
元宝枫种仁多糖提取纯化、结构表征以及降糖活性研究
2024-04-29 06:39米圣成徐晓杰魏菱鸽朱明华包怡红陈春霞
食品工业科技 2024年8期
米圣成,徐晓杰,魏菱鸽,路 祺,朱明华,包怡红,陈春霞
(1.东北林业大学生命科学学院,黑龙江哈尔滨 150000;2.东北林业大学化学与资源利用学院,黑龙江哈尔滨 150000;3.哈尔滨职业技术学院,黑龙江哈尔滨 150000)
我国Ⅱ型糖尿病(T2DM)患者占总糖尿病患的90%以上,T2DM 发病机制尚不明确,但人们普遍认为胰腺β细胞功能障碍和胰岛素抵抗是两个主要因素[1]。此外,氧化应激在T2DM 的发病机制中发挥作用。通常用于治疗这种疾病的药物是西药,这些药物往往具有副作用大、选择有限和成本高等[2]缺陷。张培莲[3]的研究发现使用胰岛素治疗糖尿病后会出现低血糖、体重增加、水肿、脂肪组织增生以及脂肪萎缩等不良反应。植物多糖具有多种生物学功能,如改善肠道健康、增强免疫力、抗氧化、抗病毒和抗应激等[4]作用。随着人们生活水平的提高和科技的进步,植物多糖在食品[5]、医药[6]和材料等[7]领域得到了广泛应用。马宁珠等[8]发现虎杖多糖能够显著降低糖尿病大鼠的糖耐量和空腹血糖。赵凯迪等[9]发现桔梗多糖可以通过改善T2DM 大鼠代谢水平和氧化应激水平起到降血糖作用。
元宝枫为槭树科槭树属的植物,因翅果形状像中国古代金锭“元宝”得以命名。2011 年我国卫生部(如今的国家卫生健康委员会)发布的“关于批准元宝籽油和牡丹籽油作为新资源食品的公告”,标志着元宝枫籽油正式成为被国家承认的食用油;2015 年国务院办公厅发布《关于加快木本油料产业发展的意见》(国办发[2014]68 号)重点提及加快元宝枫等木本油料作物产业的综合发展。近年来,科研工作者发现元宝枫种仁中除了富含有油脂[10],还含有蛋白质[11]和多糖[12],目前关于元宝枫种仁的研究还主要集中在油脂提取和其功能性脂肪酸—神经酸的纯化等方面。而对元宝枫种仁多糖的研究,仅仅局限于对元宝枫种仁多糖的提取方面,对元宝枫种仁多糖分离纯化、单糖组成、以及生物活性等方面的研究却鲜有报道。
本实验利用水提醇沉法从元宝枫种仁中提取多糖,并通过单因素和响应面法对元宝枫种仁多糖提取工艺进行优化。经Sevage 法脱蛋白、DEAE-DE 纤维素52 层析柱和SephadexG-100 凝胶层析柱分离纯化元宝枫种仁多糖,并对所获得多糖进行了红外光谱分析、同步热分析、单糖组成分析、α-淀粉酶抑制活性以及降血糖等活性检测,为元宝枫种仁多糖的开发和应用提供参考和数据支撑。
1 材料与方法
1.1 材料与仪器
元宝枫种仁 购置于陕西金旺农林科技有限公司;DEAE-DE 纤维素52、SephadexG-100 凝胶、阿卡波糖(BR,95%)、α-淀粉酶(50 U/mg)、噻唑蓝(Thiazolyl Blue,MTT)(BR,95%)、单糖标准品 美国Sigma 公司;胎牛血清(Fetal Bovine Serum,FBS) 浙江天杭生物科技有限公司;MEM 培养基 美国Gibco 公司;二甲基亚砜(Dimethyl Sulfoxide,DMSO)Biosharp 公司;葡萄糖(Glucose,GLU)试剂盒、糖原(Glycogen,Gn)测定试剂盒、己糖激酶(Hexokinase,HK)试剂盒、丙酮酸激酶(Pyruvate Kinase,PK)试剂盒、3,5-二硝基水杨酸(3,5-Dinitrosalicylic acid,DNS)南京建成生物工程研究所;HepG2 细胞 上海瑾元生物科技有限公司;其他试剂 均为国产分析纯。
IRAffinity-1 傅里叶变换红外光谱 日本岛津公司;STA449F5 同步热分析仪 德国耐驰仪器制造有限公司;Reacti-thermo 氮气吹扫仪 美国Thermo 公司;ICS5000+离子色谱仪 美国Thermo公司;HZQ-F 全温振荡培养箱 哈尔滨东联电子技术有限公司;Infinite 200 PRO 酶标仪 长沙泰肯生物技术有限公司;CYTATION5 高内涵成像分析系统 美国伯腾仪器有限公司;HL-2B 数显恒流泵上海驰唐电子有限公司;UV-5500 紫外分光光度计上海元析仪器有限公司。
1.2 实验方法
1.2.1 元宝枫种仁多糖的制备工艺研究
1.2.1.1 元宝枫种仁多糖的提取 元宝枫种仁于60 ℃烘干后,粉碎过20 目筛,室温存储备用。精确称取上述元宝枫种仁粉末100 g 于圆底烧瓶中,加入1500 mL 超纯水,用加热套回流煮沸4 h 后冷却,以5000 r/min 离心10 min,收集上清液,浓缩至50 mL 左右,加入200 mL 无水乙醇在4 ℃下醇沉12 h,在6000 r/min 离心10 min,回收多糖,使用超纯水重新溶解多糖,透析72 h,冻干得元宝枫多糖。
1.2.1.2 元宝枫种仁多糖得率 计算采用苯酚-硫酸法测定多糖(PATM)的质量浓度,并按公式(1)计算,得到PATM 质量分数得率。
式中:Y 表示提取多糖PATM 的得率,%;ρ表示多糖质量浓度,mg/mL;n 表示稀释倍数;V 表示多糖溶液的体积,mL;m 表示样品绝干质量,mg。
1.2.2 单因素实验 采用水提醇沉法提取PATM,以多糖得率为指标,对提取各因素进行研究。
1.2.2.1 提取次数 固定提取温度80 ℃,提取时间3 h,液固比30:1 mL/g,考察提取次数(提取1、2、3 次)对多糖(PATM)得率的影响。
1.2.2.2 提取温度 固定液固比30:1 mL/g,提取时间3 h,提取1 次,考察不同提取温度(60、70、80、90、100 ℃)对多糖(PATM)得率的影响。
1.2.2.3 提取时间 固定液固比30:1 mL/g,提取温度80 ℃,提取1 次,考察不同提取时间(1、2、3、4、5 h)对多糖(PATM)得率的影响。
1.2.2.4 液固比 固定提取温度80 ℃,提取时间3 h,提取1 次,考察不同液固比(10:1、15:1、20:1、30:1、40:1 mL/g)对多糖(PATM)得率的影响。
1.2.3 响应面优化试验 根据单因素实验结果,提取次数固定为1 次。选取提取温度(℃)、提取时间(h)、液固比(mL/g)三个因素,以PATM 得率为响应值,运用Box-Behnken 进行响应面试验优化设计[13],其试验因素与水平设计见表1。
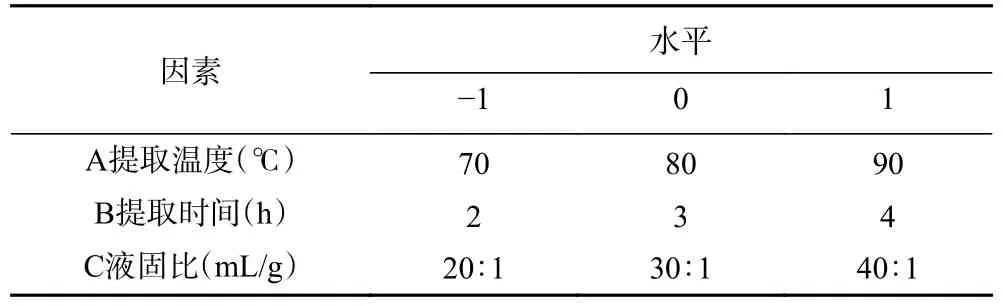
表1 响应面试验因素与水平Table 1 Response surface test factors and levels
1.2.4 元宝枫种仁多糖的分离纯化
1.2.4.1 元宝枫种仁多糖脱蛋白 采用Sevage 试剂[V(三氯甲烷):V(正丁醇)=4:1]脱除蛋白[14],将PATM 溶液与Sevage 试剂混合搅拌30 min,以5000 r/min 离心10 min,收集上清液,重复至蛋白无析出,上清液于旋转蒸发仪去除有机试剂,收集多糖溶液,用3500 Da 透析袋流水透析24 h。将透析后的多糖溶液进行浓缩、冻干,放置干燥器中以进行后续实验。
1.2.4.2 纤维素层析柱分离元宝枫种仁多糖 称取500 mg 除蛋白的PATM,配置成质量浓度为10 mg/mL 的多糖溶液。将多糖溶液缓慢注入DEAE-DE纤维素52 层析柱中,依次用超纯水、0.1、0.3、0.5 mol/L 的NaCl 溶液进行洗脱,流速为1 mL/min,每管4 mL。收集不同洗脱条件的多糖组分,采用苯酚-硫酸法处测定每管中多糖含量,记录OD 值。根据峰值段收集洗脱液,减压浓缩,用透析袋流水透析24 h 后冻干,得到4 个多糖组分:PATM-1、PATM-2、PATM-3 和PATM-4。
1.2.4.3 凝胶层析柱纯化元宝枫种仁多糖 采用SephadexG-100 凝胶层析柱对主要多糖成分PATM-3 进一步分离纯化,上样浓度为5 mg/mL,缓慢注入SephadexG-100 凝胶层析柱,0.2 mL/min 的流速去离子水进行洗脱,每管4 mL。采用苯酚-硫酸法测定洗脱液在490 nm 处吸光值,并绘制洗脱曲线[15]。根据峰值段收集洗脱液,将收集洗脱液浓缩和冷冻干燥,得到纯化组分PATM-3-1。
1.2.5 多糖和蛋白含量测定 以葡萄糖为标准品,采用苯酚-硫酸法测定PATM-3-1 多糖含量[16],其回归方程Y=1.0536X+0.0064(R2=0.9932);使用BCA 试剂盒检测PATM-3-1 多糖中蛋白质含量。
1.2.6 单糖组成测定 准确称量PATM-3-1 多糖样品5 mg,加入2 mol/L 三氟乙酸(TFA)溶液1 mL,105 ℃加热6 h。超纯氮气吹干,加入甲醇清洗,在通过超纯氮气吹干,重复甲醇清洗2~3 次。加入超纯水溶解,并通过0.22 μm 微孔膜转入色谱瓶中进行测量。HPAEC 条件为:Thermo ICS5000 离子色谱系统(ICS5000),利用电化学检测器对单糖组分进行分析检测。采用Dionex™ CarboPac™ PA10(250 mm×4.0 mm,10 μm)液相色谱柱;进样量为5 μL。流动相A(0.1 mol/L NaOH),流动相B(0.1 mol/L NaOH,0.2 mol/L NaAc),流速0.5 mL/min;柱温设置为30 ℃;洗脱梯度:0 min A 相/B 相(95:5 V/V),30 min A 相/B 相(80:20 V/V),30.1 min A 相/B 相(60:40 V/V),45 min A 相/B 相(60:40 V/V),45.1 min A 相/B 相(95:5 V/V)。数据在ICS5000 离子色谱仪上采集,并使用Chromeleon 7.2 CDS 进行处理。
1.2.7 红外光谱分析 称取2 mg 多糖样品(PATM-3-1),与200 mg 的干燥后的溴化钾在玛瑙研钵中研磨均匀,放入压片机压成薄片。以溴化钾片作为背景去除干扰,在分辨率为4 cm-1在4000~500 cm-1范围内进行红外光谱扫描[17]。
1.2.8 热重分析 采用同步热分析法分析纯化多糖的热稳定性。称取5 mg 多糖样品(PATM-3-1),放于坩埚中,以10 ℃/min 加热速率,在20~800 ℃范围进行热重分析[18]。
1.2.9 刚果红实验 采用刚果红法分析纯化多糖的三螺旋结构。取2 mL(1 mg/mL)纯化多糖溶液与2 mL(160 µmol/L)刚果红溶液混合,依次加入不同浓度(0、0.1、0.3、0.4、0.5 mol/L)的NaOH 溶液,室温静置15 min,600~400 nm 范围进行全波长扫描,测定溶液最大吸收波长。以NaOH 浓度为横坐标,以最大吸收波长为纵坐标,并绘制曲线[19]。
1.2.10 体外降糖活性研究
1.2.10.1α-淀粉酶抑制率测定 根据文献[20]描述的方法,对多糖PATM-3-1 进行α-淀粉酶的抑制率测定。用去离子水将多糖配置成不同质量浓度的溶液。以不同浓度阿卡波糖为阳性对照,取250 µL 多糖溶液,与40 µLα-淀粉酶(5 U)充分混合,37 ℃水浴10 min 后,加入质量分数为1%可溶性淀粉溶液500 µL,继续37 ℃水浴5 min,取出后加入600 µL DNS 试剂终止反应,并在沸水浴加热10 min,冷却至室温,540 nm 处检测吸光值,按公式(2)计算α-淀粉酶的抑制率。
式中:Y 表示为α-淀粉酶抑制率,%;A0表示用去离子水代替样品的吸光值;A1表示被测样品反应后的吸光值;A2表示去离子水代替α-淀粉酶溶液混合后的吸光值。
1.2.10.2 胰岛素抵抗细胞模型的建立 取对数生长期的HepG2 细胞,用0.25%胰酶进行消化,加入完全培养基调节成1×105个/mL 的细胞悬液,接种于96 孔板,每孔100 µL,细胞贴壁后,弃去培养基,加入含有不同浓度胰岛素的无血清MEM 培养基(10-9、10-8、10-7、10-6、10-5mol/L)100 µL,培养36 h 根据细胞的葡萄糖消耗量来确定最佳的胰岛素浓度。
1.2.10.3 细胞存活率测定 采用MTT 法[21]测定多糖PATM-3-1 对HepG2 细胞存活率的影响。将PATM-3-1 用MEM 培养基配制成不同质量浓度备用。将1×105个/mL 的HepG2 细胞悬液加入96 孔板,每孔100 µL。放入37 ℃、5% CO2的培养箱中培养24 h 后,加入100 µL 不同质量浓度(2560、1280、640、320、160、80 µg/mL)的PATM-3-1 继续培养24 h,加入5 mg/mL MTT 溶液50 µL 继续在培养箱中培养3 h,弃去上清液,加入150 µL DMSO 溶液,充分振荡10 min,490 nm 处检测吸光值[22]。并按照公式(3)计算HepG2 细胞存活率。
式中:Y 表示细胞存活率,%;A0表示无细胞孔的吸光值;A1表示含有HepG2 细胞和被测样品的吸光值;A2表示含有HepG2 细胞的吸光值。
1.2.10.4 细胞培养与分组 在37 ℃、5% CO2的培养条件下,用完全培养基(89% MEM 培养基、10%FBS 和1%青霉素-链霉素)培养HepG2 细胞。当细胞贴壁80%后,用0.25%胰酶进行消化传代,并选取对数生长期细胞进行实验。取对数生长期HepG2细胞,加入完全培养基调节成1×105个/mL 的细胞悬液,接种于6 孔板中每孔1 mL。将细胞分为六组:空白组、模型组(10-7mol/L 胰岛素)、阳性对照组(1 mg/mL 二甲双胍+10-7mol/L 胰岛素)、PATM-3-1低剂量组(320 μg/mL PATM-3-1+10-7mol/L 胰岛素)、PATM-3-1 中剂量组(640 μg/mL PATM-3-1+10-7mol/L 胰岛素)、PATM-3-1 高剂量组(1280 μg/mL PATM-3-1+10-7mol/L 胰岛素)。
1.2.10.5 葡萄糖消耗量和糖原含量测定 按照1.2.10.4 中的细胞分组进行培养,用0.25%胰酶消化,收集细胞离心弃上清液,用PBS 清洗细胞1~2 次,离心保留沉淀细胞。向沉淀细胞中加入0.1 mol/L pH7.4 的磷酸盐缓冲溶液,冰水浴下超声破碎。按照葡萄糖(GLU)、糖原(Glycogen)和蛋白定量(TP)试剂盒说明进行检测。
1.2.10.6 己糖激酶(HK)和丙酮酸激酶(PK)活性测定 实验分组同1.2.10.4,用0.25%胰酶消化后,收集、离心、保留沉淀细胞待用,后续操作按照己糖激酶(HK)、丙酮酸激酶(PK)和蛋白定量(TP)试剂盒说明进行检测。
1.3 数据处理
使用Design-Expert 13.0.1 软件进行响应面试验设计及试验数据处理,并对实验数据进行分析。每组实验3 个平行,实验数据以平均值±标准误差表示,利用软件SPSS Statistics26.0 进行方差分析,P<0.01表示差异性极显著P<0.05 表示差异显著,采用Origin 2021.0 进行数据和图像处理并生成图片。
2 结果与分析
2.1 单因素结果分析
2.1.1 提取次数对元宝枫种仁多糖得率的影响 提取次数对PATM 得率的影响如图1 所示,提取1 次多糖得率为17.79%,提取2 次多糖得率为20.60%,提取3 次多糖得率为21.72%。提取2 次和提取3 次多糖得率增加不显著(P>0.05)。这是由于提取1 次已经将大部分元宝枫种仁多糖提取出来,增加提取次数,PATM 得率增加不明显。考虑到能耗等因素,选择提取1 次,进行后续试验。
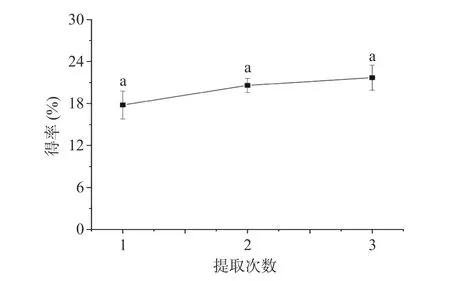
图1 提取次数对PATM 得率的影响Fig.1 Effect of extraction times on the yield of PATM
2.1.2 提取温度对多糖得率的影响 如图2 所示,随着提取温度的增加,PATM 得率呈现先增加后平稳的趋势。在80 ℃时,PATM 得率达到最大值,得率为17.97%。之后继续升高提取温度,PATM 得率也没有显著变化(P>0.05),趋于稳定。这是由于在80 ℃时能够将元宝枫种仁多糖充分提取出来,达到最大值。提取温度过高不仅能耗增加,而且会对多糖的结构造成破坏,影响后续PATM 的表征。因此,最佳提取温度确定为80 ℃。
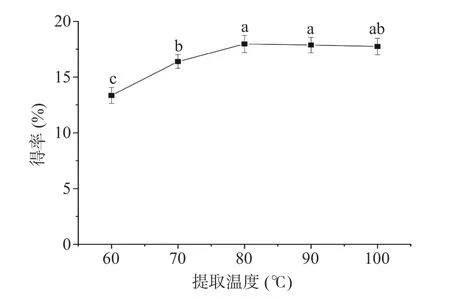
图2 提取温度对PATM 得率的影响Fig.2 Effect of extraction temperature on the yield of PATM
2.1.3 提取时间对多糖得率的影响 提取时间对PATM 得率的影响如图3 所示,提取时间在1~5 h的范围内,PATM 的得率呈现先上升后平稳的趋势。当提取时间为3 h 时,多糖得率达到最大,继续延长提取时间,多糖得率没有明显的变化,趋于稳定。这是因为经过3 h 的提取,元宝枫种仁多糖被充分提取,再继续延长提取时间也不会增加PATM 的得率。因此,最佳提取时间确定为3 h。
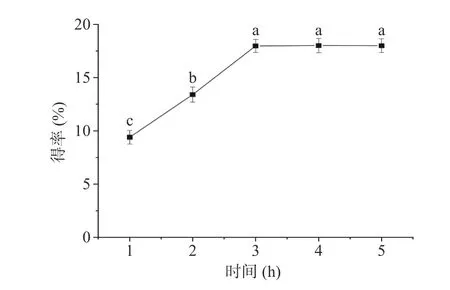
图3 提取时间对PATM 得率的影响Fig.3 Effect of extraction time on the yield of PATM
2.1.4 液固比对多糖得率的影响 如图4 所示,液固比在10:1~40:1 mL/g 的范围内,随着液固比的增加元宝枫种仁多糖得率先上升后下降,以30:1 mL/g的液固比为转折点,PATM 得率略微下降。因此,最佳液固比确定为30:1 mL/g。
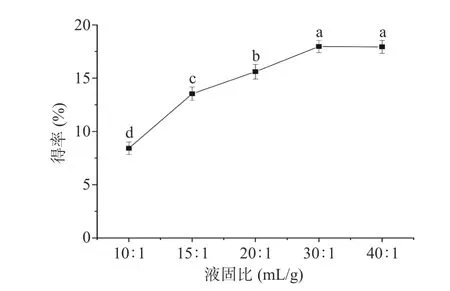
图4 液固比对PATM 得率的影响Fig.4 Effect of liquid-solid ratio on the yield of PATM
2.2 响应面优化分析
基于单因素实验结果,考虑到提取次数对PATM得率影响较小,故将提取次数固定为1 次。以提取温度、提取时间和液固比三个因素为自变量,PATM得率为响应值,通过响应面分析法对PATM 的提取工艺进行三因素三水平优化。响应面试验方案及结果见表2。通过Box-Behnken 对比实验数据进行拟合,得到PATM 得率的回归方程如下:
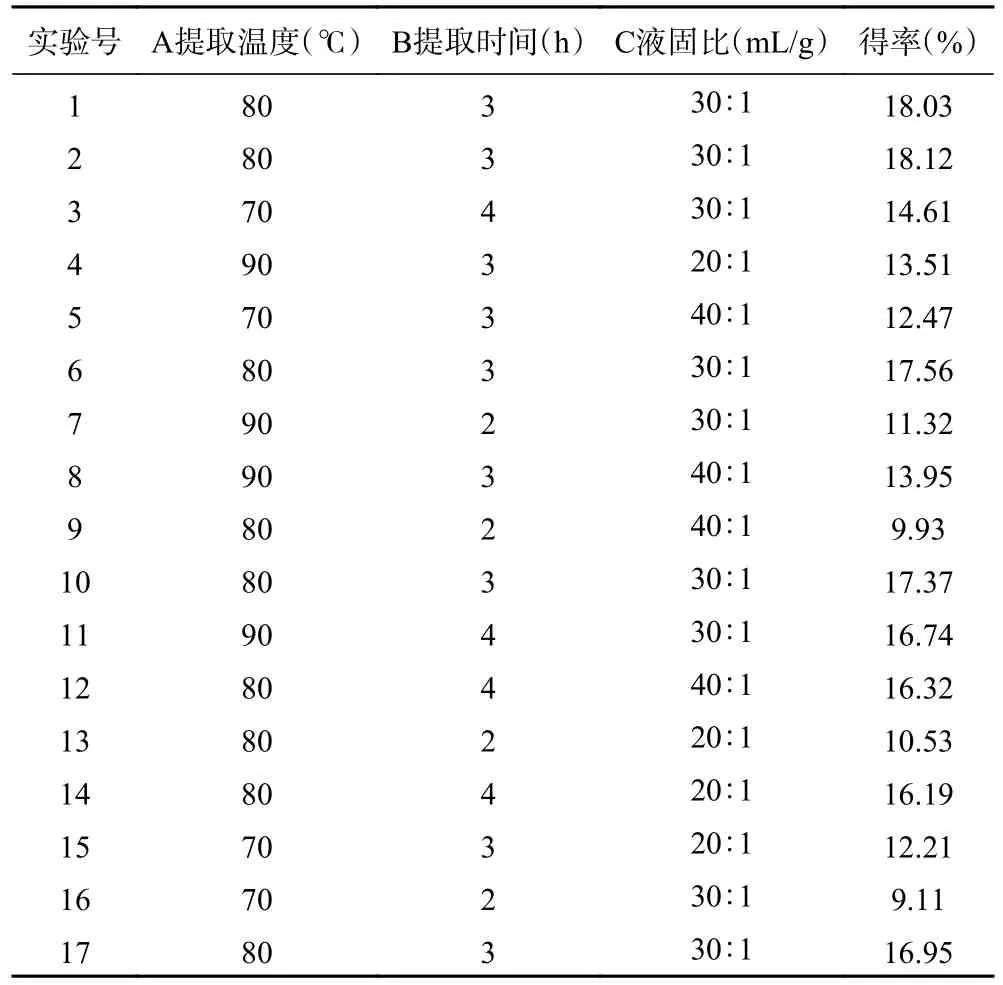
表2 响应面试验设计及试验结果Table 2 Response surface test design and test results
回归模型方差分析结果,见表3,该回归模型极其显著(P<0.0001),失拟项P=0.5120>0.05,说明失拟项差异不显著;决定系数R2=0.9893,校正后R2adj=0.9756,说明该回归方程拟合度良好,实验方法可信度较高[23],可用于PATM 得率的分析和预测。
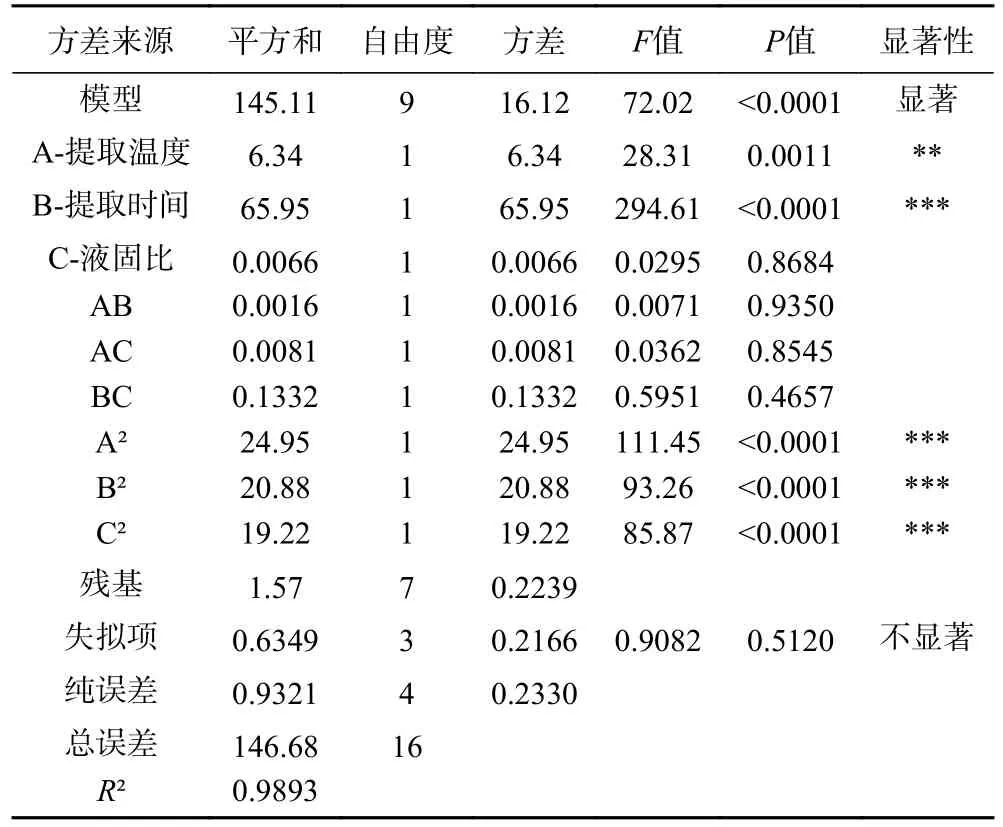
表3 回归模型及方差分析结果Table 3 Regression model and analysis of variance results
依据回归模型,可预测PATM 最佳提取工艺为:提取温度79.879 ℃,提取时间3.341 h,液固比30.259:1 mL/g,此时PATM 得率理论值为18.316%。考虑到实际可操作性,将最佳提取工艺参数调整为:提取温度80 ℃,提取时间3.5 h,液固比30:1 mL/g,进行3 次平行实验,PATM 多糖平均得率为18.17%±0.48%,这也与理论值相近,可见该模型能较好地预测实验结果。
2.3 元宝枫种仁多糖的分离纯化
2.3.1 纤维素层析柱分离纯化 如图5 所示,经DEAE-纤维素52 阴离子交换柱对PATM 进行纯化,PATM 经由NaCl 溶液洗脱,得到四个多糖组分:PATM-1(16.44%)、PATM-2(3.58%)、PATM-3(79.20%)、PATM-4(0.76%)。在比较样品中每个级别组分的百分比后,PATM-3 被认为是PATM 的主要组分,收集并用于SephadexG-100 凝胶层析柱进一步纯化。
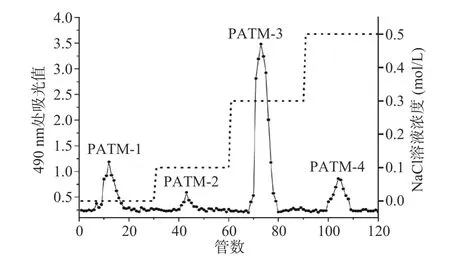
图5 DEAE-纤维素52 阴离子交换柱纯化PATMFig.5 Purification of PATM by DEAE-Cellulose 52 anion exchange column
2.3.2 凝胶层析柱分离纯化 SephadexG-100 凝胶层析柱是根据分子量大小对多糖样品进行分离纯化,大分子量的多糖分子先流出凝胶柱,而小分子量的多糖分子后流出,从而实现分离。如图6 所示,PATM-3经由SephadexG-100 凝胶层析柱分离纯化得到PATM-3-1、PATM-3-2、PATM-3-3。将PATM-3-1收集进行浓缩、透析和冻干,测定结果表明,PATM-3-1 多糖的纯度为97.17%。
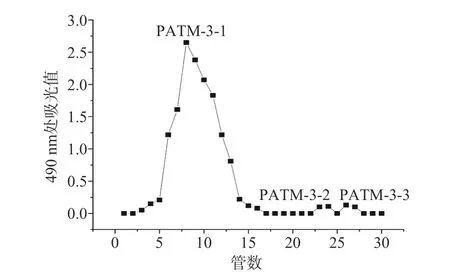
图6 SephadexG-100 凝胶柱纯化PATM-3Fig.6 Purification of PATM-3 by SephadexG-100 gel column
2.4 单糖组成分析
如图7A 所示,标准品L-岩藻糖、D-氨基半乳糖、L-鼠李糖、L-阿拉伯糖、D-氨基葡萄糖、D-半乳糖、D-葡萄糖、D-木糖、D-甘露糖、D-果糖、D-核糖、D-半乳糖醛酸、L-古罗糖醛酸、D-葡萄糖醛酸和D-甘露糖醛酸出峰时间依次为3.68、7.05、7.45、7.75、8.91、9.77、11.25、13.19、13.85、15.88、17.27、34.17、34.96、36.68、39.03 min,峰形匀称且分离度较好,无干扰现象,可用于多糖组成分析。通过与标准品对比,PATM-3-1(图7B)是由D-半乳糖醛酸、L-阿拉伯糖、D-半乳糖、L-鼠李糖、D-葡萄糖、D-木糖、D-甘露糖、D-葡萄糖醛酸、L-岩藻糖、D-核糖和D-氨基葡萄糖组成,其摩尔质量之比为12.40:12.81:5.08:3.86:2.69:2.31:1:0.66:0.32:0.06:0.02,说 明PTAM-3-1 为酸性杂多糖,主要以D-半乳糖醛酸和L-阿拉伯糖两种单糖为主。
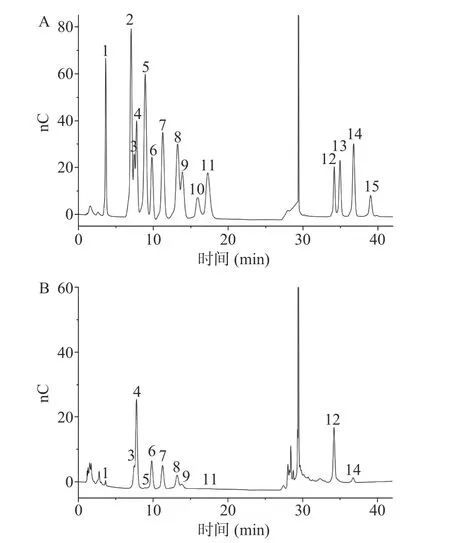
图7 单糖标准品(A)和PATM-3-1(B)的色谱图Fig.7 Chromatograms of monosaccharide standard (A) and PATM-3-1 (B)
2.5 红外光谱分析
FT-IR 光谱是阐明多糖特征官能团的重要工具之一,PATM-3-1 的红外光谱表现出多糖的典型吸收峰(图8)。在3283 cm-1处出现强而宽的特征峰,归因于O-H 拉伸振动;在2928 cm-1处的弱吸收峰归因于C-H 拉伸振动[24];1740 cm-1左右的吸收峰归因于-OH 羧酸羰基的拉伸振动[25],这表明多糖结构中存在糖醛酸;1643 cm-1处的条带归因于-OH 的弯曲振动;1600 cm-1处的条带归因于糖醛酸的CO 拉伸振动[26];1411 和1240 cm-1处的峰是C-H 的可变角度振动,可以判断为多糖;1077 和1013 cm-1之间的特征吸收峰归因于吡喃糖环的拉伸振动[27-28];767 cm-1处的吸收峰则表明该多糖为β构型吡喃糖。
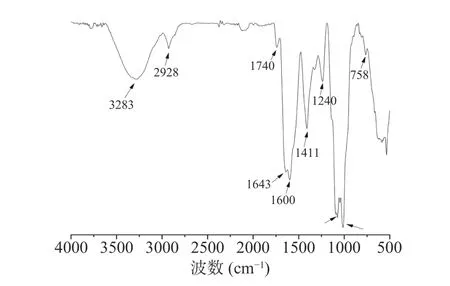
图8 PATM-3-1 红外光谱图Fig.8 Infrared spectrogram of PATM-3-1
2.6 热重分析
PATM-3-1 的TG 和DTG 曲线见图9,其中存在两个显著的热损失:第一个是由于水分蒸发,多糖从40 ℃到126.27 ℃损失了8.80%的质量,这一结果表明多糖具有一定的持水能力;第二个是多糖热分解产生的,从221.25 ℃至443.73 ℃损失了54.90%的质量,在260.70 ℃达到最快的热分解速率。
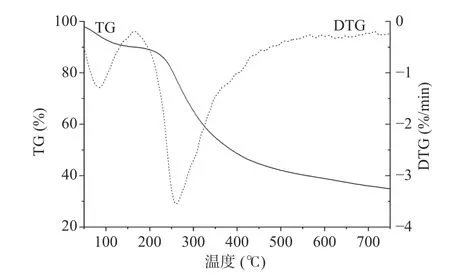
图9 PATM-3-1 的TG-DTG 曲线Fig.9 TG-DTG curve of PATM-3-1
2.7 刚果红分析
刚果红能与三股螺旋结构的多糖结合在一起形成络合物,从而使其最大吸收波长发生改变。实验结果如图10 所示,元宝枫种仁多糖PATM-3-1 与刚果红络合物的最大吸收波长在0~0.4 mol/L NaOH 浓度下向长波长方向偏移,在0.4~0.5 mol/L NaOH 浓度下最大吸收波长明显下降,证明元宝枫种仁多糖PATM-3-1 具备三股螺旋结构[29]。
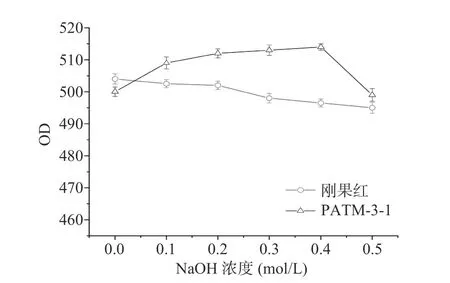
图10 PATM-3-1 在不同 NaOH 浓度下最大吸收波长变化Fig.10 Maximum absorption wavelength of PATM-3-1 under different NaOH concentrations
2.8 体外降血糖活性分析
2.8.1α-淀粉酶抑制活性α-淀粉酶抑制剂可以与体内的α-淀粉酶发生作用,阻止淀粉转化为葡萄糖,可以有效控制血糖水平[30]。因此,寻找α-淀粉酶的天然抑制剂是目前国内外降糖的研究热点之一。在本研究中,不同质量浓度的PATM-3-1 对α-淀粉酶抑制活性见图11,PATM-3-1 在0.3~5 mg/mL 浓度范围内表现出对α-淀粉酶的抑制率呈现剂量依赖关系,PATM-3-1 和阿卡波糖α-淀粉酶的半抑制质量浓度IC50值分别为2.04 和0.032 mg/mL。宋田源等[31]的研究发现,红毛藻多糖对α-淀粉酶抑制随着浓度增加而增强,其IC50值为1.26 mg/mL,PATM-3-1对α-淀粉酶的半数抑制浓度与红毛藻多糖相接近,由此可以看出PATM-3-1 展现出良好的α-淀粉酶抑制活性。
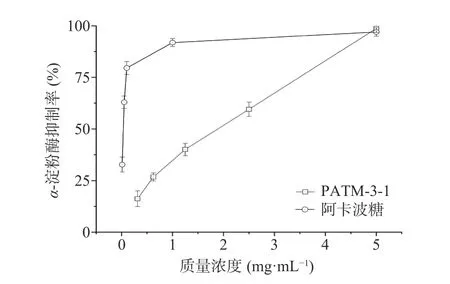
图11 不同质量浓度的PATM-3-1 对α-淀粉酶的抑制率Fig.11 α-Amylase inhibition rate of different mass concentration of PATM-3-1
2.8.2 胰岛素抵抗模型 目前认为胰岛素抵抗(insulin resistance,IR)不仅是2 型糖尿病的发病基础,也是贯穿多种代谢相关疾病的主线,是心血管疾病、高脂血症、高尿酸血症及代谢综合症的共同病理基础。构建胰岛素抵抗细胞模型是进行IR 相关疾病治疗研究及药物研发的有效途径[32]。通过含有不同浓度的胰岛素(10-5~10-9mol/L)的MEM 培养基培养36 h,诱导HepG2 细胞产生胰岛素抵抗(图12),模型组与正常组相比,葡萄糖的消耗量明显降低,显著低于正常组(P<0.05),说明胰岛素抵抗模型建立成功。其中胰岛素浓度为10-7mol/L 时,其葡萄糖的消耗量最低,显著低于其他浓度模型组(P<0.05)。因此,选择该浓度的胰岛素来构建胰岛素抵抗的HepG2细胞模型,用于进行后续实验。
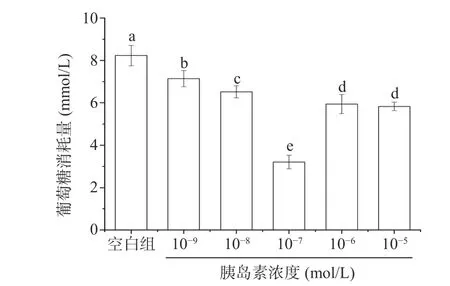
图12 胰岛素抵抗HepG2 细胞模型Fig.12 Insulin-resistant HepG2 cell model (IR-HepG2)
2.8.3 细胞毒性 采用MTT 法检测PATM-3-1 对HepG2 细胞毒性,并依据HepG2 细胞存活率筛选出后续实验所使用PATM-3-1 的安全剂量。不同质量浓度的PATM-3-1 对HepG2 细胞干预24 h 后的细胞存活情况见图13。可以看出在80~2560 μg/mL的浓度范围内,PATM-3-1 对HepG2 细胞无明显的抗增殖作用,这表明元宝枫种仁多糖PATM-3-1 对HepG2 细胞无毒性,80~2560 μg/mL 可以作为后续实验PATM-3-1 的安全浓度范围。
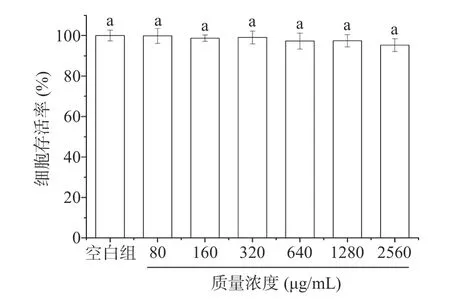
图13 不同质量浓度的PATM-3-1 对HepG2 细胞存活率的影响Fig.13 Effect of different mass concentration of PATM-3-1 on the survival rate of HepG2 cells
2.8.4 葡萄糖消耗量和糖原含量分析 在正常机体内细胞利用胰岛素将葡萄糖分解为机体提供能量。当机体产生胰岛素抵抗时,细胞对胰岛素敏感性降低,导致细胞对葡萄糖摄取量下降[33]。使用改善胰岛素抵抗的药物会提高细胞对葡萄糖的消耗量[34]。不同质量浓度的PATM-3-1 对胰岛素抵抗的HepG2 细胞(Insulin Resistance-HepG2,IR-HepG2)葡萄糖消耗量的影响见图14A。空白组、模型组、阳性对照组、高剂量组、中剂量组和低剂量组细胞的葡萄糖消耗量分别为8.63、2.86、8.69、8.55、7.53 和7.03 mmol/L。与空白组细胞相比,模型组细胞的葡萄糖消耗量显著降低(P<0.05),降低约为66.86%;与模型组相比,样品组和阳性对照组细胞的葡萄糖消耗量显著提高(P<0.05),并且高剂量组、中剂量组和低剂量组细胞的葡萄糖消耗量与PATM-3-1 呈现剂量依赖关系。空白组、高剂量组和阳性对照组细胞的葡萄糖消耗量无明显差异,这说明PATM-3-1 能有效调节IR-HepG2 细胞对葡萄糖的吸收,高剂量PATM-3-1作用与阳性药相当。
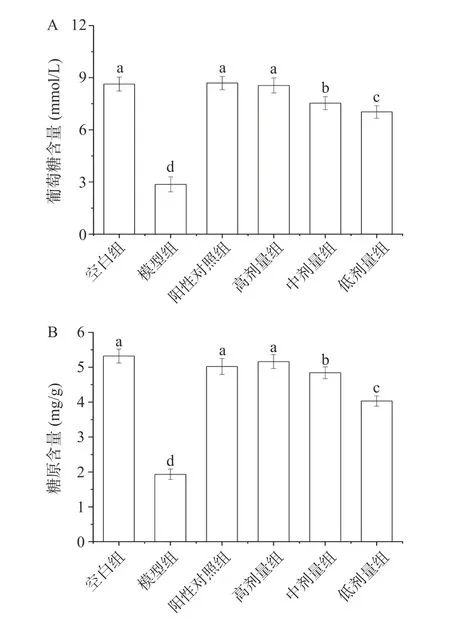
图14 不同质量浓度的PATM-3-1 对IR-HepG2 细胞葡萄糖含量(A)和糖原含量(B)的影响Fig.14 Effects of different mass concentration of PATM-3-1 on glucose content (A) and glycogen content (B) in IR-HepG2 cells
此外,细胞摄取葡萄糖经各种酶作用合成糖原,糖原的合成与分解对维持血糖平衡极为重要。不同质量浓度的PATM-3-1 对IR-HepG2 细胞合成糖原的影响见图14B,其中空白组、模型组、阳性对照组、高剂量组、中剂量组和低剂量组的糖原含量分别为5.32、1.93、5.02、5.16、4.84 和4.03 mg/g。与空白组相比,模型组细胞的糖原含量显著降低(P<0.05),降低约为63.72%。与模型组相比,经由PATM-3-1 和阳性药(二甲双胍)处理后的IRHepG2 细胞的糖原含量得到显著提高(P<0.05),这表明PATM-3-1能发挥改善IR-HepG2 细胞糖代谢紊乱的作用。
2.8.5 己糖激酶(HK)和丙酮酸激酶(PK)活性分析己糖激酶(HK)被认为是糖酵解途径中催化己糖磷酸化为磷酸己糖的一种重要的酶。丙酮酸激酶(PK)是糖酵解中的另一种重要酶,胰岛素抵抗会降低己糖激酶和丙酮酸激酶的活性。因此,调节HK 和PK 活性是促进葡萄糖代谢的关键因素[35]。如图15A 和图15B 所示,空白组、模型组、阳性对照组、高剂量组、中剂量组和低剂量组细胞的HK 活性分别为16.18、7.32、15.17、15.91、13.57 和11.93 nmol/min/mg prot;细胞的PK 活性分别为74.93、52.31、73.30、72.66、68.73 和67.33 U/g prot。与空白组相比,模型组细胞的HK 和PK 活性显著降低(P<0.05);而阳性对照组细胞的HK 和PK 活性显著升高。与模型组相比,经PATM-3-1 处理后,细胞的HK 和PK 活性显著提高(P<0.05),呈现与PATM-3-1 剂量依赖关系。这说明PATM-3-1 具有增强IR-HepG2 细胞HK和PK 活性,促进糖代谢,发挥降糖作用。
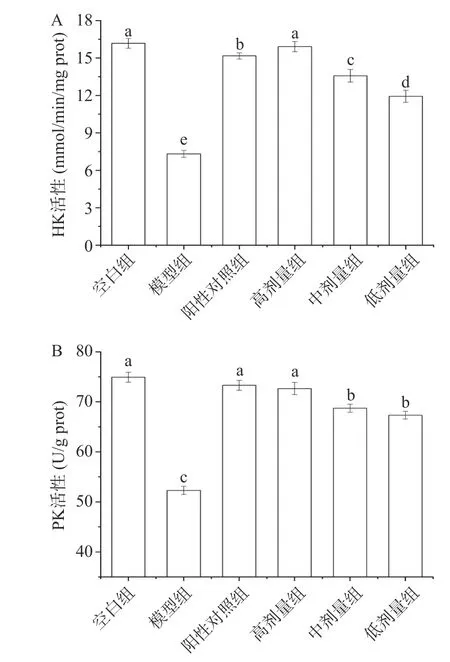
图15 不同质量浓度的PATM-3-1 对IR-HepG2 细胞HK活性(A)和PK 活性(B)的影响Fig.15 Effects of different mass concentration of PATM-3-1 on HK activity (A) and PK activity (B) in IR-HepG2 cells
3 结论
本研究通过单因素和响应面试验优化了元宝枫种仁多糖的提取工艺,在最佳提取工艺条件下,多糖平均得率为18.17%,通过脱蛋白、DEAE-DE 纤维素52 层析柱和SephadexG-100 凝胶层析柱分离纯化得到不具有三螺旋结构的多糖PATM-3-1,其纯度为97.17%,它是由D-半乳糖醛酸、L-阿拉伯糖、D-半乳糖、L-鼠李糖、D-葡萄糖、D-木糖、D-甘露糖、D-葡萄糖醛酸、L-岩藻糖、D-核糖和D-氨基葡萄糖单糖组成。PATM-3-1 属于β构型吡喃糖,具有良好的热稳定性和降糖活性,能够有效调节IR-HepG2 细胞对葡萄糖吸收和糖原合成,增强HK 和PK 活性,从而改善IR-HepG2 细胞的糖代谢。PATM-3-1 展现出良好的降血糖活性,为植物元宝枫种仁多糖在药物和功能性食品中的研发、提取和应用奠定了理论基础。
© The Author(s) 2024.This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
河北果树(2021年2期)2021-04-24
中国生物防治学报(2021年1期)2021-04-18
经济林研究(2021年1期)2021-04-10
中国粮油学报(2019年4期)2019-07-12
山东林业科技(2019年2期)2019-06-03
吉林农业(2019年9期)2019-01-06
中国粮油学报(2018年12期)2018-03-19
新疆农业科学(2017年4期)2017-06-08
农村.农业.农民(2016年24期)2016-04-02
食品工业科技(2014年15期)2014-03-11
