
干旱胁迫下氮素对猴樟幼苗生长及光合特性的影响
2024-06-01 00:35:51唐佳乐段海波王静
湖北农业科学 2024年4期
唐佳乐 段海波 王静


摘要:为研究猴樟(Cinnamomum bodinieri)幼苗对干旱胁迫的响应及外源添加氮素对其抗旱性调控的效应,以1年生猴樟幼苗为试验材料,以霍格兰营养液为氮源,设置缺氮(N0)、施氮(N1)2个氮素水平,并对其进行干旱处理,分别于停止浇水后0 d(正常水分,W0)、3 d(轻度干旱,W1)、5 d(中度干旱,W2)、8 d(重度干旱,W3)进行生长指标、叶绿素含量和光合气体交换参数的测定。结果表明,干旱胁迫会抑制猴樟幼苗的生长及生物量的积累,降低叶片叶绿素含量,诱导气孔关闭,降低胞间CO2浓度,抑制叶片的净光合速率。外源添加氮素能够促进猴樟幼苗的生长,且主要体现为对地上部分生长的促进,并能够提高幼苗叶片叶绿素含量,显著提高干旱胁迫下猴樟幼苗的净光合速率。
关键词: 干旱胁迫; 猴樟(Cinnamomum bodinieri)幼苗; 氮素; 生长; 光合特性
中图分类号:S688.4 文献标识码:A
文章编号:0439-8114(2024)04-0096-05
Effects of nitrogen on growth and photosynthetic characteristics of Cinnamomum bodinieri seedlings under drought stress
Abstract: To investigate the response to drought stress and the effect of exogenous nitrogen addition on drought resistance, 1-year-old Cinnamomum bodinieri seedlings were used as experimental materials, and the Hoagland nutrient solution was used as the nitrogen source. Two nitrogen levels, nitrogen deficiency (N0) and nitrogen application (N1) were set, and drought treatment was applied. The growth indexes, chlorophyll content and photosynthetic gas exchange parameters were measured at 0 d(normal water, W0), 3 d(mild drought, W1), 5 d(moderate drought, W2) and 8 d(severe drought, W3) after stopping watering, respectively. The results showed that drought stress could inhibit the growth and biomass accumulation of Cinnamomum bodinieri seedlings, reduce leaf chlorophyll content, induce stomatal closure, reduce intercellular carbon dioxide concentration, and inhibit the net photosynthetic rate of leaves. Exogenous nitrogen addition could promote the growth of Cinnamomum bodinieri seedlings, mainly reflected in the growth of the above-ground part, and could increase the chlorophyll content of the seedlings, and significantly increase the net photosynthetic rate of the seedlings under drought stress.
Key words: drought stress; Cinnamomum bodinieri seedling; nitrogen;growth; photosynthetic characteristics
随着人们对高质量生活水平的需求显著增加,城市绿化面积不断增加,极大改善了人居环境,但也给城市绿化养护管理带来了一系列难题,如在夏季高溫期间城市绿地灌溉用水量大幅度增加,甚至高达城市用水量的30%[1],造成了城市水资源的严重短缺,城市绿化的养护管理成本显著提高,因此创建节水型园林迫在眉睫。筛选及培育抗旱性园林植物是构建节水型园林的重要发展方向,但通过外部因素提高植物抗旱能力也是一条见效时间快、成本低的切实可行路径[2]。
水分和氮素是植物生长过程中所必需的元素及物质,有研究表明水分与氮素之间存在显著的耦合效应,干旱胁迫通过影响植物的形态建成、水分吸收及运输、光合作用等众多重要生理代谢过程抑制植物的生长发育[3-6],其中光合作用过程对干旱胁迫极为敏感[7],有众多研究表明干旱胁迫通过诱导气孔关闭、增加CO2扩散阻力、抑制光合酶活性降低净光合速率[8]。外源添加氮素可以促进干旱胁迫下根系的生长,并增加干旱胁迫下植物叶绿素含量及提高净光合速率,进而提高植物的抗旱性[8-10],同时,在城市园林绿化养护管理中外源施加氮肥简便易行,成本较低,且可以大幅度减少对城市水资源的利用。
猴樟(Cinnamomum bodinieri)是樟科樟属的一种大型常绿乔木,主要分布在湘鄂及云贵川地区[11],具有树形优美、生长速度快、抗病虫害能力强等特性[12],是优良的行道树和庭荫树树种。有研究表明猴樟具有一定的抗旱能力[13],但面对日益突出的城市水资源缺乏问题,急需进一步通过外部手段提高植物的抗旱能力,但关于氮素对猴樟抗旱能力的影响鲜见详细报道。因此,本研究以猴樟一年生幼苗为研究对象,研究干旱胁迫对猴樟幼苗生长及光合特性的影响及外源氮素对其抗旱性调控的效应,以期为猴樟的抗逆性研究提供参考,同时为构建节水型园林过程中合理利用氮肥提高植物抗旱性提供一定的理论依据。
1 材料与方法
1.1 材料
选择生长良好、长势较一致的无病虫害猴樟一年生实生苗为试验材料,采用上口径23.5 cm、底面直径25 cm、高27 cm的软塑双色花盆为移栽容器,盆底放置托盆。土壤采用湖南省常见的耕作土(粉碎后过3 mm筛),均匀装入盆中(每盆装土4.5 kg),而后灌水使土壤相对含水量保持在70%,并静置3 d使土壤自然沉降。以霍格兰营养液为氮源。
1.2 试验设计
试验设置2個氮素水平:不施氮(缺氮霍格兰营养液,N0)、施氮(全素霍格兰营养液,N1),每个区组设置3个重复,每个重复移栽10株苗木。
苗木移栽后采用常规养护管理缓苗,缓苗期间对N0苗木每天添加200 mL 缺氮霍格兰营养液,N1苗木每天添加200 mL 全素霍格兰营养液。缓苗15 d后停止浇水,分别于停止浇水0 d(正常水分,CK)、 3 d(轻度干旱,T1)、5 d(中度干旱,T2)、8 d(重度干旱,T3)测定猴樟幼苗生长及光合指标。
1.3 指标测定方法
1.3.1 生长指标的测定 分别于停止浇水处理0、3、5、8 d利用钢卷尺及数显式游标卡尺测定植株的苗高及地径,并将植株分地上、地下部收获,用清水洗净后于105 ℃杀青30 min,随后转移至65 ℃烘箱烘干至恒重,并测定植物干重。
1.3.2 叶片叶绿素含量的测定 分别于停止浇水处理0、3、5、8 d取猴樟苗木新鲜叶片(每个处理选取3棵植株,每棵植株选取2片叶片混合取样),立即放入低温保温箱保存,并于实验室中洗净表面污物,擦拭干净,用剪刀剪去中脉,剪成短丝状混匀(宽1 mm左右的细丝),将相同植株的叶片混匀。参考张宪政[14]的乙醇丙酮混合液法测定叶绿素含量。
1.3.3 气体交换参数测定 分别于停止浇水0、3、5、8 d的9:00—11:30测定。测定时,从每个处理组中选取5株苗木,每株苗木选择完全展开叶4片(分别位于植株东南西北4个方向)进行气体交换参数的测定。采用Li-6400XT型(Licor,USA)光合仪的2 cm×3 cm红蓝光叶室测定。将光合仪叶室的光合有效辐射设定为1 000 μmol/(m2·s),控制叶室空气温度在28~33 ℃,叶室空气湿度保持在40%~60%,保持大气CO2浓度。
1.4 数据统计分析
分别采用 Excel软件进行数据的记录及整理, 采用SPSS 26软件进行数据分析,采用Sigmaplot 14软件绘图。
2 结果与分析
2.1 干旱胁迫下氮素对猴樟幼苗生长发育的影响
2.1.1 干旱胁迫下氮素对猴樟幼苗苗高、地径的影响 由图1A可知,干旱胁迫会抑制猴樟幼苗苗高的增长,随着干旱程度的增加,幼苗苗高逐渐降低,且在中度及重度干旱胁迫下幼苗苗高显著降低,而轻度胁迫下幼苗苗高与正常水分条件下无显著差异。在轻度干旱胁迫下,外源施加氮肥可以显著提高猴樟幼苗的苗高(12.3%),但在中度及重度干旱胁迫下外源施加氮肥对猴樟幼苗苗高的增加无显著影响。
由图1B可知,干旱胁迫会抑制猴樟幼苗地径的增加,随着干旱程度的增加,幼苗地径逐渐降低,且在重度干旱胁迫下幼苗地径较正常水分和轻度干旱显著降低,而轻度及中度胁迫下幼苗地径与正常水分条件下无显著差异。此外,干旱胁迫条件下,外源施加氮肥可以在一定程度上促进猴樟幼苗地径的增加,但无显著差异。
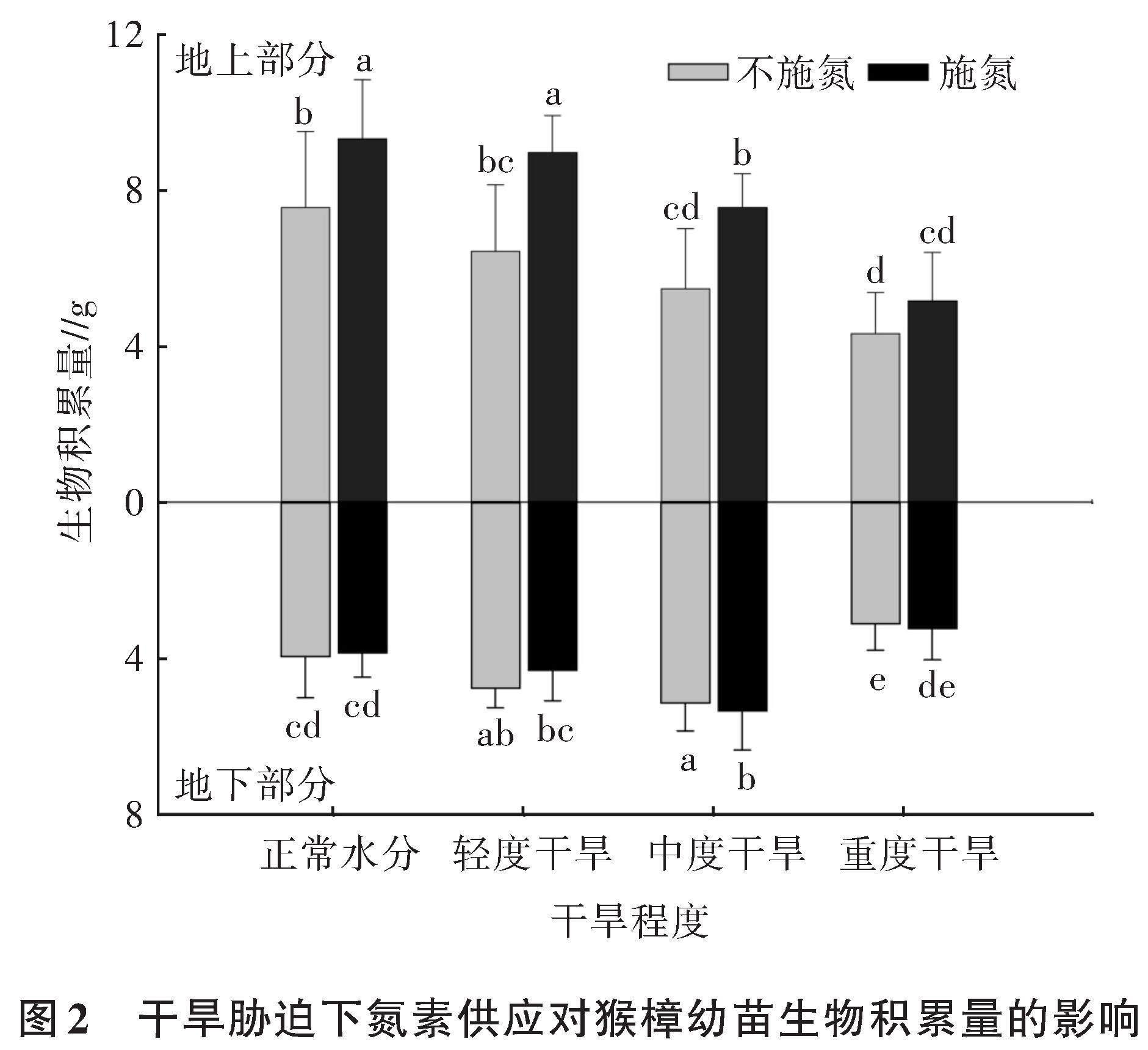
2.1.2 干旱胁迫下氮素对猴樟幼苗生物积累量的影响 如图2所示,随着干旱程度的增加,猴樟幼苗的生物积累量逐渐降低,且在中度及重度干旱胁迫下较正常水分显著降低,在中度、重度干旱胁迫下分别降低了27.5%、44.4%,而轻度干旱胁迫下地上部分生物积累量与对照组相比没有显著差异。在轻度和中度干旱胁迫下,外源施加氮肥可以显著提高猴樟幼苗地上部分生物积累量,分别提高了39.5%、38.0%,而在重度干旱胁迫下无显著提高。
对于地下部分(图2)而言,干旱程度较低时,干旱胁迫会促进猴樟幼苗地下部分生物积累量的增长,且在中度干旱胁迫下幼苗地下部分生物积累量增长最为显著(30.3%),重度干旱胁迫会显著抑制幼苗地下部分生物积累量的增长。此外,中度和重度干旱胁迫条件下,外源施加氮肥可以在一定程度上促进幼苗地下部分生物量积累,但无显著差异。而在轻度干旱胁迫下,外源施加氮肥能够在一定程度上抑制幼苗地下部分生物量积累,但差异不显著。
2.2 干旱胁迫下氮素对猴樟叶片叶绿素含量的影响
由图3A可以看出,随着干旱程度的加剧,幼苗叶片的叶绿素a含量逐渐降低。在轻度干旱和中度干旱胁迫下,外源施加氮肥可以显著提高猴樟幼苗叶绿素a的含量,分别提高了11.4%和5.5%,但在重度干旱胁迫下外源施加氮肥对猴樟幼苗叶绿素a含量的增加无显著影响。由图3B可以看出,在重度干旱胁迫下,猴樟叶片叶绿素b含量较对照组显著降低,而轻度和中度干旱对猴樟叶片叶绿素b含量没有显著影响。此外,在干旱胁迫条件下,外源施加氮肥可以在一定程度上促进猴樟幼苗叶绿素b含量的增加。由图3C可以看出,随着干旱程度的加剧,猴樟幼苗叶片的总叶绿素含量逐渐降低。在轻度和中度干旱胁迫下,外源施加氮肥可以显著提高猴樟幼苗总叶绿素含量,轻度、中度干旱胁迫下分别提高了9.7%和5.0%,但在重度干旱胁迫下外源施加氮肥对猴樟幼苗总叶绿素含量的增加无显著影响。由图3D可知,在干旱胁迫下,外源氮素的施加对叶绿素a与叶绿素b含量比值无显著影响。
2.3 干旱胁迫下氮素对猴樟叶片光合气体交换参数的影响
由表1可知,干旱胁迫会显著抑制猴樟幼苗净光合速率,且在中度及重度干旱胁迫下达到极显著水平。外源施加氮素可以显著提高干旱胁迫下猴樟幼苗的净光合速率,提高了11%~30%。
由于外界環境引起的干旱胁迫,猴樟幼苗气孔导度受到显著影响。外源添加氮素可以增加猴樟幼苗气孔导度,其中,在轻度干旱胁迫下,外源施加氮素可以显著促进猴樟幼苗气孔导度(15.7%),但在中度及重度干旱胁迫下,外源氮素对猴樟幼苗气孔导度的影响不显著。
猴樟幼苗叶片的胞间CO2浓度在干旱胁迫的作用下逐渐降低,且除施氮下的中度干旱外,猴樟叶片胞间CO2浓度在中度及重度干旱胁迫下与CK之间达显著或极显著水平。除重度干旱胁迫外,外源添加氮素会显著影响干旱胁迫下猴樟幼苗胞间CO2浓度,其中,猴樟幼苗叶片的胞间CO2浓度在轻度和中度干旱胁迫下受到外源施加氮素的作用而显著降低。
干旱胁迫会显著降低猴樟幼苗蒸腾速率,随着干旱程度的增加,幼苗蒸腾速率逐渐降低。在轻度和中度干旱胁迫下,外源施加氮肥可以显著提高猴樟幼苗蒸腾速率,但在重度干旱胁迫下外源施加氮肥对猴樟幼苗蒸腾速率的增加无显著影响。
3 小结与讨论
植物的生长发育过程受到外部环境因素的影响,外源施加氮素对猴樟幼苗的生长发育具有显著的促进作用。在猴樟幼苗生长发育过程中,通过施氮能够有效缓解干旱胁迫对猴樟幼苗造成的生长受阻[15]。本试验结果表明,在不同程度的干旱胁迫下,猴樟幼苗的苗高均表现为施氮>缺氮,可见施氮在一定程度上促进了猴樟幼苗地上部的生长,猴樟幼苗对干旱胁迫的响应得到了一定程度上的减缓,对干旱胁迫的适应性有所增强。此外,本试验中除重度干旱胁迫会显著抑制猴樟幼苗地径的增长外,其余干旱胁迫程度对猴樟幼苗地径均无显著影响,而外源施加氮素虽可以在一定程度上促进猴樟幼苗地径的增加,但亦无显著性差异。因此,猴樟幼苗地径对干旱胁迫并不敏感,外源氮素的添加对猴樟幼苗的地径生长无显著的促进作用。总体而言,氮素对猴樟幼苗苗高的促进作用大于地径。
结合猴樟幼苗地上部分及地下部分生物积累量的情况来看,干旱胁迫会显著抑制猴樟幼苗地上部分生物量的积累,此外,除重度干旱胁迫外,轻、中度干旱胁迫会显著促进猴樟地下部分生物量的积累。在干旱胁迫下,外源添加氮素可以有效促进猴樟幼苗地上部分生物量的积累,而对地下部分无明显促进作用。这与刘明丽[16]、谢志良等[17]、吴秀宁等[18]分别在玉米、棉花、小麦上的研究结论一致。因此,可以推测在适度的干旱胁迫下,猴樟幼苗可能通过将能量及物质集中作用于地下部分的生长,提高根系对外界水分的吸收,从而维持植物体内水分的平衡,而外源添加氮素主要表现为促进地上部分生物量的积累,提高叶片生物量,以维持较大的光吸收面积,表明施氮可以显著提高幼苗的净光合速率。
非气孔因素对植物的光合作用速率具有重要影响。本研究表明,猴樟幼苗叶片叶绿素含量会随着干旱胁迫的发生而降低,而净光合速率也相应下降。郑听等[19]对太子参的研究表明叶绿素含量与净光合速率呈正相关。叶绿素组成也在一定程度上受到干旱胁迫的影响,本研究结果表明,叶绿素a较叶绿素b对干旱胁迫的敏感度更高。在外源氮素的施加下,一定干旱程度下猴樟幼苗的叶绿素含量显著增加、净光合速率显著提高,但在重度干旱时,猴樟幼苗叶绿素含量在外源施加氮素条件下无显著增加,因此,可以认为干旱胁迫下猴樟幼苗净光合速率的降低可能是由于猴樟叶片的叶绿素含量降低导致。叶绿素含量的变化直接影响光合作用的速率[20],在一定程度的干旱胁迫下外源氮素的施加可以提高猴樟幼苗的叶绿素含量。
在植物进行光合作用时气孔因素也对净光合速率有限制作用。本研究结果表明,干旱胁迫下猴樟幼苗叶片胞间CO2浓度及气孔导度由于外界环境因素的影响均呈显著下降趋势,而蒸腾速率也相应显著下降,说明猴樟幼苗叶片气孔受到干旱胁迫的诱导发生闭合,这与Peeva等[21]的研究结果相似。外源添加氮素可以有效提高干旱胁迫下猴樟幼苗叶片的气孔导度,促进叶片对CO2的吸收。此外,本试验中也观测到在外源添加氮素的条件下,猴樟幼苗的气孔导度增加,而胞间CO2浓度却显著下降,净光合速率显著提高,进一步说明在施氮条件下,猴樟幼苗具有更高的CO2同化能力,与吴秀宁等[18]、张雅倩等[22]的研究结果一致。气孔关闭是植物本身的一种自我保护调节,以此来减少蒸腾作用失水,但光合作用速率也由于胞间CO2的固定减缓而下降,植物根系对外界环境中的水分和矿质元素的吸收也因为蒸腾拉力降低而受到影响[23]。因此,干旱胁迫通过降低叶绿素含量和诱导气孔关闭降低猴樟幼苗的净光合速率,而外源施加氮素可以促进猴樟幼苗叶绿素含量增加并促进叶片气孔开放。
本试验以猴樟幼苗为研究对象,研究其对干旱胁迫的生理响应及外源氮素对其抗旱性调控的效应。随着干旱胁迫的加剧,猴樟幼苗的苗高、地径和生物积累量均逐渐下降,外源氮素的施加可以促进苗高、地径的增长,并有效促进地上部分生物量的积累,但对地下部分生物量的积累无明显促进作用。干旱胁迫的加剧会导致猴樟幼苗叶绿素a、叶绿素b 的含量均逐渐下降,且外源施加氮素之后叶绿素各指标均能有一定程度的提高。此外,干旱胁迫降低了猴樟幼苗的Pn、Ci、Tr、Gs,外源施加氮素后,幼苗的净光合速率明显提高,外源氮素的施加促进了叶片气孔开放,在一定程度上提高了猴樟幼苗对干旱胁迫的适应性。
参考文献:
[1] 李 芳.西安城市道路绿地耐旱型草本植物景观设计研究[D].西安:西安建筑科技大学,2021.
[2] 邵 帅.菊芋对土壤逆境胁迫的响应及氮素的调控效应研究[D].哈尔滨:东北林业大学,2016.
[3] YANG X Y,LU M Q,WANG Y F,et al.Response mechanism of plants to drought stress[J]. Horticulturae,2021,7(3):50.
[4] GUPTA A,RICOMEDINA A,CA?ODELGADO A I. The physiology of plant responses to drought[J].Science,2020,368(6488):266-269.
[5] 杨明凤,王金梅,吉春容,等.干旱胁迫对棉花生长发育和光合荧光参数的影响[J].中国农学通报, 2021,37(13):22-28.
[6] 韩志平,张海霞,张 巽,等.水分胁迫对黍子幼苗生长和生理特性的影响[J].中国农业气象, 2019,40(8): 502-511.
[7] ZHANG X B,LEI L,LAI J S,et al. Effects of drought stress and water recovery on physiological responses and gene expression in maize seedlings[J].BMC plant biology,2018,18(1):68.
[8] LEI D,LU Z F,GAO L M,et al. Is nitrogen a key determinant of water transport and photosynthesis in higher plants upon drought stress?[J].Frontiers in plant science,2018(9):1143.
[9] 王亦豪,石丹丹,欧阳翔,等.氮素调控对不同穗型超级早稻光合特性和产量的影响[J].杂交水稻, 2021,36(4): 88-95.
[10] 张立新,李生秀.长期水分胁迫下氮、钾对夏玉米叶片光合特性的影响[J].植物营养与肥料学报, 2009,15(1): 82-90.
[11] 于志民,刘 玮,邓志平,等.不同类型及规格育苗容器对猴樟1年生苗生长的影响[J].南方农业学报,2016,47(12):2118-2123.
[12] 程许娜.猴樟对低温胁迫的生理响应及其耐寒性分析[D].郑州:河南农业大学, 2012.
[13] 刘长成,刘玉国,郭 柯.四种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性[J].植物生态学报,2011,35(10):1070-1082.
[14] 张宪政.植物叶绿素含量测定——丙酮乙醇混合液法[J].辽宁农业科学,1986(3):26-28.
[15] PALTA J A,CHEN X,MILROY S P,et al. Large root systems: Are they useful in adapting wheat to dry environments?[J].Functional plant biology,2011,38(5):347-354.
[16] 刘明丽.干旱胁迫下氮素对玉米生理特性的影响[D].济南:山东师范大学,2016.
[17] 谢志良,田长彦.膜下滴灌水氮对棉花根系形态和生物量分配变化的影响[J].干旱区资源与环境, 2010,24(4):138-143.
[18] 吴秀宁,刘 英,王新军,等.干旱胁迫下氮素对小麦幼苗生长及光合生理的影响[J].湖北农业科学, 2020,59(8): 21-24.
[19] 郑 听,熊鹏飞.遮阴对太子参叶片叶绿素含量和净光合速率影响[J].贵州科学,2021,39(3):24-27.
[20] 刘振威,高 佳,孙 丽,等.南瓜叶片叶绿素含量与净光合速率的关系[J].河南科技学院学报(自然科学版), 2008,36(4): 27-29.
[21] PEEVA V,CORNIC G. Leaf photosynthesis of Haberlea rhodopensis before and during drought[J].Environmental & experimental botany,2009,65(2-3):310-318.
[22] 张雅倩,林 琪,刘家斌,等.干旱脅迫对不同肥水类型小麦旗叶光合特性及产量的影响[J].麦类作物学报,2011,31(4):724-730.
[23] 黄琳琳.干旱胁迫和不同氮素水平对苹果根系氮素吸收和代谢的影响研究[D].陕西杨凌:西北农林科技大学,2018.
猜你喜欢
舰船科学技术(2022年11期)2022-07-15 07:51:56
西藏农业科技(2019年3期)2019-11-04 00:35:10
现代园艺(2018年3期)2018-02-10 05:18:12
新课程·下旬(2017年7期)2017-08-14 11:12:50
上海农业学报(2017年3期)2017-04-10 12:39:12
西北林学院学报(2015年5期)2015-01-03 05:12:20
植物营养与肥料学报(2011年5期)2011-11-06 07:30:52
植物营养与肥料学报(2011年2期)2011-10-26 03:52:30
植物营养与肥料学报(2011年2期)2011-10-26 03:52:10
植物营养与肥料学报(2011年3期)2011-10-24 06:15:00
