
氮素水平对籼粳杂交稻产量和贮藏蛋白的影响
2024-05-07 04:40孙建龙张新城任韵王丰颖陆鸿英钱伟红马善林
浙江农业科学 2024年4期
孙建龙,张新城,任韵,王丰颖,陆鸿英,钱伟红,马善林*
(1.吴兴尹家圩粮油植保农机专业合作社,浙江 湖州 313000;2.湖州市农业科学研究院,浙江 湖州 313000;3.湖州市农业种质资源创新与应用重点实验室,浙江 湖州 313000;4.吴兴区农业技术推广服务中心,浙江 湖州 313000)
近年来,水稻育种目标从高产为主到高抗、优质和高产并重,育种理念也从高产优质逐步提升为“少投入、多产出、重环保”[1]。籼粳杂交稻具有营养生长旺盛、茎秆粗壮、颖花数多、库容量大、增产潜力大等特点[2]。然而其产量潜力的充分实现通常依赖高氮投入,高氮不仅对粒型有影响,同时改变蛋白组分比例,给稻米品质改良带来不利影响[3-4]。
施用氮肥是提高水稻产量最常用的栽培措施。研究表明,产量通常先随施氮量增加而增加,随后呈下降趋势,最佳施氮量因品种而异[5-7]。水稻粒型包括粒长、粒宽和粒厚,决定着千粒重大小,并以此来影响水稻产量。水稻籽粒依据粒长,可分为特长粒(>9.1 mm)、长粒(8.1~9.0 mm)、中长粒(7.1~8.0 mm)和短粒(<7 mm)4 类;依据长宽比,可分为细长形(>3.1)、椭圆形(2.21~3.0)、卵圆形(1.8~2.2)和短圆形(<1.8)4 类[8]。粒长与粒宽的比值(长宽比)不仅是籽粒形状的一种表现形式,而且会影响水稻品质[3,9]。
过量施氮或氮肥后移会显著增加籽粒蛋白质,导致米饭变硬,降低稻米食味品质[6,10-11]。不同穗型籼粳杂交稻的直链淀粉含量差异不大,而蛋白质含量差异较大,是决定食味品质的关键因子[12]。优良食味水稻品种籽粒蛋白质含量通常较低,且随着氮素水平的增加而上升,各蛋白组分含量与食味品质的相关性因水稻类型而异[13]。蒋晶晶等[14]也发现,超级稻弱势粒中蛋白质含量高于强势粒,硬度增大,口感较差,通过优化栽培模式可以提高弱势粒食味品质,从而提升整穗品质。研究进一步发现,籼粳杂交稻弱势粒中醇溶蛋白含量显著高于强势粒,而谷蛋白无显著差异,两种蛋白组分随施氮量增加而增加,醇溶蛋白是稻米食味变差的主要原因[3]。
随着蛋白质或蛋白组分含量在蒸煮食味品质研究中逐步得到重视,对强弱势粒分别研究有助于为大穗型超级稻的遗传改良和高效栽培提供理论依据。然而,以往研究多集中在静态样品(成熟期),对强弱势粒蛋白组分积累动态及其对氮素的响应关注并不多,蛋白质在胚乳中的分布也鲜见报道。基于此,本研究以大穗型超级稻甬优12和甬优17为材料,比较了不同氮肥水平下产量、粒型、粒重动态、强弱势粒的蛋白组分积累动态和蛋白质分布差异,研究结果可为籼粳杂交稻高产优质栽培提供参考。
1 材料与方法
1.1 材料种植
本试验以籼粳杂交稻甬优12和甬优17为材料,均由宁波市种子有限公司育成。试验于2018年在浙江大学长兴试验站进行,土壤为黏壤土,全氮含量1.05 g·kg-1,速效磷含量20.43 mg·kg-1,速效钾含量102.5 mg·kg-1。5月20日播种,6月20日移栽,栽插密度为30.0 cm×16.5 cm,双苗移栽。设置4个氮肥水平,即对照(N0,不施氮)、低氮(LN,100 kg·hm-2)、中氮(MN,200 kg·hm-2)、高氮(HN,300 kg·hm-2),3个重复。常规管理按当地高产栽培方式管理。
1.2 样品获取
于抽穗旺盛期对开始抽穗且大小基本一致的穗子进行挂牌标记,记为花后0 d,并对标记的穗子于花后7、14、21、28、35、42 d进行动态取样,共取6次。按Zhang等[3]的方法分为强弱势粒样品,烘干至恒重,测定粒重,随后将样品手工去壳后磨粉用于蛋白组分分析。成熟期收获水稻籽粒,自然晾干,部分样品用于测定稻谷长宽,去壳脱糙,少量糙米用于考马斯亮蓝染色观察和蛋白质组分测定,部分精研后用于测定精米长宽。
1.3 产量分析
成熟期调查每穴穗数,每个小区调查 30 穴,取代表性材料3穴进行考种,得到穗数、每穗粒数、结实率和千粒重,并计算理论产量。
1.4 粒型测定
将10粒稻谷和精米首尾相连或背腹紧密相连,用尺子测量长度和宽度,并计算平均粒长、粒宽和长宽比。
1.5 蛋白组分测定
4种蛋白组分按照清蛋白、球蛋白、醇溶蛋白和谷蛋白的顺序依次提取,参考Zhang等[3]方法。
1.6 切片染色观察
先将成熟糙米浸泡在2.5%戊二醛固定液中12 h,用剃须刀切开种子,样品继续固定48 h。样品用磷酸盐缓冲液冲洗3次,然后用梯度乙醇(30%、50%、70%、90%、100%)连续脱水,再用梯度树脂(25%、50%、75%、100%×2)渗透样品,包埋在伦敦白胶树脂中。60 ℃聚合48 h,用徕卡超薄切片机切成2 μm的半薄切片,将切片放置1%的考马斯亮蓝R250中染色20 min,随后用7%的乙酸溶液清洗掉表面的染液,并用蒸馏水清洗干净,低温烤干,置于光学显微镜观察并拍照。
1.7 数据分析
试验数据采用Excel 2019软件进行整理与计算,并用 SPSS17.0软件进行统计分析,采用Duncan法进行多重比较(P<0.05)。
2 结果与分析
2.1 氮素水平对产量及其构成因子的影响
施氮显著提高两品种的产量,品种对氮素的响应存在差异,甬优17对氮素响应较甬优12更敏感。甬优17产量随施氮量的增加而增加,而甬优12在氮素处理间无显著差异(P>0.05),在低氮条件下就能获得较高的产量(表1)。随施氮量的增加,两品种穗数均显著提高(P<0.05)。与对照(N0)相比,甬优12和甬优17在高氮条件下分别增加45.54%和71.90%,但两品种平均穗数基本相当。氮素水平对穗粒数和结实率均无显著影响,但品种间存在显著差异(P<0.05)。甬优12每穗粒数高达459.6,显著高于甬优17的373.7,而甬优12的结实率(82.0%)则显著低于甬优17(88.3%)。方差分析表明,穗数主要受氮素调控,穗粒数、结实率、千粒重主要由基因型决定,千粒重亦受氮素调控(表1)。

表1 氮素水平对产量及其构成因子的影响Table 1 Effects of different nitrogen treatments on yield and its components
2.2 氮素水平对粒形和籽粒灌浆的影响
甬优12稻谷粒长和长宽比分别为7.80~7.89和2.41~2.45,分别属于中长粒和椭圆形;甬优17为8.40~8.52和2.66~2.70,分别属于长粒和椭圆形。不论是精米还是稻谷,甬优12的粒长均显著小于甬优17,而粒宽反之,从而导致较小的长宽比(表2)。低氮对粒长和粒宽没有显著影响,中高氮处理差异显著(P<0.05),但幅度较小,导致长宽比和粒重轻微下降,但氮素均未能引起粒型变化(表2)。就平均粒重而言,强势粒显著高于弱势粒,甬优17略高于甬优12。在灌浆前17~18 d,粒重几乎呈线性增长,28 d后几乎不再增加(图1中 a和b)。方差分析也表明,粒型主要由基因型决定,同时受氮素调控,但影响较小。

a图表示甬优12,b图表示甬优17。SG表示强势粒;IG表示弱势粒;YY表示甬优。图2同。图1 强、弱势粒增重动态以及成熟期强、弱势粒醇溶蛋白和谷蛋白含量Fig.1 Grain weight during grain filling stage,prolamin and glutelin content of superior and inferior grains at maturity

表2 氮素水平对精米和稻谷粒型的影响Table 2 Effects of different nitrogen treatments on grain types of milled rice and paddy
2.3 氮素对贮藏蛋白最终含量和籽粒积累动态的影响
强弱势粒间,甬优12除醇溶蛋白含量在低氮条件和谷蛋白含量在高氮条件下差异显著外(P<0.05),其他处理均不显著;甬优17醇溶蛋白含量在4个氮素处理中均差异显著,而谷蛋白含量仅在中高氮条件差异显著(图1中 c和d)。氮素处理间,甬优12贮藏蛋白(醇溶蛋白和谷蛋白)含量仅在高氮条件显著升高,而甬优17在低氮条件就显著提高(P<0.05),表明甬优17更易受氮素影响,尤其是弱势粒。值得注意的是,甬优12强势粒醇溶蛋白含量高于甬优17,而弱势粒反之,谷蛋白含量甬优17高于甬优12。
甬优12醇溶蛋白积累量呈现先上升后下降再上升的波浪趋势,而甬优17在花后28 d或35 d达到峰值后呈下降趋势(图2中a和b)。与甬优12不同的是,甬优17弱势粒中醇溶蛋白积累量在施氮条件下明显上升,缩小了与强势粒的差距(图3),主要与其含量显著提高有关(图1中 c)。有趣的是,两品种间谷蛋白积累的模式恰好与醇溶蛋白积累模式相反,尽管达到峰值的时间略有差异(图2)。

a和c图表示甬优12;b和d图表示甬优17。图2 强弱势粒醇溶蛋白和谷蛋白积累动态Fig.2 Prolamin and glutelin contents of superior and inferior grains
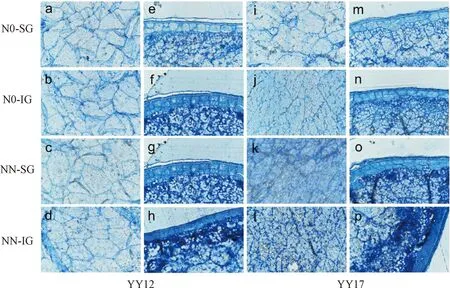
a~d和i~l表示胚乳中心,e~h和m~p表示胚乳外层。图3 强弱势粒不同部位的蛋白质分布Fig.3 The protein distribution in different parts of superior and inferior grains
2.4 氮素对强弱势粒中蛋白质积累部位的影响
经考马斯亮蓝染色后,可以清晰看到甬优12内胚乳中蛋白质积累在强弱势粒间差异不明显(图3中a~d),而在胚乳外层弱势粒明显高于强势粒,且施氮促进蛋白质积累(图3中e~h)。甬优17在不施氮条件下,强弱势粒间内胚乳中蛋白质积累差异不明显(图3中i~j),而弱势粒胚乳外层略有增加(图3中m~n)。在高氮条件下,内胚乳中和胚乳外层蛋白质积累均明显增加,且胚乳外层尤为突出。以上结果表明,贮藏蛋白主要富集在籽粒胚乳外层,弱势粒蛋白质浓度高于强势粒,甬优17对氮素响应较甬优12更敏感,印证了以上结果(图1中c~d)。
3 讨论
3.1 产量与粒型对氮素的响应
施用氮肥是提高产量最主要的栽培措施。众多研究表明,产量先随施氮量增加而增加,随后呈下降趋势,最佳施氮量因品种而异[5-7]。本研究中,施氮显著提高了产量,主要由穗数的大幅提升引起(表1),与前人结果一致[15-16]。产量在高氮条件(300 kg·hm-2)下未曾下降,可能与籼粳杂交稻营养生长旺盛、颖花数多、库容量大、耗氮量大等特点有关[2]。粒重主要由粒型(包括粒长、粒宽和粒厚)决定,且与灌浆程度有关,遗传力为 40%~60%,遗传稳定,受外界环境因素影响较小[8]。本研究中,稻谷的粒长、粒宽和长宽比在中高氮条件下虽略有下降,但幅度不大,仍未能引起粒型变化,从而导致粒重变化也不大(表1和表2),证明了粒型和粒重的高遗传力[8,17]。然而,强弱势粒间粒重差异显著,这主要是由于灌浆程度不同引起(图1中a~b)。千粒重不仅取决于籽粒库容量(粒型)也取决于其充实度(灌浆)。通常,弱势粒灌浆起步迟,灌浆不充分,大穗型超级稻中尤为突出[18]。因此,整穗粒长或粒宽在高氮条件下有所下降,可能与弱势粒比例大幅增加有关[3]。
3.2 蛋白组分积累动态与分布特征
众所周知,施氮显著增加蛋白质含量[6,10-12,19]。多数研究表明,弱势粒中蛋白质含量显著高于强势粒[3,20-21],但Ma等[22]研究结果与之相反,这可能与研究材料不同有关。本研究结果与前人基本一致[20-21],另外还对蛋白积累动态进行检测发现,醇溶蛋白与谷蛋白积累模式呈互补状态(图2),稻米中蛋白质的合成处于动态平衡状态,当某一贮藏蛋白被抑制,其他贮藏蛋白则会相应增加[23]。水稻强弱势粒间贮藏蛋白(醇溶蛋白和谷蛋白)浓度在品种间存在差异,甬优12差异较小,而甬优17弱势粒显著高于强势粒,且甬优17对氮素更敏感。另外,不论强弱势粒还是施氮与否,甬优12蛋白质积累在内胚乳中均没有明显差异,而甬优17在高氮下,内胚乳中蛋白质积累显著增加,弱势粒中尤为明显,这也正好解释了甬优17稻米食味品质在高氮下急剧下降的原因[3]。再则,贮藏蛋白主要富集在胚乳外层,表明碾磨程度大,将会去除大量蛋白质,有利于食味品质的提升,但会牺牲部分精米产量。
4 结论
施氮均能显著提高产量和蛋白质积累量,但品种对氮素响应存在差异。甬优12在低氮下就能获得较高产量,有望实现高产优质的平衡。高氮能影响粒长或粒宽,但无法改变粒型。醇溶蛋白和谷蛋白积累存在互补效应,有利于维持贮藏蛋白的相对平衡。弱势粒蛋白质浓度尽管高于强势粒,但因其粒重小导致总积累量仍然低于强势粒。贮藏蛋白主要富集在籽粒胚乳外层,甬优17对氮素响应较甬优12更敏感。
猜你喜欢
中国油脂(2022年1期)2022-02-12
光谱学与光谱分析(2021年11期)2021-11-11
广东农业科学(2021年3期)2021-04-23
中国食品学报(2021年3期)2021-04-22
江苏农业学报(2020年2期)2020-05-27
山东农业科学(2018年11期)2018-01-17
江苏农业科学(2016年6期)2016-07-25
广东农业科学(2016年11期)2016-03-29
华北农学报(2016年1期)2016-03-18
农业科技与装备(2015年5期)2015-05-30
