
褪黑素引发对干旱胁迫下辣椒种子萌发和幼苗生长的影响
2024-06-28 02:58魏茜雅林欣琪梁腊梅秦中维李映志
中国农业科技导报 2024年4期
魏茜雅 林欣琪 梁腊梅 秦中维 李映志

摘要:为提高干旱胁迫下朝天椒种子的萌发势和幼苗生长势,采用1、5、25、50、75、100、125、150 μmol·L-1的褪黑素溶液对朝天椒种子进行引发处理,用10% PEG溶液模拟水分胁迫,分析朝天椒种子萌发和幼苗生长状况及其生理变化。结果表明,与未引发种子相比,100 μmol·L-1褪黑素溶液引发的朝天椒种子在干旱胁迫下发芽势、发芽率、发芽指数和活力指数显著增加425.01%、122.73%、178.55%和796.93%;干旱胁迫下朝天椒幼苗的全长、根长、株高、鲜重、根鲜重、地上鲜重和植株干重显著增加146.70%、221.74%、70.00%、48.88%、121.66%、39.38%和285.71%。生理分析表明,100 μmol·L-1 褪黑素溶液引发的种子的过氧化氢酶(catalase,CAT)活性、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性和可溶性糖(soluble sugar,SS)含量最高,丙二醛(malondialdehyde,MDA)和过氧化氢(hydrogen peroxide,H2O2)含量最小;100 μmol·L-1褪黑素引发的种子在干旱胁迫下萌发后,幼苗的过氧化物酶(peroxidase,POD)活性、CAT活性、APX活性、SS、可溶性蛋白(solubleprotein,SP)和脯氨酸(proline,Pro)含量最大;MDA和活性氧类(reactive oxygen species,ROS)含量最低。以上结果表明,使用100 μmol·L-1的褪黑素溶液对朝天椒种子进行引发处理,能够促进种子在干旱胁迫下的萌发和生长。研究结果可为朝天椒耐旱栽培和育种提供参考。
关键词:干旱胁迫;朝天椒;褪黑素;幼苗生长
doi:10.13304/j.nykjdb.2022.0893
中图分类号:S641.3 文献标志码:A 文章编号:10080864(2024)04004612
辣椒(Capsicum annuum L.) 属于茄科辣椒属一年或多年生草本植物[1]。辣椒是浅根系植物,抵御干旱胁迫能力较弱,如果遭受干旱胁迫,会导致辣椒的产量、品质和光合速率等下降[2]。因此,生产上对耐旱辣椒品种或耐旱栽培措施的需求十分迫切。
种子引发是通过调控种子吸水状态并激活种子生理代谢,使种子质量与植物抗逆能力得到提升的一种种子处理方式[3]。种子引发可以促进生物的调节,在提高幼苗各组织器官的生长和发育等方面表现出重要作用。研究表明,种子引发能明显缓解干旱胁迫带来的压力,减少对乌里胡枝子种子萌发的损伤,保护植株正常生长[4];种子引发能促进玉米植株对营养物质的吸收和转运,促进幼苗生长[5]。
褪黑素(melatonin,MT)是一种多功能的效性分子,可以调节萌发、成熟和衰老等多种生理过程,并提高植株对逆境胁迫的适应能力[6]。研究表明,外源褪黑素处理能有效缓解锑(Sb)对水稻幼苗生长和抗氧化系统的作用[7];褪黑素能与钙离子互作降低高温胁迫对黄瓜幼苗的过氧化伤害[8];褪黑素能提高桃苗的抗旱能力[9]。然而,使用褪黑素对辣椒种子进行引发的研究鲜见报道。本研究通过不同水平的褪黑素对朝天椒种子进行引发处理,评价其对朝天椒种子干旱胁迫下萌发和生长的影响,并分析引发后种子以及种子在干旱胁迫下的生理变化,探究褪黑素引发处理影响抗旱性的可能作用机制。
1 材料与方法
1.1 试验材料及处理
选用朝天椒品种‘茂蔬360为试验材料。挑选饱满、健康的种子分组避光浸没于0(T0)、1(T1)、5(T5)、25(T25)、50(T50)、75(T75)、100(T100)、125(T125)和150 μmol·L-1(T150)的10 mL褪黑素溶液中进行引发,以未引发种子为对照(CK)。12 h后取出种子,用蒸馏水冲洗3~5遍,并避光回干至原始含水量。每个处理120粒种子,重复3次。引发后取上述不同水平褪黑素引发处理和对照的朝天椒种子各1~2 g,进行种子生理指标的测定,试验重复3次。
1.2 种子萌发试验
在玻璃培养皿中铺垫3 层滤纸,用10%PEG[10]完全润湿滤纸,取经引发和未引发的种子每组各60粒均匀铺在培养皿中,然后在恒温25 ℃的培养箱中进行种子萌发试验,重复3次。每天观察并记录发芽种子数,直到确定连续3 d都无种子发芽后停止试验。
1.3 幼苗生长试验
将发芽后的种子均匀地移栽到含PEG(10%)[10]的椰糠基质中继续进行避雨生长,每2 d浇灌1次50 mL 的10% PEG,处理14 d后测定形态指标和生理指标。
1.4 指标测定
1.4.1 种子萌发相关指标的测定
种子的发芽势(germination power,GP)[11]、发芽率(germinationrate,GR)[12]、发芽指数(germination index,GI)[13]和活力指数[14]按照下列公式计算。
GP = 种子发芽达高峰时的发芽数/供试种子数(1)
GR= (n N ) × 100% (2)
GI= Σ(Gt Dt ) (3)
活力指数= GI × S (4)
式中,n 为发芽结束后发芽种子数量,N 为供试种子数;Gt为第t 天的发芽数,Dt为相应的发芽时间;S 为幼苗根的长度。
1.4.2 形态指标测定
将朝天椒幼苗的根系用去离子水清洗干净后吸干表面水分,用毫米尺测定株高和根长,用电子天平称其幼苗地上鲜重和根鲜重[15]。幼苗置于105 ℃烘箱中杀青15 min,再在80 ℃下烘干至恒重,测定幼苗单株干重。每个处理分析3株,重复3次。
1.4.3 生理指标的测定
可溶性糖(solublesugar,SS)含量的测定采用蒽酮比色法[16];丙二醛(malondialdehyde,MDA)含量的测定采用硫代巴比妥酸比色法[16];抗坏血酸过氧化物酶(ascorbateperoxidase,APX)的活性采用Yoshiyμki法测定[17];可溶性蛋白(soluble protein,SP)含量采用考马斯亮蓝G-520 法测定[16];过氧化物酶(peroxidase,POD)、脯氨酸(proline,Pro)、过氧化氢酶(catalase,CAT)、过氧化氢(hydrogen peroxide,H2O2)采用试剂盒(索莱宝)测定;超氧化物歧化酶(superoxide dismutase,SOD)活性使用试剂盒(苏州科铭)测定;超氧阴离子(superoxide anion,O2· -)含量采用羟胺氧化法测定[18];抗坏血酸(ascorbicacid,AsA) 和脱氢抗坏血酸(dehydroascorbate,DHA)含量采用分光光度法[19]测定。
1.5 数据处理
采用Microsoft Excel 2019软件和SPSS 24.0软件进行统计分析和绘图。数据为平均值±标准差(n=3)。
2 结果与分析
2.1 不同水平褪黑素引发处理对干旱胁迫下朝天椒种子萌发的影响
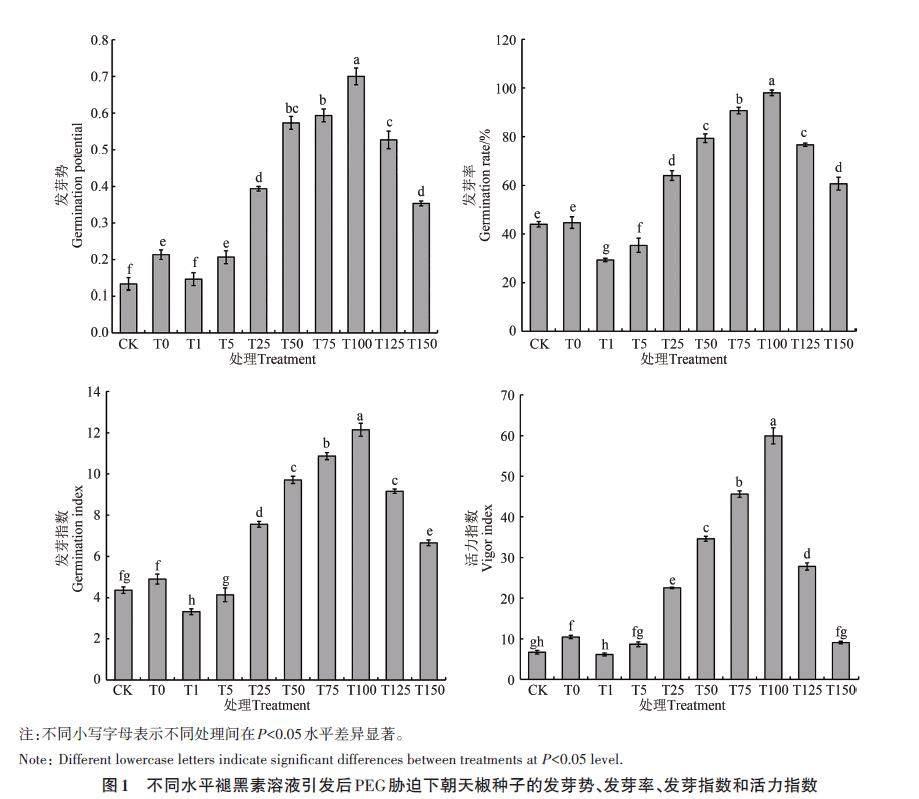
在干旱胁迫(10% PEG)下,不同水平褪黑素引发后朝天椒种子的萌发情况如图1所示,随着引发液水平的增加,朝天椒种子的发芽势、发芽率、发芽指数和活力指数呈抛物线型变化趋势;褪黑素为100 μmol·L-1 时,发芽势、发芽率、发芽指数和活力指数均达到最大值,与其他处理间差异显著(P<0.05);此时与未引发种子相比,发芽势、发芽率、发芽指数和活力指数分别增加425.01%、122.73%、178.55% 和796.93%,表明100 μmol·L-1的褪黑素能够有效缓解干旱胁迫对朝天椒种子萌发的抑制作用。
2.2 不同水平褪黑素引发处理对干旱胁迫下朝天椒形态指标的影响
干旱(10% PEG)胁迫下,不同水平褪黑素引发处理后朝天椒幼苗的全长、根长、株高和根冠比如图2所示,随着引发液水平的增加,朝天椒幼苗的全长、根长、株高和根冠比均呈抛物线型变化趋势,褪黑素水平为100 μmol·L-1时,朝天椒植株的全长、根长、株高和根冠比均达到最大值,与未引发种子相比,分别显著增加146.70%、221.74%、70.00% 和60.92%(P<0.05),表明100 μmol·L-1 的褪黑素溶液对于提高干旱胁迫下幼苗的株高和根长有显著作用。
干旱(10% PEG)胁迫下,不同水平褪黑素引发处理后朝天椒幼苗的鲜重、根鲜重、地上鲜重和植株干重如图3所示,随着褪黑素水平的增加,幼苗鲜重、根鲜重、地上鲜重和植株干重均呈抛物线型变化趋势;褪黑素为100 μmol·L-1时,朝天椒幼苗的鲜重、根鲜重、地上鲜重和植株干重达到最大值,此时与未引发种子相比,分别显著增加48.88%、121.66%、39.38%和285.71%(P<0.05)。
2.3 不同水平褪黑素引发处理对朝天椒种子生理的影响
2.3.1 对朝天椒种子MDA 和活性氧含量的影响
如图4所示,经不同水平褪黑素溶液引发处理后,朝天椒种子的MDA和H2O2含量随着引发液含量水平的增加呈先降低后上升的趋势。低水平褪黑素引发处理提高了朝天椒种子的活性氧(reactive oxygen species,ROS)含量,褪黑素含量为100 μmol·L-1时,引发后种子的MDA和H2O2含量均为最小值,与未引发相比分别下降56.01% 和21.89%,差异显著(P<0.05);100 μmol·L-1褪黑素引发处理后的朝天椒种子的超氧阴离子含量与未引发相比无显著差异。
2.3.2 对朝天椒种子SS、SP 和Pro 含量的影响
如图5所示,经不同水平褪黑素引发处理后朝天椒种子的SS 和SP 含量随着引发液水平的增加呈抛物线型变化趋势。引发后种子的SS 含量在100 μmol·L-1 时达到最大值,与未引发相比增加了283.35%;引发后种子的SP 含量在75 μmol·L-1 时达到最大值,与未引发相比增加了109.72%,差异显著(P<0.05);引发后种子的Pro含量在75和100 μmol·L-1时与未引发相比无显著差异。
2.3.3 对朝天椒种子POD 和CAT 活性的影响
如图6所示,经不同水平褪黑素引发处理后朝天椒种子的POD和CAT活性均有所提高,引发后种子的POD活性在75 μmol·L-1时达到最大值,与未引发相比增加346.18%;引发后种子的CAT活性在100 μmol·L-1 时达到最大值,与未引发相比增加143.80%,差异显著(P<0.05)。
2.3.4 对朝天椒种子APX活性、AsA和DHA含量的影响
经不同水平褪黑素引发处理后朝天椒种子的APX活性、AsA、DHA含量和AsA/DHA 均随着引发液水平的增加呈抛物线型变化趋势(图7),褪黑素含量为100 μmol·L-1时,引发后种子的APX活性、AsA和DHA含量为最大值,与未引发相比增加196.97%、161.54%和91.54%;AsA/DHA在褪黑素含量为50 μmol·L-1时达到最大值,与未引发相比显著增加49.27%(P<0.05)。
2.4 褪黑素种子引发处理对干旱胁迫下朝天椒幼苗生理的影响
2.4.1 对干旱胁迫下朝天椒幼苗MDA和ROS含量的影响
如图8所示,干旱胁迫下不同水平褪黑素引发处理后朝天椒幼苗的MDA、H2O2 和超氧阴离子含量均在100 μmol·L-1时最小,与未引发相比分别降低78.70%、80.89%和46.25%,差异显著(P<0.05)。由此可以看出,褪黑素引发可作为缓解干旱胁迫对辣椒幼苗生长胁迫的有效途径之一。
2.4.2 对干旱胁迫下朝天椒幼苗SS、SP和Pro含量的影响
如图9所示,褪黑素含量显著影响干旱胁迫下幼苗的SS、SP和Pro含量,随着褪黑素溶液含量水平的升高,幼苗体内的SS、SP和Pro含量呈先增大后减小的趋势,并都在100 μmol·L-1 时的值最大,与未引发相比分别增加81.07%、268.03%和661.93%,差异显著(P<0.05)。
2.4.3 对干旱胁迫下朝天椒幼苗SOD、POD 和CAT 活性的影响
脂膜过氧化酶活性的高低显著影响着植物的抗逆能力。如图10所示,干旱胁迫下不同水平褪黑素引发处理后幼苗的POD和CAT活性明显增大并都在100 μmol·L-1时的值最大,与未引发相比分别增加551.52%和255.52%;SOD活性在75 μmol·L-1时的值最大,与未引发相比增加240.46%,差异显著(P<0.05)。
2.4.4 对干旱胁迫下朝天椒幼苗APX活性、AsA和DHA 含量的影响
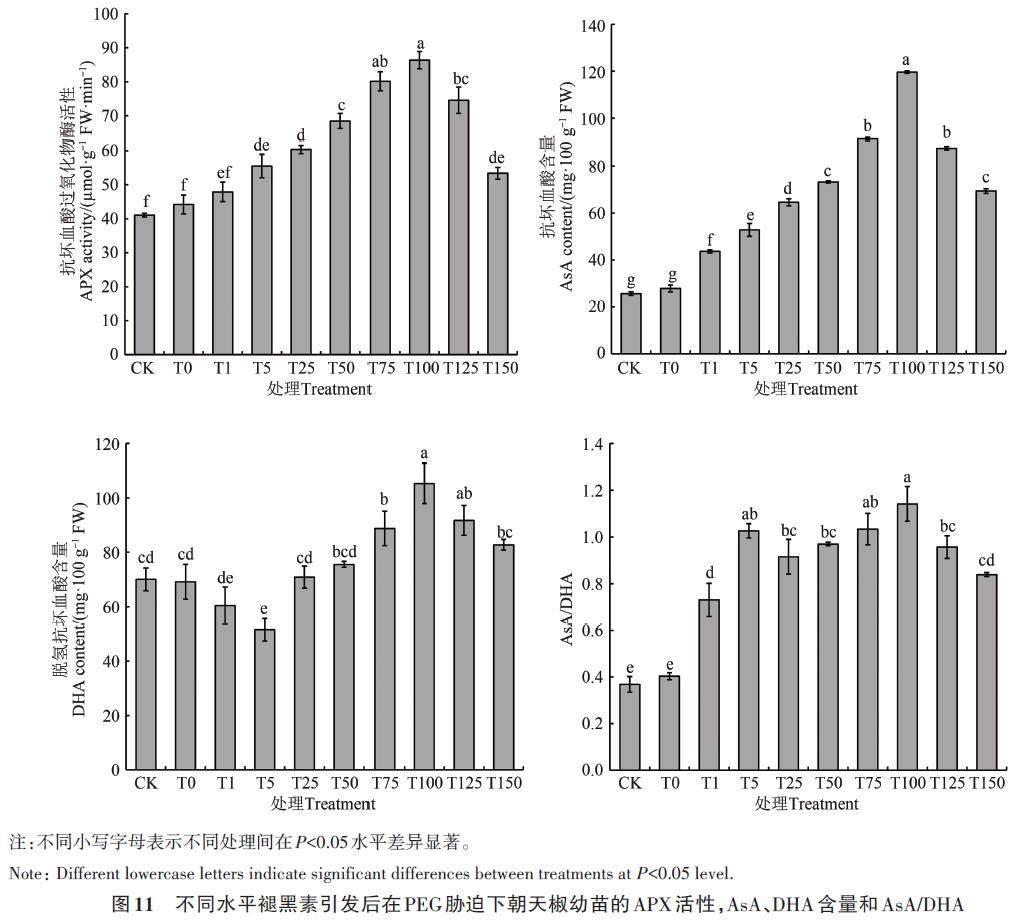
如图11所示,经不同水平褪黑素引发处理后干旱胁迫下朝天椒幼苗的APX 活性、AsA、DHA 含量和AsA/DHA 均在100 μmol·L-1 时达到最大值,与未引发相比分别增加110.26%、366.54%、50.27%和210.19%,差异显著(P<0.05)。
3 讨论
3.1 褪黑素引发处理对干旱胁迫下朝天椒种子萌发的影响机制
本研究表明,在褪黑素引发朝天椒种子处理中,100 μmol·L-1 的褪黑素能明显抑制种子MDA的增加和H2O2 的积累,保护膜结构不受损伤;有效提高SS和SP含量,维持细胞渗透压并调节植物生理过程;显著促进种子内POD、CAT 和APX活性,保持种子的活力;并提高种子AsA、DHA含量和AsA/DHA,保护细胞处于良好的生境。这与褪黑素能促进玉米种子抗氧化系统,提高新陈代谢能力的结果一致[20],说明适宜水平的褪黑素可以有效清除种子内的活性氧,维持活性氧代谢平衡,从而降低活性氧对细胞膜系统造成的氧化损。而100 μmol·L-1的褪黑素引发处理种子中超氧阴离子含量与未引发相比无显著差异,且与其他研究不一致,可能是因为朝天椒种子较其他植物清除超氧阴离子能力较弱。
研究发现,干旱胁迫在植株生长过程中严重影响种子萌发和植株发育[21]。本研究表明,100 μmol·L-1的褪黑素种子引发处理可促进干旱胁迫下朝天椒种子的萌发,其发芽率、发芽势、发芽指数和活力指数均显著高于未引发处理的种子,这与苗含笑等[22]发现褪黑素引发小麦种子可提高小麦种子的萌发,并增加单穗穗重,进而增加产量的结果一致。100 μmol·L-1的褪黑素种子引发处理能促进干旱胁迫下朝天椒种子萌发的原因可能是褪黑素能调节种子的生理状态,促使种子生命活动强度增大。适宜水平的褪黑素可以有效缓解干旱胁迫对种子萌发的抑制作用,显著提高种子的萌发能力。
3.2 褪黑素种子引发处理提高朝天椒幼苗干旱胁迫耐性的机制
干旱胁迫可以通过改变植株的生理和生化机制对植株造成不利影响。例如,通过改变植株的形态学和新陈代谢抑制气孔开启并减少叶光合作用[23]。本研究表明,100 μmol·L-1褪黑素引发处理后,干旱胁迫下幼苗的全长、根长、株高、根冠比、鲜重和干重显著增加,说明100 μmol·L-1 褪黑素引发处理对朝天椒地上和地下生物量都有促进作用。这与李爱等[24]发现外源褪黑素能提高干旱胁迫下紫苏幼苗的植株全长、根长和生物量结果一致。说明适宜水平的褪黑素有利于辣椒幼苗对水分的吸收,进而缓解干旱胁迫对幼苗的损伤,提高耐旱性。
渗透调节是一种重要的调节机制,可以保证植物在各种胁迫环境中能够承受的渗透压力[25]。干旱胁迫导致的水分损失进一步限制了营养物质的运输,从而降低植株新陈代谢,产生更多的活性氧,导致脂膜受到损伤[26]。本研究表明,干旱胁迫下,100 μmol·L-1 褪黑素引发处理后可使幼苗渗透调节物质显著增加,且朝天椒幼苗中的抗氧化酶活性比未引发的幼苗有明显上升趋势,说明100 μmol·L-1褪黑素具有修复干旱胁迫下辣椒幼苗细胞膜结构并维持细胞内环境的稳定性的作用;能缓解水分缺乏给植株带来的渗透压力,降低胁迫带来的负面压力;能提高幼苗在干旱胁迫中的自我防御体系,维持膜结构及其稳定性,且朝天椒植株抗氧化能力的提高显著降低了幼苗MDA和ROS含量,抑制干旱胁迫对朝天椒幼苗造成的脂膜过氧化反应,从而增强了辣椒的耐旱性。孟祥萍[27]研究表明,褪黑素引发能增加冬小麦在水分胁迫下的酶活性,抑制活性氧的产生。
还原型抗坏血酸(AsA)作为最主要的非酶抗氧化剂可以通过APX 的催化作用提高植物的氧化能力,减少非生物胁迫诱导的氧化胁迫[7],AsA/DHA 可作为调节抗氧化机制的信号[28]。本研究表明,干旱胁迫下,100 μmol·L-1褪黑素引发处理可使幼苗APX 活性、AsA、DHA 含量和AsA/DHA显著升高,说明100 μmol·L-1褪黑素可以通过提高APX活性,催化分解AsA来保持良好的氧化还原状态,并缓解干旱胁迫对幼苗的损伤;且AsA能与其他抗氧化剂共同组成抗氧化系统,保护幼苗免受有氧代谢和干旱胁迫造成的氧化损害。徐向东等[29]研究发现,叶面喷施褪黑素能提高黄瓜幼苗的抗坏血酸代谢系统。
综上所述,100 μmol·L-1 的褪黑素引发处理能够促进朝天椒种子的萌发和幼苗根系的生长;降低幼苗MDA和ROS含量的积累;提高幼苗SS、SP、Pro和AsA含量,并增强APX、SOD、POD、CAT活性,从而减缓干旱胁迫对朝天椒种子和幼苗的伤害,提高耐旱性。
参 考 文 献
[1] 牛彩霞, 郁继华, 张韵, 等. 钠盐对辣椒种子萌发和幼苗生长的影响[J]. 甘肃农业大学学报, 2006, 41(1): 34-38.
NIU C X, YU J H, ZHANG Y, et al .. Effects of salt stress onseed germination and seedling growth of pepper [J]. J. GansuAgric. Univ., 2006,41(1): 34-38.
[2] 宋志荣. 干旱胁迫对辣椒生理机制的影响[J]. 西南农业学报, 2003, 16(2): 53-55.
SONG Z R. Effects of drought stress on physiologicalmechanism of hot pepper [J]. Southwest China J. Agric. Sci.,2003, 16(2): 53-55.
[3] 石晓琪,米素娟,钟天航,等.种子引发提高草类植物抗旱性的表现及机理[J].草地学报,2022,30(10):2692-2700.
SHI X Q, MI S J, ZHONG T H, et al .. Performance andmechanism of seed priming to improve drought tolerance inherbage [J]. Acta Agrestia Sin., 2022,30(10):2692-2700.
[4] 董秋丽, 王聪聪, 郑川, 等. 褪黑素引发对达乌里胡枝子种子干旱萌发的影响[J]. 中国草地学报, 2022, 44(7): 114-120.
DONG Q L, WANG C C, ZHENG C, et al .. Effects of melatoninpriming on germination of Lespedeza davurica seeds underdrought stress [J]. Chin. J. Grassland, 2022, 44 (7): 114-120.
[5] 杜锦, 肖萌, 郝娜娜, 等. 不同药剂引发对干旱胁迫下玉米种子萌发及幼苗生长的影响 [J]. 种子, 2014, 33(11): 43-46.
DU J, XIAO M, HAO N N, et al .. Effect of seed priming withdifferent agents on seed germination and seedling growth inmaize (Zea mays L.) under water deficit stress [J]. Seed, 2014,33 (11): 43-46.
[6] 肖珍珍, 隋晓青, 石国庆, 等. 外源褪黑素不同浸种浓度和时长对干旱胁迫下无芒雀麦种子萌发的影响 [J]. 草地学报, 2022,30(3):655-660.
XIAO Z Z, SUI X Q, SHI G Q, et al .. Effects of differentconcentrations and immersion durations of exogenousmelatonin on seed germination of bromus inermis underdrought stress [J]. Acta Agrestia Sin., 2022, 30(3):655-660.
[7] 储玉檀,李颜,黄益宗,等.外源褪黑素对锑胁迫下水稻幼苗生长和抗氧化系统的影响[J]. 环境科学,2023,44(4):2356-2364.
CHU Y T, LI Y, HUANG Y Z, et al .. Effects of exogenousmelatonin treatment on the growth and antioxidant system ofrice seedlings under antimony stress [J]. Chin. J. Environ. Sci.,2023,44(4):2356-2364.
[8] 徐晨潇,张晓宇,刘超越,等.外源褪黑素与钙离子互作对高温胁迫下黄瓜幼苗过氧化伤害的缓解效应[J]. 应用生态学报, 2022,33(10):2725-2735.
XU C X, ZHANG X Y, LIU C Y, et al .. Alleviating effect ofexogenous melatonin and calcium on the peroxidation damagesof cucumber under high temperature stress [J]. Chin. J. Appl.Ecol., 2022,33(10):2725-2735.
[9] 古咸彬, 陆玲鸿, 宋根华, 等. 外源褪黑素预处理对干旱胁迫下桃苗生长的缓解效应[J]. 植物生理学报, 2022, 58(2):309-318.
GU X B, LU L H, SONG G H, et al .. The mitigative effect ofexogenous melatonin pretreatment on peach seedling growthunder drought stress [J]. Plant Physiol. J., 2022,58 (2): 309-318.
[10] 张爱民, 蓬桂华, 王永平, 等. PEG模拟干旱胁迫对辣椒种子萌发的影响[J]. 辣椒杂志, 2015, 13(3): 36-38,41.
ZHANG A M, PENG G H, WANG Y P, et al .. Effects of PEG-6000 on seed germination in pepper [J]. J. China Capsicum,2015, 13 (3): 36-38,41.
[11] 徐蓉蓉, 高静, 任炳浩, 等. 温度对干旱、盐胁迫下两种黄芪属种子萌发和幼苗生长的影响[J]. 生态学杂志, 2020,39(9): 2930-2943.
XU R R, GAO J, REN B H, et al .. Effects of temperature onseed germination and seedling growth of two Astragalus speciesunder drought and salt stresses [J]. Chin. J. Ecol., 2020, 39 (9):2930-2943.
[12] 谢浩然, 杨月娟, 朱刚, 等. 温度及赤霉素预处理对枫香种子萌发的影响[J]. 现代园艺, 2021, 44(18): 10-13.
XIE H R, YANG Y J, ZHU G, et al.. The effects of temperatureand gibberellin pretreatment on the germination of maple seeds[J] Mod. Hortic., 2021, 44 (18): 10-13.
[13] 王辉, 马向丽, 另如贵, 等. 舟叶橐吾浸提液对三种牧草种子萌发及幼苗生长的影响[J]. 草地学报, 2022, 30(1): 93-99.
WANG H, MA X L, LING R G, et al .. Allelopathic effects ofLigularia cymbulifera extracts on seed germination and seedlinggrowth of three forages [J]. Acta Agrestia Sin., 2022, 30(1):93-99.
[14] 徐劼, 胡博华, 戈涛, 等. 镉胁迫对生菜种子萌发及幼苗生理特性的影响[J]. 湖北农业科学, 2014, 53(20): 4892-4896.
XU J, HU B H, GE T, et al .. Effects of cadmium stress on seedgermination and physiological characteristics of lettuceseedling [J]. Hubei Agric. Sci., 2014, 53 (20): 4892-4896.
[15] 刘金海, 蒋金娟, 罗富成, 等. 外源二聚丙三醇对非洲狗尾草种子萌发及幼苗生长的影响[J]. 草地学报, 2022, 30(4):950-956.
LIU J H, JIANG J J, LUO F C, et al .. Effects of exogenousdiglycerol treatment on seed germination and seedling growthof Setaria sphacelata‘ Narok [J]. Acta Agrestia Sin., 2022, 30(4):950-956.
[16] 刘家尧, 刘新. 植物生理学实验教程[M]. 北京:高等教育出版社, 2010:1-110.
[17] NAKANO Y, ASADA K. Hydrogen peroxide is scavenged byascorbate-specific peroxidase in spinach chloroplasts [J]. PlantCell Physiol., 1981, 22(5): 867-880.
[18] 王爱国, 罗广华. 植物的超氧物自由基与羟胺反应的定量关系[J]. 植物生理学通讯, 1990(6): 55-57.
WANG A G, LUO G H. Quantitative relation between thereaction of hydroxylamine and superoxide anion radicals inplants [J]. Plant Physiol. Comm., 1990(6): 55-57.
[19] 王俊力, 王岩, 赵天宏, 等. 臭氧胁迫对大豆叶片抗坏血酸-谷胱甘肽循环的影响[J]. 生态学报, 2011, 31(8): 2068-2075.
WANG J L, WANG Y, ZHAO T H, et al .. Effects of ozone onAsA-GSH cycle in soybean leaves [J]. Acta Ecol. Sin., 2011,31(8): 2068-2075.
[20] CAO Q J, LI G, CUI Z G, et al .. Seed priming with melatoninimproves the seed germination of waxy maize under chillingstress via promoting the antioxidant system and starchmetabolism [J/OL]. Sci. Rep., 2019, 9(1): 15044 [2022-09-23].https://doi.org/10.1038/s41598-019-51122-y.
[21] 边才苗, 金则新, 张俊会, 等. 云锦杜鹃种子萌发及对干旱胁迫的响应[J]. 植物研究, 2006,26(6):718-721.
BIAN C M, JIN Z X, ZHANG J H, et al .. Response of seedgermination of Rhododendron fortunei to drought stress [J].Bull. Botan. Res., 2006,26(6):718-721.
[22] 苗含笑,李东晓,王久红,等.褪黑素对干旱胁迫下小麦生长发育和产量的影响[J].干旱地区农业研究,2020,38(5):161-167,191.
MIAO H X, LI D X, WANG J H, et al .. Effects of melatonin onthe growth and yield of wheat under drought condition [J].Agric. Res. Arid Areas, 2020,38(5):161-167,191.
[23] 郝田, 范宁丽, 于景金. CO2浓度升高影响植物生长发育的研究进展[J]. 江苏农业科学, 2020, 48(21): 52-56.
HAO T, FAN N L, YU J J. Research progress on effects ofelevated CO2 concentration on plant growth and development[J]. Jiangsu Agric. Sci., 2020, 48 (21): 52-56.
[24] 李爱,李林,孙汪亮,等.外源褪黑素对干旱胁迫下紫苏种子萌发的影响[J].北方园艺,2018(5):140-144.
LI A, LI L, SUN W L, et al .. The effect of exogenous melatoninon the germination of perilla seeds under drought stress [J].Northern Hortic., 2018 (5): 140-144.
[25] 朱亚, 李松, 赵永平, 等. 干旱胁迫对不同品种决明幼苗生理生化特性的影响[J]. 陕西农业科学, 2022, 68(7): 1-5.
ZHU Y, LI S, ZHAO Y P, et al .. Effect of drought stress on physiological and biochemical characteristics of seedlings ofdifferent varieties of Cassia obtusifolia L. [J]. Shaanxi J. Agric.Sci., 2022, 68(7): 1-5.
[26] ZHANG W, YU X, LI M, et al .. Silicon promotes growth androot yield of Glycyrrhiza uralensis under salt and droughtstresses through enhancing osmotic adjustment and regulatingantioxidant metabolism [J]. Crop Protect., 2018, 107: 1-11.
[27] 孟祥萍. 褪黑素引发种子对冬小麦水分胁迫下生长及生理的影响[D]. 杨凌:西北农林科技大学, 2016.
MENG X P. Effects melatonin priming seedlings treatment ongrowth and physiolgy of wheat under water stress [D].Yangling: Northwest A&F University, 2016.
[28] BASHRI G, MOHAN S. Exogenous IAA differentially affectsgrowth, oxidative stress and antioxidants system in Cd stressedTrigonella foenum-graecum L. seedlings: toxicity alleviation byup-regulation of ascorbate-glutathione cycle [J]. Ecotoxicol.Environ. safe., 2016,132: 329-338.
[29] 徐向东, 孙艳, 郭晓芹, 等. 褪黑素对高温胁迫下黄瓜幼苗抗坏血酸代谢系统的影响[J]. 应用生态学报, 2010,21(10):2580-2586.
XU X D, SUN Y, GUO X Q, et al .. Effects of exogenousmelatonin on ascorbate metabolism system in cucumberseedlings under high temperature stress [J]. Acta Ecol. Sin.,2010,21(10):2580-2586.
(责任编辑:胡立霞)
基金项目:广东海洋大学创新强校项目(GDOU2013050217,GDOU2016050256)。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
安徽农学通报(2016年21期)2016-12-22
山东农业科学(2016年11期)2016-12-17
现代园艺(2016年2期)2016-03-15
中国蔬菜(2015年9期)2015-12-21
小说月刊(2014年1期)2014-04-23
