
喀斯特栎类老龄林的空间结构分析
2024-04-14 12:51骆秀琼何吉安李远发
林业科学研究 2024年2期
骆秀琼,何吉安,李远发,2*
(1. 广西大学林学院,广西森林生态与保育重点实验室,广西 南宁 530004;2. 来宾金秀大瑶山森林生态系统广西野外科学观测研究站,广西 来宾 545700)
喀斯特森林深受生物(如,竞争、种子扩散限制、负密度制约等)和非生物因子(如,生境过滤和人为干扰等)的双重影响[1],它们在区域、地区水平上的类型相差很大。有关喀斯特原始林或老龄林的物种组分和结构的研究正处于快速发展中,主要聚焦在群落和林分水平,包括活立木种群或是同属物种的分布格局和空间关联[2-5]、生境或立地类型对生物多样性和活立木分布格局的影响[6-7]、地上和地下生物量的分配模式[8]以及枯死木的分布格局[9]。这些研究侧重于分析喀斯特森林的水平结构,但很少涉及垂直结构。
中国西南地区是全球喀斯特发育的典范和三大集中区域之一[10-11]。由于人口密度大、开荒和放牧等诸多原因,该区域的大部分森林曾遭受过严重破坏,甚至产生石漠化现象,仅在一些偏远地区或自然保护区留存有少量老龄林或原始林[8-9]。其中,南盘江流域的喀斯特老龄林包含多个栎类树种[6-7,12-13]。栎类树种广泛分布于全球其他国家和地区[14-15]。一些栎类林分呈现出明显的垂直分层现象[6],即数种栎类(如,栓皮栎(Quercus variabilisBlume)、白栎(Quercus fabriHance))占据林分上层,珍珠花属(LyoniaNutt.)的灌木以及一些小乔木和更新占据了林分下层。这些喀斯特老龄林的垂直分层缺乏量化研究,形成机理也尚不清楚。垂直分层作为喀斯森林的重要结构特征,对探究生境——物种关联以及当地植被恢复与重建、石漠化的治理十分重要。
1 材料与方法
1.1 试验地概况与样地建立
试验区域坐落于广西雅长兰科植物国家级自然保护区(106°11′31″~106°27′04″ E, 24°44′16″~24°53′58″ N)的花坪保护站,属于中国西南地区的滇、黔、桂3 省接壤之地,也是云贵高原向广西丘陵地过渡的山原地带。该保护站年均气温16.3 ℃,极端高温可达41.1 ℃,极端低温为-3 ℃,冬季时有霜雪,年均日照1 467.0 h,年均降水量1 051.7 mm,降水集中在夏季,春秋两季干旱少雨。植被以天然次生林为主,一些高山区域存在数量极少的老龄林(最老树龄 > 300 a),郁闭度在90%以上[6-7]。栎类树种在老龄林占据绝对优势,常见伴生种包括化香树(Platycarya strobilaceaSieb. et Zucc.)、铁坚油杉(Keteleeria davidiana(Bertr.)Beissn.)、盐肤木(Rhus chinensisMill.)、西南桦(Betula alnoidesBuch.-Ham. ex D. Don)、枫香(Liquidambar formosanaHance)、灯台树(Cornus controversaHemsley)等。灌木层以珍珠花属(珍珠花(Lyonia ovalifolia(Wall.)Drude)、毛叶珍珠花(Lyonia villosa(Wall. ex C. B. Clarke)Hand.-Mazz.))为优势种,其伴生树种主要有水红木(Viburnum cylindricumBuch.-Ham. ex D. Don)、 大叶紫珠(Callicarpa macrophyllaVahl )、 猴耳环(Archidendron clypearia(Jack)I. C. Nielsen)等。草本植物盖度低,以五节芒(Miscanthus floridulus(Lab.)Warb. ex Schum et Laut.)和数种小蕨类为主。藤本植物主要由粗叶悬钩子(Rubus alceaefoliusPoiret)、插田泡(Rubus coreanusMiq.)、爬山虎(Parthenocissus tricuspidata(Siebold & Zucc.)Planch.)、 金花忍冬(Lonicera chrysanthaTurcz.)构成,但数量不多。
本研究根据CTFS(Center for Tropical Forest Science)样地建设标准,在喀斯特栎类老龄林中建立了一个面积为200 m × 110 m 的样地[4](图1)。首先,用全站仪(南方测绘公司,型号NTS-372R10)沿着坡面水平方向定位样地的第一条边界(200 m),再逆时针旋转机器确定剩余3 条边界,形成闭合差小于1/400 的样地。其次,用全站仪将样地划分为 220 个10 m × 10 m 的小样方,在每个小样方的顶点钉入60 cm 长的PVC 管,用螺纹钢加固作为标记,并在各个标记之间用塑料红绳连接划分小样方边界。最后,逐一定位每个小样方内所有活树(树高1.3 m,DBH ≥ 1 cm)的相对坐标(x,y,z),检尺活立木的胸径、树高。为每株DBH ≥ 5 cm 的活立木钉上带有唯一编号的铝牌,为幼树和幼苗挂上带编号的塑料红牌。该样地共有4 596 株活立木,分别属于62 种(表1)。
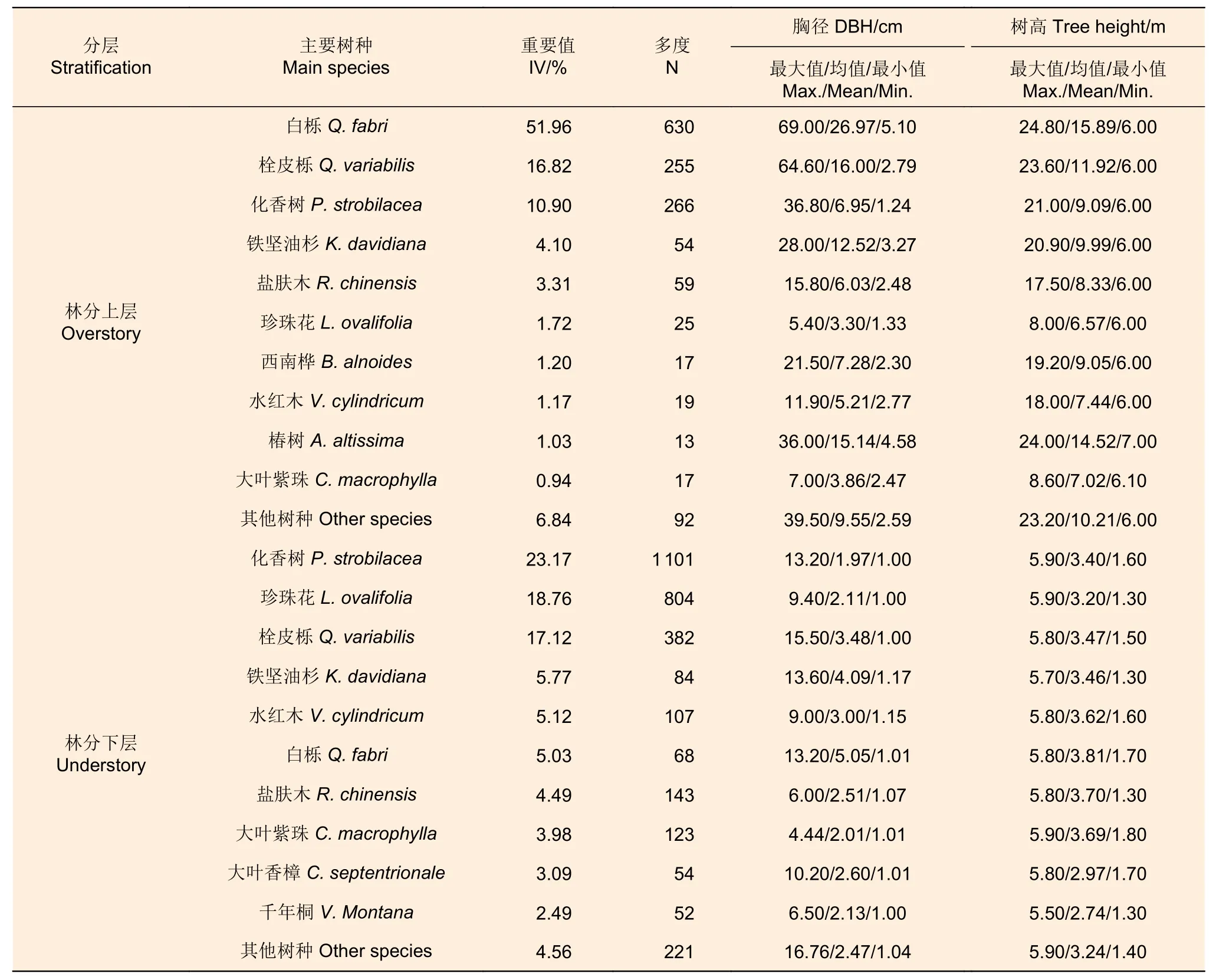
表1 栎类老龄林的分层属性 Table 1 Vertical attributes of old growth oak forest
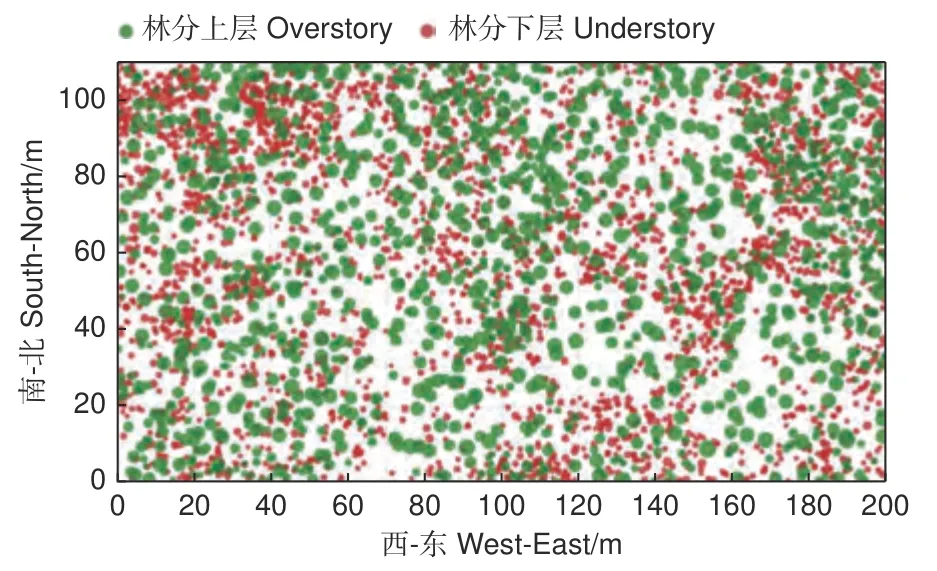
图1 树木点分布Fig. 1 distribution of tree points
1.2 数据分析
1.2.1 分层及优势种的确定 树高和生活型(即乔木或灌木)是划分垂直结构的重要依据[12-13,19]。树高分布图显示6 m 可作为林分上层和林分下层的界限(图2e)。用负指数函数拟合了林分、林分上层和林分下层的径阶分布,用重要值(IV)(式1)计算每个树种在分层中的重要性,并选择重要值排名前3 的树种作为该林层的优势种。林分上层的优势种为白栎、栓皮栎和化香树,林分下层的优势种为化香树、珍珠花和栓皮栎。这些树种的重要值总和高于50%,多度(N)也明显大于其他树种,较好地反映林分、林分上层和林分下层的物种构成基本情况(表1)。
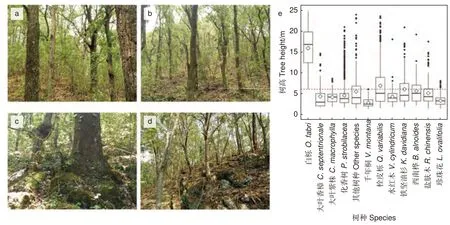
图2 树高分布Fig. 2 tree height distribution
式中,相对频度是指某一物种的频度与所有物种频度总和的百分比,相对多度是指某一物种的多度与所有物种多度的百分比,相对胸高断面积指某一物种的胸高断面积占样地面积的百分比。
1.2.2 空间格局分析 空间格局用成对相关函数g(r) 分析(式2)。g(r) 量化了样方内以任一点为圆心、半径为r的圆环区域上成对树木之间的距离关系,消除了K(r) 函数(式3)的统计积累误差效应,有效解决物种分布对尺度的依赖难题,很好地解释了树木点分布随尺度的变化,备受推崇[10,15]。因试验样地生境异质性十分突出(图2a~d),零模型使用泊松异质性零模型(HP),用g(r) 的单变量函数g11(r) 分析林分上层、林分下层和各层优势种的分布格局。当观测值落在包迹线之内、之上和之下分别表示随机、聚集和规则分布;用g(r)的双变量函数g12(r) 分析层间、层间优势种和层内优势种的空间关联。当观测值落在包迹线之内、之上和之下分别表示无关联或随机关联、正关联和负关联[10,15]。
其中,N为株数,r为样圆半径,wij为树木i与树木j之间的权重,A为样地面积。
1.2.3 标记特征分析 任意两株树木之间的种类和个体大小随观测尺度的变化用基于点过程的标记相关函数(MCF)kf(r) 分析(式5),即分析了距离r上第i和第j两株相邻木标记特征的空间自相关性。其原理是采用了一个具有2 参数f(m1,m2)的检验函数f分析距离r上2 个标记点之间的乘积或类别与对应期望值之间的差异。公式5 中,Eij为点过程中相距r的第i和j株树的条件期望值,m(i) 和m(j) 为第i,j株树上的属性,m和m'是独立地从标记的边缘分布中抽取的随机标记。当标记点为定性属性(例如,树种)时,f(m1,m2) =1(m1=m2)。若观测值落在包迹线之内,表示种间相互独立,即任何一株树木i的相邻木j是其同种或异种的概率相等;若观测值落在包迹线之上,表示同种聚集或异种排斥;若观测值落在包迹线之下,表示异种聚集或同种排斥。当标记点为定量属性(例如,胸径)时,f(m1,m2) =m1×m2。若观测值落在包迹线之内,表示尺度r上树木大小相互独立,即不同大小的树木混生;若观测值落在包迹线之上,表示两棵树的胸径之间成正相关;若观测值落在包迹线之下,则认为两棵树的胸径之间成负相关,即尺度r上两棵树的胸径乘积小于全部树木胸径的均值[16]。
空间格局和标记特征的分析过程均用蒙特卡罗(MC)方法随机模拟任意观测尺度上的观测值199 次,产生置信度为 95% 的包迹线,以判断观测值偏离理论值的显著程度。为了便于统计,在分析空间格局和标记特征时,将r的等间距离设为0.001 m;分析空间关联时,将r的等间距离设为1 m。数据分析和图形处理均在R 语言(https://www.R-project.org)的spatstat 数据包中完成。
2 结果与分析
2.1 物种多度和丰富度的分布特征
总体上,林分、林分上层和林分下层的径阶分布都呈倒J 型,即多度随胸径的增大而减小,并逐渐趋于平缓(图3a~c)。稍有不同的是,林分上层多度的变化比林分和林分下层的更平缓,且缺乏部分小径木。相似的情形也出现在物种丰富度的分布中,即林分的物种丰富度随胸径的增大而减小,当径阶为1~3 cm 时有最大值,为51 种(图3a)。林分上层的物种丰富度呈先上升后下降的变化趋势,在径阶7~9 cm 时达最大值24,随后减小至0(图3b)。林分下层的物种丰富度变化与林分的相似,即在径阶为1~3 cm 时为最大值49,随后减小至0(图3c)。
其实古意的心思我一直都很明白,他恼恨的倒不是二乐,而是我借着二乐来打扰他的好事。从我有记忆起,古意就一直在做对不起康美娜的事情,他频频与女人约会,不停地出入情场。
2.2 分层及优势种的分布格局
林分上层仅在小尺度(r= 0~0.5 m、4~6 m)上呈现聚集分布,而在多数尺度(r= 0.5~4 m、6~50 m)上为规则和随机分布(图4a)。优势种白栎在所有尺度上都是随机分布(图4b)。化香树在r= 0~8 m 为聚集分布,在r= 8~10 m、16~18 m 为随机分布,而在其他尺度上为规则分布(图4c)。栓皮栎仅在r= 0~1 m 上为聚集分布,在r= 1~12 m、13~19 m、23~32 m、33~45 m 上为随机分布,而在其他尺度上为规则分布(图4d)。林分下层在小尺度(r= 0~7.5 m)上为聚集分布,且聚集强度随尺度的增大而减小,在r= 7.5~46 m 为随机分布,随后转为规则分布(图4e)。优势种化香树、珍珠花和栓皮栎的分布格局变化趋势相同,即随尺度的增大由聚集分布过渡为随机分布,随后转变为规则分布(图4f~h)。不同的是栓皮栎的聚集分布尺度(r= 0~34 m)和随机分布尺度(r= 35~48 m)大于化香树和珍珠花。
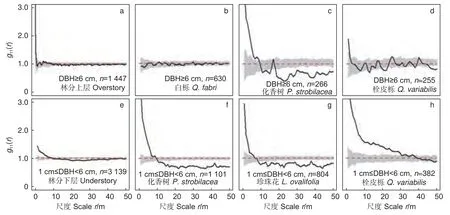
图4 林分上层、林分下层以及各自主要树种的空间分布Fig. 4 Spatial distributions of overstory, understory and their main populations
2.3 层间、层内主要树种的空间关联
林分上层和林分下层之间的关系以随机关联为主,仅在部分小尺度(r= 3~4 m)上为正关联(图5a)。它们的主要种群也以随机关联为主,只有个别种间对在小尺度上呈现为正关联或负关联(图5b~j)。除了林分上层的白栎与林分下层的化香树在小尺度r= 0~2 m 上呈现为负关联,层间栓皮栎在尺度r= 2~3 m 上为正关联,而层间其他树种之间为随机关联。林分上层主要树种之间以随机关联为主,仅有白栎和化香树在小尺度r=0~2 m、白栎和栓皮栎在小尺度r= 0~1 m 上表现为负关联(图5k~m)。林分下层主要树种之间在所有尺度上均为随机关联(图5n~p)。
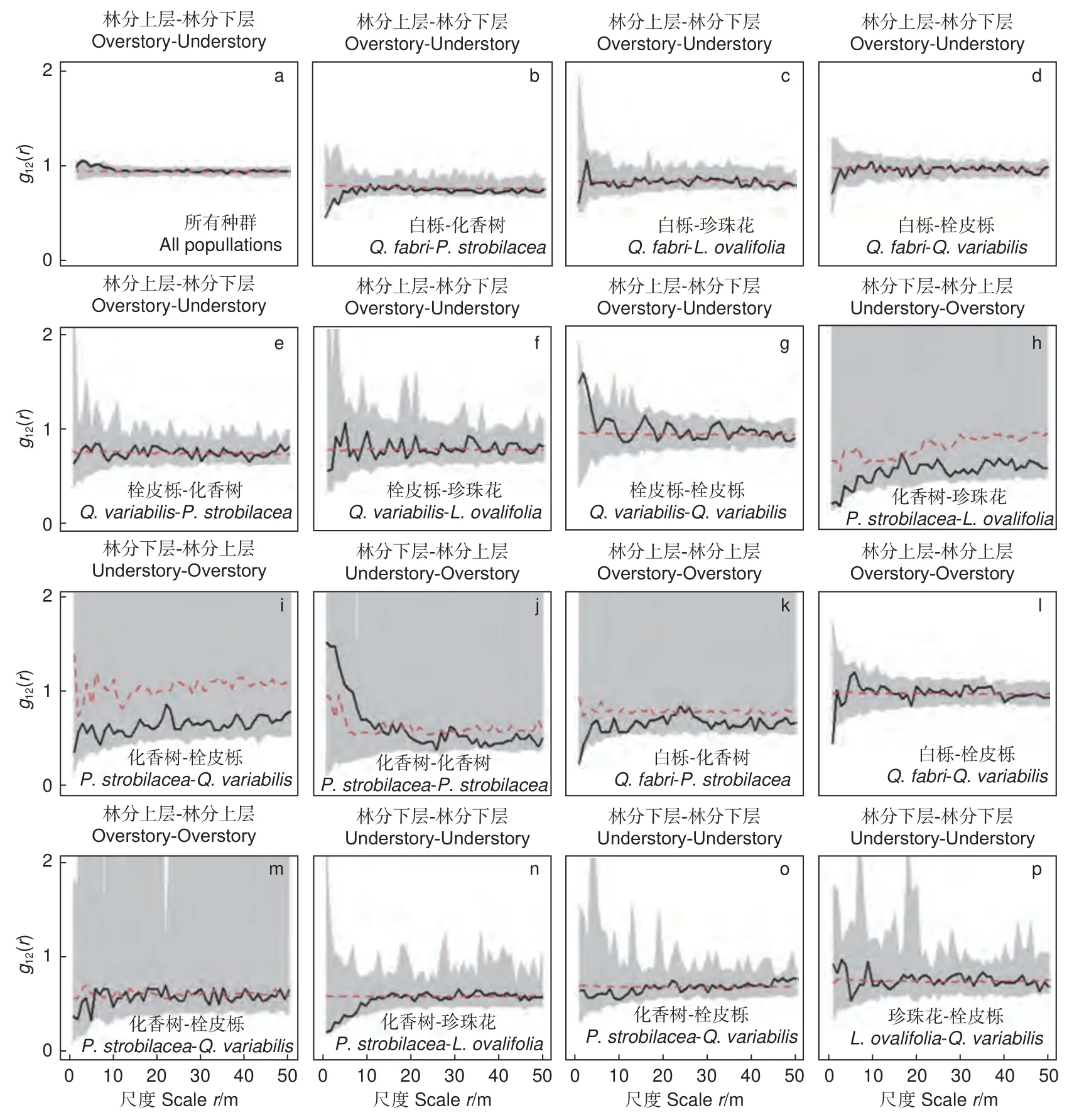
图5 层间和层内的空间关联Fig. 5 Spatial associations of main populations in and among overstory and understory
2.4 林分及分层的树种和胸径的分布
林分的树种在每个观测尺度上均表现为同种聚集(图6a)。林分上层的树种在尺度r= 0~21 m上为同种聚集,在尺度r= 21~30 m 上种间相互独(图6b)。林分下层的树种分布与林分高度相似(图6c)。在林分水平上,小树在r= 0~25 m 范围内相互聚集,不同大小的树木在r= 25~38.5 m上混生,而大树在r= 38.5~50 m 上聚集(图6d)。林分上层的树木大小展现出与林分相似的特征(图6e)。相比之下,林分下层的观测值在r=0~16 m 上更接近95% MC 模拟区间,并在大范围内(r= 16~50 m)呈不同大小树木混生的状态(图6f)。
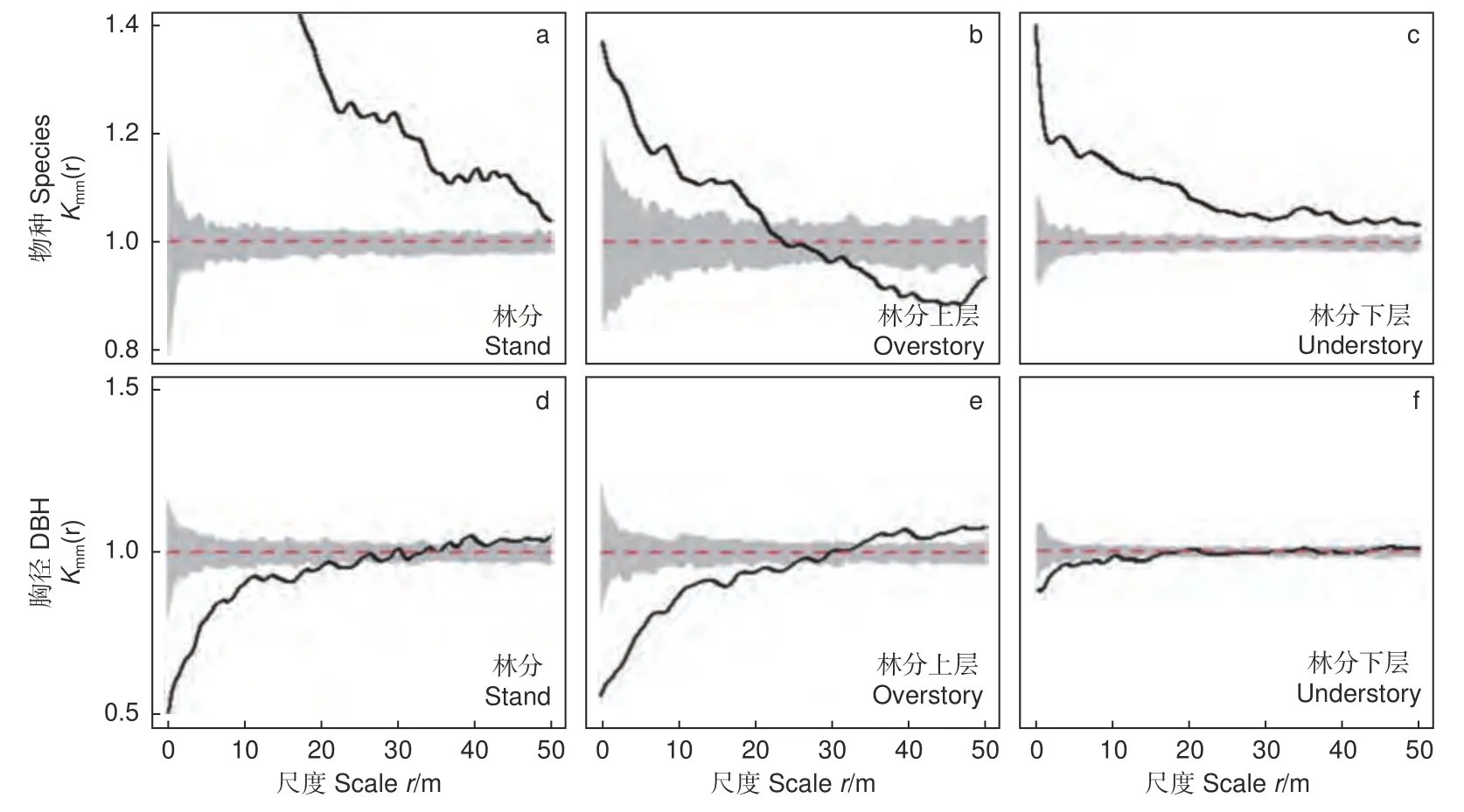
图6 林分、林分上层、林分下层的树种和胸径标记特征Fig. 6 Mark character of tree species and DBH of the stand, overstory and understory
3 讨论
3.1 喀斯特森林的基础结构特征
径级结构是物种对生境长期适应的结果,综合反映群落受干扰状况、稳定性以及演替发展的趋势[14]。中国西南地区南盘江流域喀斯特栎类老龄林的径阶分布呈倒J 型,其q值远大于前人提出的理想值1.2~1.5 或1.3~1.7[20],说明林分下层个体数量异常丰富。林分下层的径阶分布曲线陡峭,亦反映小树株数多,常表示更新充足[21-22]。本研究中,尽管样地存在不少乔木树种的更新,但林分下层主要由乔木的小树和小乔木组成(表1)。相反,林分上层的径阶分布平缓很多,分布范围较宽,径阶变异大,其q值小于理想值。显然,除了径阶距之外[23],q值的大小还取决于分析群体,这在以前的研究中尚未提及。胸径的倒J 型分布可能是个体增大,种间竞争加强的结果[24]。总体上,该喀斯特林分的径阶结构与同一地区的其他喀斯特常绿落叶阔叶混交林[10,21-22]、次生林[5]以及非喀斯特地区的栎类阔叶林[25]、栎类天然混交林[14]相似,共同反映了区域林分的生长特征,并且这种特性似乎不受物种组分、多样性和生境的影响。尽管林分径阶分布已经有了许多的研究,但很少涉及生境。
林分下层的物种丰富度和多度均高于林分上层,接近林分的数量,支持假设1,也与邻近松栎混交林[12-13]和亚热带其他分层研究结果相似[26]。林冠层结构使林分下层形成复杂的光影空间[27],另加上突出的喀斯特异质性[1,7],共同为林下物种的生存提供了大量的微生境。但一些生境缺乏足够多的基质和水分,不能支撑大树生存,只适合小树生长[4]。负密度制约也可能是导致林分下层物种比林分上层更加丰富的另一个原因[24,28]。它主要发生在小树阶段,促进多物种共存。相反,大树阶段的负密度制约可能不明显,体现在丰富度随径阶增加而缓慢减少。然而,一些热带岛屿上,森林物种丰富度在垂直方向的分布与本研究结果相反[29],暗示陆地与岛屿上森林物种多样性和结构之间的关系不同,毕竟它们的演替驱动力不同。本研究还发现DBH = 1~3 cm 的小树的丰富度远低于相邻径阶,该处是林分上层和林分下层在物种组分上的分界,可能由少量乔木的小树引起,反应了林分在垂直方向上不存在绝对的分层界线,而是一种相对的过渡状态,与林地实际情况相符。
3.2 喀斯特森林水平分布格局对分层的响应
在排除生境异质性以后,林分上层以随机分布为主,暗示竞争较小,与相邻区域的松栎混交林研究结果相似[13]。随机分布常见于林分上层,而少见于林分下层,被认为是树木竞争的结果[15,30]。种间或种内对光照、水分和营养等生境资源的竞争引发自疏,掩盖了由种子分布限制和更新产生的格局[15,31],使分布格局趋向于非聚集性分布。同一种群不同生活史的空间格局亦表明,大径木比中小径木更接近随机分布[30,32]。林分下层及其3 个主要种群在中小尺度上呈聚集分布,支持假设1,可由许多因素导致,例如生境偏好、生境异质性、生境过滤、外源干扰以及动物取食[11-13]。小乔木的有限植株高度及乔木的大粒种子重力进一步限制了种子传播距离,加强林分下层的聚集强度[33-34]。除此之外,当前异质性模型无法排除的微生境也可能影响林分下层物种组分与分布[35]。优势种群的分布格局可能完全不同,这种现象广泛发生[11,36],但很少有人关注其差异形成的原因。本研究中林分上层的化香树在小尺度上呈聚集分布,而白栎和栓皮栎呈随机分布或以随机分布为主,或许是种间的发育阶段、个体数量、种间关系、生理和生长特性以及环境的适应能力差异造成[27-28],可能利于物种共存,提高群落稳定性。
3.3 喀斯特森林种间关联的分层响应
林分上层与林分下层、层间主要树种均以随机关联为主,说明层间竞争和依赖不明显,支持假设2,与温带、亚热带、中亚热带森林的分层研究相符[12,33,36]。林分上层和林分下层的树种对光照的依赖不同[27],有效避免了直接竞争,可能是它们保持随机关联的主因。欧洲橡木(Quercuspetraea)和水青冈(Fagussylvatica)混交林有类似现象[15]。此外,垂直分层减少种间竞争[12,27],不同的分层方法亦有可能影响层间的物种关系[36]。林分上层优势种以随机关联为主,只有白栎与化香树、栓皮栎在小尺度上为负关联,表明种间存在微弱的竞争关系。林分演替到后期,生态位分化减弱种间竞争和依赖[17,24],暗示林分上层主要种群可能长期保存。林分下层主要树种之间在所有观测尺度上均为随机关联,与Li 等人[13]在同区域松栎次生林的分层研究结果相似。这可能与它们各自种内聚集程度有关,种内聚集意味着种间分离,产生交互作用的概率降低[2-7]。在中国东北长白山阔叶红松混交林中,同属树种呈聚集分布,无论种间还是它们不同大小个体几乎都呈随机关联[34],与本研究的结果一致。种间关联向随机方向发展或保持随机关系是自然界植被群落演替的重要趋势[17],也体现在喀斯特栎类老龄林中。
3.4 喀斯特森林树种和大小的空间分布对分层的响应
同种聚集在南盘江流域的喀斯特老龄林中十分普遍。阳性树种(如,白栎、化香树、栓皮栎)和耐阴树种分别偏好生长在遮荫少和多的地方,易形成种内聚集。同时,受裸露岩石和地形的阻挡,种子难以扩散,聚集性加强。林分上层种内聚集程度比林分下层低,暗示林分上层树种周围是其他树种的概率比林分下层的大,即大树混交度高,而小树混交度低。Wang 等人[24]和Pommerening 等人[37]发现大树周围多是异种,而小树周围多是同种,Li等人[25]亦发现松栎混交林的树木大小和混交度均呈显著正相关,皆支持本研究的结果。这种现象可能与负密度制约有关[31],也有人将其解释为竞争效应[38]。邻体死亡为其他树种的到来和生存提供了机会,也使存活的树木分布格局趋向规则分布。林分上层的个体大小差异明显,小径木聚集,而林分下层多为同种小乔木和乔木的小树,个体大小相近。林分上层的小径木可能比大径木生长缓慢,加速个体大小分化[39]。物种生活史阶段和生物特性的差异也可以导致上下层树木大小分化[12]。较大的分化也说明林分上层承受的竞争压力大于林分下层,与它们的分布格局相符,即大小分化随演替的进行而越明显[40]。垂直分层上的树木大小分化以及物种分配可更好地占据和利用生长空间[37]。
4 结论
垂直结构分析有助于认知喀斯特森林层内、层间的物种数量、组分和关联以及群落构建,从而揭示物种多样性形成和共存机理。中国西南地区喀斯特栎类老龄林的垂直分层明显,呈上下两层。林分上层主要由乔木树种组成,径阶分布范围广,物种多度和丰富度远低于林分下层。林分下层的小乔木、幼苗、幼树数量充足,更新状态良好,每个径阶都有足够多的物种和个体,强力支撑群落的物种多样性和稳定。排除生境异质性的影响,林分上层主要呈随机分布,层内竞争较弱。层间的物种组成、个体数量及物种多样性不同,与种间和上下层的分布格局有关。层间以及主要树种之间以随机关联为主,种间竞争缓和,垂直分层有利于物种共存和提高群落稳定性。林分上层和林分下层的同种、小树聚集,表明大小分化在喀斯特森林的物种分配和生境资源利用方面起关键作用,可能是群落演替的重要驱动力。这些发现丰富了有关喀斯特植被和林分垂直结构的研究,强调了垂直分层可能是物种共存和群落维持的重要机制,应在喀斯特森林植被恢复、群落重建和生物多样性保护中予以重视。
猜你喜欢
植物研究(2023年5期)2023-09-09
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
阅读(低年级)(2020年8期)2020-11-06
乡村地理(2019年2期)2019-11-16
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
数学杂志(2017年3期)2017-06-15
少年文艺·开心阅读作文(2017年1期)2017-02-24
考古与文物(2016年5期)2016-12-21
杂草学报(2015年2期)2016-01-04
文化月刊·下旬刊(2014年6期)2014-08-28
