据CO2饱和点计算Pnmax,公式如下。
将b、c和Rp带入方程,Pnmax=0,求解即为Γ。
1.5 山葡萄综合评价
利用SPSS软件完成方差分析[2122]和主成分分析[23]。数据分析前采用隶属函数法统一数量纲,如指标结果与评价结果呈正相关,采用公式(6);如指标结果与评价结果呈负相关,采用公式(7)。
式中,Uin、Uin 分别为第n 个样本第i 个指标转化后的隶属函数值,Xin指第n 个样本第i 个指标的原始测定值,Ximax和Ximin分别指样本组中第i 个指标的最大值与最小值。
主成分分析:采用标准化后的数据进行主成分分析,提取出对光响应指标有显著影响的主成分,得出各成分的分值Fjn,综合分值Dn以相应公因子的贡献率Ej为权重,根据公式(8)计算。
式中,Dn表示以主成分分析法分析得到的各样品CO2响应参数的综合分值;Fjn表示第n 个样品第j个特征值>1的主成分的分值;m 为特征值>1的主成分数量;Ej为第j 个主成分的贡献率。
2 结果与分析
2.1 Pn对Ci浓度变化响应及拟合
为准确了解山葡萄Pn对Ci的响应,采用3种模型拟合4个山葡萄品种的Pn。由表1和图1可知,直角双曲线修正模型的拟合效果最佳,所得的Pnmax、Γ 和CSP与实测值最接近,拟合指数最接近1;直角双曲线模型和Michaelis-Menten模型拟合‘北冰红和‘雪兰红的Pnmax、Γ 和CSP,其拟合值远高于实测值;拟合‘北国红和‘双红的Pnmax、Γ和CSP,均低于实测值。
4个山葡萄品种Pn-Ci响应曲线的变化情况如图1所示。当Ci为0~600 μmol·mol-1时,4个品种叶片的Pn 迅速增加,增速由高到低表现为‘北国红>‘双红>‘北冰红>‘雪兰红;当Ci 高于Ca时,‘北冰红和‘双红仍有较明显的增加态势,而在相同Ci下,‘北国红和‘雪兰红的Pn显著低于‘北冰红和‘双红。
?、Γ 和CSP 是判定植物对Ci 需求的重要指标,同时也可反映不同Ca环境下植被叶片光合作用能力及不同CO2含量对植被生长发育影响[6,8,24]。由表1可知,4个山葡萄品种的? 表现为‘雪兰红>‘北冰红>‘双红>‘北国红,表明‘北冰红叶片光合作用对CO2的利用最为充分。Γ 和CSP是反映植被利用CO2 效率的重要指标[25]。4个山葡萄品种的Γ 表现为‘雪兰红<‘双红<‘北冰红<‘北国红,说明‘雪兰红利用低水平CO2的能力最强;CSP 表现为‘双红>‘北国红>‘雪兰红>‘北冰红,说明‘双红对高水平CO2的利用率最高。‘双红具有较低的Γ 和较高的CSP,即在高CO2和低CO2环境中均具备较高的光合效率。
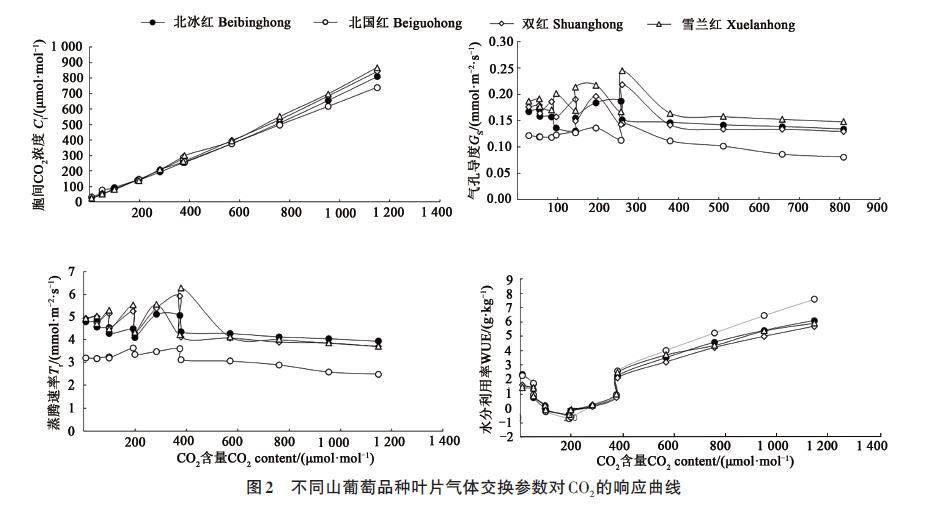
2.2 气体交换参数-Ca响应曲线
不同Ca环境下,4个山葡萄品种叶片的Ci、Gs、Tr和WUE对Ca的响应曲线如图2所示。随着Ca的升高,4个山葡萄品种的Gs和Tr均先升高后降低,最后趋于平缓,其中Gs在Ca为200~300 μmol·mol-1时达到最大;Tr在Ca为300~400 μmol·mol-1时达到最大。WUE 随着Ca 的升高呈先降低后升高趋势,4个山葡萄品种均在Ca为200 μmol·mol-1时最低,在Ca 为200~400 μmol·mol-1 时呈缓慢递增,在Ca 为400~1 200 μmol·mol-1 呈快速递增趋势。叶片Ci 随着Ca 的升高呈快速递增趋势,并在Ca 为1 200 μmol·mol-1时达到最高。
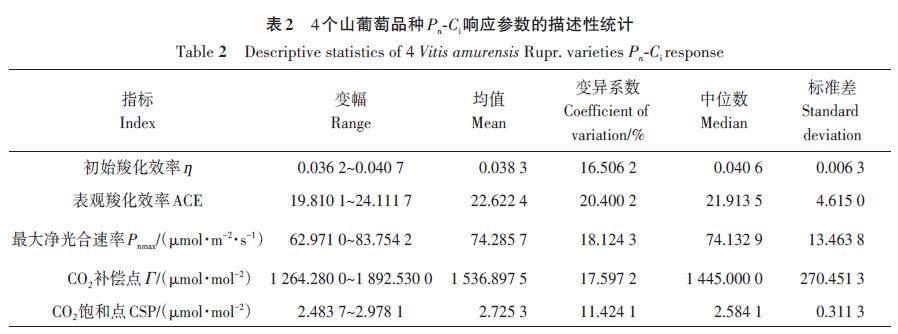
2.3 CO2响应参数描述性分析
供试4个山葡萄品种的5项PnCi响应参数的变异系数均小于30%(表2),说明品种间PnCi响应参数值接近,差异较小。比较均值和中位数发现,除CSP外,其他指标的均值接近中位数,说明这些指标离群点较少,供试山葡萄的各项PnCi响应参数指标测定值均在可接受范围内。
2.4 主成分分析
采用隶属函数法对4个山葡萄品种PnCi响应参数统一数量纲后进行主成分分析,结果如表3所示。从5项响应参数中,提取出2个特征根>1的主成分,累积贡献率达84.613%,即综合了绝大部分的原始信息。因此,在综合评价过程中,采用PC1和PC2进行分析。
旋转后的主成分载荷矩阵经过3 次迭代收敛后,得到旋转主成分载荷矩阵(表4)。PC1综合了? 、Pnmax、Γ 和Rp 的信息,其载荷值分别为0.934 8、0.583 9、0.969 4、-0.783 9,其中? 、Pnmax、Γ 在PC1 上呈正向分布,Rp 呈负向分布,即f1 越大,? 、Pnmax 和Γ 越大,Rp 越低;PC2主要为CSP信息,载荷值为0.957 8,且在PC2上呈正向分布,即PC2越大,CSP越高。
将旋转载荷矩阵各主成分的载荷值除以对应主成分的特征根可得得分矩阵,得分矩阵开平方根与标准化后数据的乘积即为各主成分得分(f1和f2)及综合得分(fZ),关系式如下。
f1=0.579 0X1+0.457 7X2+0.589 7X3-0.222 4X4-0.530 3X5 (9)
f2=-0.469 0X1-0.359 3X2+0.384 3X3+0.814 8X4-0.637 8X5 (10)
不同品种的综合排序如表5所示。对于f1,各品种表现为‘雪兰红>‘双红>‘北冰红>‘北国红,其中,‘雪兰红得分最高,其? 最高,且Γ 最低,即该品种的光能利用效率最高,在低Ca 下适应性最佳;‘双红排名第2,其优势为在不同Ca水平下均可保持较高的光能利用效率;‘北国红得分最低,表明该品种在低Ca 下适应性较差,且光能利用效率较低。对于f2,各品种表现为‘双红>‘北国红>‘雪兰红>‘北冰红,其中,‘双红得分最高,表明该品种在长日照、强光照环境下的耐受力较强,光能利用效率较高。对于fz,各品种表现为‘雪兰红>‘双红>‘北冰红>‘北国红,即‘雪兰红得分最高,表明其在不同Ca水平下的光能利用效率均较高。
3 讨论
当前,植物PnCi响应模型主要为最早被提出的直角双曲线模型,以及在此基础上改进的MichaelisMenten模型、二项式拟合和直角双曲线修正模型,这些模型在不同程度均存在一定缺陷[6,8,26]。直角双曲线模型和MichaelisMenten模型在拟合过程中无法得到CSP,因而无法得出植物在CO2达到饱和状态或过饱和状态时植物的光合作用状况[2728]。二项式拟合的结果相较于其他模型误差较大,尤其在低CO2环境下,误差尤甚。直角双曲线修正模型经过改进,能较好地拟合出植物的CSP,解决了上述模型的弊端,在大量研究中被采用[29-31]。本研究发现,伴随Ca的提高,4个山葡萄品种叶片的Gs 下降,单位面积Tr 下降,整体WUE 提高。3种不同PnCi响应模型对不同山葡萄品种光合特性的拟合效果存在显著差异,直角双曲线模型与MichaelisMenten模型拟合的Pnmax、Γ 和CSP相较于实测值偏大或偏小;直角双曲线修正模型的拟合效果最为理想,各参数的拟合值与实测值最为接近,这可能是由于直角双曲线修正模型在原有模型基础上考虑曲线弯曲程度,曲线拐点更为显著,因此拟合的曲线更符合CO2的响应特征,能够更好地反映4个山葡萄品种叶片光合速率随CO2含量变化的趋势。
伴随全球温室效应加剧,大气中CO2含量对植物的影响逐渐成为研究热点。研究表明,Ca提高会从2个方面影响植物的光合作用,一方面是增加CO2 对Rubisco 酶结合位点的竞争,提高Rubisco酶结合位点的羧化速率[32];另一方面是抑制呼吸作用,叶片Pn 随着Ci 的提高而增加[3334]。本研究发现,4个山葡萄品种叶片的Ci均随着Ca的提高而增加,且在1 200 μmol·mol-1 时达到最大。继续增加Ca 或长期处于高水平Ca 下是否会导致山葡萄叶片Ci降低有待进一步研究。郑凤英等[35]研究发现,Ca 的升高会提高叶片的WUE,降低Gs。Bloom[36]研究认为,Tr会随着Gs的下降而下降。本研究结果与前人研究结果一致,即随着Ca的升高,山葡萄叶片的Gs和Tr协同下降。
吕扬等[37]发现,植物叶片的羧化效率与CO2的利用情况呈显著正相关,即羧化效率越高,叶片CO2利用率越高。尹丽等[38]认为,提高植物的Pnmax和CSP、降低Γ,能有效增加植物光合作用对CO2的利用率,促进有机物的积累。由于原始PnCi响应参数的计量单位未统一,对4 个山葡萄品种PnCi响应进行综合评价时,一些指标会对综合评价结果产生负面影响。因此,本研究采用隶属函数法对数据结果进行转化,能兼顾正、负影响指标对综合评价的效果,使评价结果更为客观合理[39-40]。本研究选择的光响应参数依据试验区独特的区位环境和优选目标而定,?、Pnmax、CSP、Rp越大越好,Γ 越小越好;主成分分析提取到2个特征根>1的主成分,其累计贡献率达84.613%。经综合评价,4个山葡萄品种叶片的光合CO2响应的综合得分表现为‘雪兰红>‘双红>‘北冰红>‘北国红。
参考文献
[1] 肖石红,张卫强,黄芳芳,等. CO2加富对施盐处理下银叶树幼苗叶片光合特性的影响[J].中国水土保持科学,2020,18(5):127-135.
XIAO S H, ZHANG W Q, HUANG F F, et al .. Effect of CO2enrichment on photosynthetic characteristics in the leaves ofHeritiera littoralis seedlings under salinity treatment [J]. Sci.Soil Water Conserv., 2020, 18(5):127-135.
[2] 刘晓聪,董家华,欧英娟.大气中CO2与O3浓度升高对植物光合作用影响研究[J].环境科学与管理,2016, 41(3):152-155.
LIU X C, DONG J H, OU Y J. Research progress on effects ofelevated atmospheric O3 and CO2 on plant photosynthesis [J].Environ. Sci. Manage., 2016, 41(3):152-155.
[3] 郭世博,张方亮,张镇涛,等.全球气候变暖对中国种植制度的可能影响XIV.东北大豆高产稳产区及农业气象灾害分析[J].中国农业科学,2022,55(9):1763-1780.
GUO S B, ZHANG F L, ZHANG Z T, et al .. The Possibleeffects of global warming on cropping systems in China ⅪⅤ.distribution of high-stable-yield zones and agro meteorologicaldisasters of soybean in Northeast China [J]. Sci. Agric. Sin.,2022, 55(9):1763-1780.
[4] IPCC. Climate change 2013: The physical science basis.Working group Ⅰ contribution to the fifth assessment report ofthe intergovernmental panel on climate change, 2013.
[5] JIANG Y L, XU Z Z, ZHOU G S, et al .. Elevated CO2 canmodify the response to a water status gradient in a steppegrass: from cell organelles to photo-synthetic capacity to plantgrowth [J/OL]. BMC Plant Biol., 2016, 16(1):157 [2022-09-20].https://doi.org/10.1186/s12870-016-0846-9.
[6] 叶子飘,康华靖,段世华,等.不同CO2浓度下大豆叶片的光合生理生态特性[J].应用生态学报,2018,29(2):583-591.
YE Z P, KANG H J, DUAN S H, et al .. Photosynthetic physioecologicalcharacteristics in soybean leaves at different CO2concentrations [J]. Chin. J. Appl. Ecol., 2018, 29(2):583-591.
[7] 康华靖,陶月良,权伟,等.植物光合CO2响应模型对光下(暗)呼吸速率拟合的探讨[J].植物生态学报,2014,38(12):1356-1363.
KANG H J, TAO Y L, QUAN W, et al .. Fitting mitochondrialrespiration rates under light by photosynthetic CO2 responsemodels [J]. Chin. J. Plant Ecol., 2014, 38(12):1356-1363.
[8] 叶子飘.光响应模型在超级杂交稻组合-Ⅱ优明86中的应用[J].生态学杂志,2007,26(8):1323-1326.
YE Z P. Application of light-response model in estimatingthe photosynthesis of super-hybrid rice combination ⅡYouming 86 [J]. Chin. J. Ecol., 2007, 26(8):1323-1326.
[9] HARLEY P C, THOMAS R B, REYNOLDS J F, et al ..Modelling photosynthesis of cotton grown in elevated CO2 [J].Plant Cell Environ., 2010,15(3):271-282.
[10] 叶子飘.光合作用对光和CO2响应模型的研究进展[J].植物生态学报,2010,34(6):727-740.
YE Z P. A review on modeling of responses of photosynthesisto light and CO2 [J]. Chin. J. Plant Ecol., 2010, 34(6):727-740.
[11] 赖帅彬,潘新雅,简春霞,等.转苜蓿MsOr 基因烟草光合-光响应和光合-CO2 响应曲线特征研究[J]. 草地学报,2020,28(1):20-30.
LAI S B, PAN X Y, JIAN C X, et al .. Characteristics ofphotosynthetic light response and photosynthetic CO2 responsecurves in transgenic Alfalfa MsOr gene tobacco [J]. ActaAgrestia Sin., 2020, 28(1):20-30.
[12] 王秀伟,毛子军.7个光响应曲线模型对不同植物种的实用性[J].植物研究,2009,29(1):43-48.
WANG X W, MAO Z J. Practicability of 7 light responsivecurve models to different plant species [J]. Bull. Bot. Rese.,2009, 29(1):43-48.
[13] 杨欢,张庆田,刘洪章,等.东北山葡萄区域化及酿酒特性研究概况[J].北方果树,2016(3):1-4.
YANG H, ZHANG Q T, LIU H Z, et al .. The research situationof Northeast Vitis amurensis regionalization and vinificationcharacteristics [J]. Northern Fruits, 2016(3):1-4.
[14] 左倩倩,郑婷,纪薇,等.中国地方葡萄品种分布及收集利用现状[J].中外葡萄与葡萄酒,2019(5):76-80.
ZUO Q Q, ZHANG T, JI W, et al .. The present situation ofdistribution and collection and utilization of grape varieties inChina [J]. Sino-Overseas Grapevine Wine, 2019(5):76-80.
[15] 牛生洋,刘崇怀,刘强,等.葡萄种质果实有机酸组分及其含量特性[J].食品科学,2022,43(12):228-234.
NIU S Y, LIU C H, LIU Q, et al .. Composition and contents oforganic acids in different grape germplasms [J]. Food Sci.,2022, 43(12):228-234..
[16] 徐美隆,乔改霞,仝倩,等.山葡萄砧木对‘黑比诺葡萄耐旱寒的影响研究[J].中外葡萄与葡萄酒,2021(6):66-70.
XU M L, QIAO G X, TONG Q, et al .. Effect of Vitis amurensisrootstock on drought and cold tolerance of ‘Pinot Noirgrapevine [J]. Sino-Overseas Grapevine Wine, 2021(6):66-70.
[17] 袁军伟,李敏敏,贾楠,等.21份葡萄砧木品种资源耐盐性鉴定[J].西北农业学报,2019,28(4):602-606.
YUAN J W, LI M M, JIA N, et al .. Evaluation of salt stresstolerance identification in twenty-one grape rootstock [J]. ActaAgric. Bor-Occid. Sin., 2019, 28(4):602-606.
[18] 金宇宁.东北葡萄产区不同产地及树龄对‘北冰红酒质影响的研究[D].北京:中国农业科学院,2021.
JIN Y N. Study on the effects of different places and tree ageon the wine quality of ‘Beibinghong in northeast grapeproduction area [D]. Beijing: Chinese Academy of AgriculturalSciences, 2021.
[19] 裴辰玉,张薇,李悦,等.CO2浸渍法对北冰红山葡萄酒中成分影响的研究[J].食品工业,2019,40(3):315-319.
PEI C Y, ZHANG W, LI Y, et al .. Study on the effect ofmaceration carbonique composition of Beibinghong grapewine [J]. Food Ind., 2019, 40(3):315-319.
[20] 刘阳阳,潘越,王世伟,等.不同山葡萄品种光响应模型拟合及综合评价[J].中国农业科技导报,2022,24(2):104-114.
LIU Y Y, PAN Y, WANG S W, et al .. Light response modelfitting and comprehensive evaluation for Vitis amurensis [J]. J.Agric. Sci. Technol., 2022, 24(2):104-114.
[21] 公丽艳,孟宪军,刘乃侨,等.基于主成分与聚类分析的苹果加工品质评价[J].农业工程学报,2014,30(13):276-285.
GONG L Y, MENG X J, LIU N Q, et al .. Evaluation of applequality based on principal component and hierarchical clusteranalysis [J]. Trans. Chin. Soc. Agric. Eng., 2014, 30(13):276-285.
[22] 潘越,史彦江,陈淑英,等.喷施叶面肥对平欧杂种榛‘新榛1号嫩枝扦插的影响[J]. 江西农业大学学报,2016,38(5):920-926.
PAN Y, SHI Y J, CHEN S Y, et al .. Effect of foliar fertilizerson the hybrid hazel ‘New hazel one twig cutting [J]. ActaAgirc. Univ. Jiangxiensis, 2016, 38(5):920-926.
[23] 潘越,史开奇,刘珩,等.伊犁不同山葡萄品种的光响应模型筛选及光能利用效率评价[J]. 经济林研究,2022,40(1):178-188.
PAN Y, SHI K Q, LIU H, et al .. Light response model fittingand light use efficiency evaluation for different Vitis amurensisvarieties in Yili region [J]. Non-wood For. Res., 2022, 40(1):178-188.
[24] 李丽霞,刘济明,黄小龙,等.不同氮素条件米槁幼苗光合作用对CO2响应特征[J].东北农业大学学报,2017,48(2):29-36.
LI L X, LIU J M, HUANG X L, et al .. Response characteristicof Cinnamomum migao seedlings photosynthesis to CO2 indifferent conditions of nitrogen [J]. J. Northeast Agric. Univ.,2017, 48(2):29-36.
[25] 陆佩玲,于强,罗毅,等.冬小麦光合作用的光响应曲线的拟合[J].中国农业气象,2001,22(2):13-15.
LU P L, YU Q, LUO Y, et al .. Fitting light response curvesof photosynthesis of winter wheat [J]. Chin. J. Agrometeorol.,2001, 22(2):13-15.
[26] 马兴东,郭晔红,李梅英,等.施氮对干旱区黑果枸杞光合-CO2响应及药效成分的影响[J].西北植物学报,2020,40(7):1209-1218.
MA X D, GUO Y H, LI M Y, et al .. Leaf CO2 response curveand fruit medicinal components of Lycium ruthenicum affectedby nitrogen application in the arid area [J]. Acta Bot. Bor-Occid. Sin., 2020, 40(7):1209-1218.
[27] KYEI-BOAHEN S, LADA R, ASTATKIE T, et al .. Photosyntheticresponse of carrots to varying irradiances [J].photosynthetica, 2003, 41(2):301-305.
[28] LEAKEY A D B, URIBELARREA M, AINSWORTH E A,et al .. Photosynthesis, productivity, and yield of maize are notaffected by open air elevation of CO2 concentration in theabsence of drought [J]. Plant Physiol., 2006, 140(2): 779-790.
[29] 任博,李俊,同小娟,等.太行山南麓栓皮栎和刺槐光合作用-CO2响应模拟[J].应用生态学报,2018,29(1):1-10.
REN B, LI J, TONG X J, et al .. Simulation on photosynthetic-CO2 response of Quercus variabilis and Robinia pseudoacaciain the southern foot of the Taihang mountain, China [J]. Chin.J. Appl. Ecol., 2018, 29(1):1-10.
[30] 郭芳芸,哈蓉,马亚平,等.CO2浓度升高对宁夏枸杞苗木光合特性及生物量分配影响[J]. 西北植物学报,2019,39(2):302-309.
GUO F Y, HA R, MA Y P, et al .. Effects of elevated CO2concentration on photosynthesis characteristics and biomassallocation of Lycium barbarum seedlings [J]. Acta Bot. Bor-Occid. Sin., 2019, 39(2):302-309.
[31] YU Q. Simulation of the stomatal conductance of winter wheatin response to light, temperature and CO2 changes [J]. Ann.Bot., 2004, 93(4):435-441.
[32] 董志新,韩清芳,贾志宽,等.不同苜蓿(Medicago sativa L.)品种光合速率对光和CO2 浓度的响应特征[J]. 生态学报,2007,27(6):2272-2278.
DONG Z X, HAN Q F, JIA Z K, et al .. Photosynthesis rate inresponse to light intensity and CO2 concentration in differentalfalfa varieties [J]. Acta Ecol. Sin., 2007, 27(6):2272-2278.
[33] JAVAID A, MAHMOOD N. Growth, nodulation and yieldresponse of soybean to biofertilizers and organic manures [J].Pakistan J. Bot., 2010, 42(2):863-871.
[34] 李唯.植物生理学[M].北京:高等教育出版社,2012:71-72.
[35] 郑凤英,彭少麟.不同尺度上植物叶气孔导度对升高CO2的响应[J].生态学杂志,2003,22(1):26-30.
ZHENG F Y, PENG S L. Responses of plant stomatalconductance to elevated CO2 at different scales. [J]. Chin. J.Ecol., 2003, 22(1):26-30.
[36] BLOOM A J. Resoured limitation in plants an economicanalogy [J]. Annu. Rev. Ecol. Syst., 1985(16): 541-549.
[37] 吕扬,刘廷玺,闫雪,等.科尔沁沙丘-草甸相间地区黄柳和小叶锦鸡儿光合速率对光照强度和CO2 浓度的响应[J].生态学杂志,2016,35(12):3157 -3164.
LYU Y, LIU T X, YAN X, et al .. Response of photosyntheticrate of Salix gordejevii and Caragana microphylla to lightintensity and CO2 concentration in the dune-meadowtransitional area of Horqin sandy land [J]. Chin. J. Ecol., 2016,35(12):3157-3164.
[38] 尹丽,胡庭兴,刘永安,等.施氮量对麻疯树幼苗生长及叶片光合特性的影响[J].生态学报,2011,31(17):4977-4984.
YIN L, HU T X, LIU Y A, et al .. Effect of nitrogen applicationrate on growth and leaf photosynthetic characteristics ofJatropha curcas L. seedlings [J]. Acta Ecol. Sin., 2011, 31(17):4977-4984.
[39] 董菊兰,麻文俊,王军辉,等.氮素对楸树无性系幼苗的生长影响研究[J].西部林业科学,2012,41(5):31-35.
DONG J L, MA W J, WANG J H, et al .. Growth responses ofCatalpa bungei to nitrogen [J]. J. West China For. Sci., 2012,41(5):31-35.
[40] 冯会丽,吴正保,史彦江,等.基于因子分析的灰枣优良无性系果实品质评价[J].食品科学,2016,37(9):77-81.
FENG H L, WU Z B, SHI Y J, et al .. Fruit quality evaluation ofsuperior clones of Zizyphus jujube cv. Huizao based on factoranalysis [J]. Food Sci., 2016, 37(9):77-81.
(责任编辑:张冬玲)
基金项目:新疆维吾尔自治区重点研发计划项目(2020B02045-2-3)。
猜你喜欢
山东农业科学(2017年2期)2017-03-15 18:49:11
中国房地产·学术版(2016年10期)2016-11-18 19:36:55
大学教育(2016年11期)2016-11-16 20:33:18
中小企业管理与科技·上旬刊(2016年10期)2016-11-15 10:22:56
考试周刊(2016年84期)2016-11-11 23:57:34
现代经济信息(2016年19期)2016-10-20 21:11:15
商业经济研究(2016年14期)2016-09-14 08:58:20
江苏农业科学(2015年8期)2015-09-10 09:46:28
现代农业科技(2015年13期)2015-09-06 06:05:52
电脑知识与技术(2015年10期)2015-05-29 13:36:10
