
尾巨桉林分生长对间伐和修枝响应的分析与预测
2024-04-14 12:51张士韬欧阳林男陈少雄杨嘉麒
林业科学研究 2024年2期
张士韬,欧阳林男,陈少雄*,杨嘉麒
(1. 中国林业科学研究院速生树木研究所,广东 湛江 524022;2. 南京林业大学林学院,江苏 南京 210037)
人工林是世界森林资源的重要组成部分。它在木材生产、改善环境和景观建设和减缓气候变化等方面发挥着重要作用。随着世界整体对自然森林的保护越来越强,人工林所承担的经济和生态压力越来越重[1]。我国是全球第二大木材消耗国,人工林的大规模建立支撑了我国现代林业的发展,其覆盖面积居世界第一,达7 954 万hm²,2022 年人工造林120.1 万hm²,占全年总造林面积的31.4%[2]。但由于营林技术不规范,我国人工林仍然面临着结构混乱、质量低下、生态稳定性差和生态功能较弱等问题,不合理采伐和幼龄林超强度采伐现象依然普遍[3]。我国木材进口量仍是全球第一[4],人工林发展的一系列问题导致其无法满足木材刚性需求。人工林抚育是保障幼林成活、成林、成材的关键措施[5]。间伐与修枝是人工林抚育的重要手段,两项手段及其变化,如方式、强度、时机和地点等都直接作用到树冠结构[6],而树冠结构、大小等都通过影响林木光合作用和各器官生物量的分配从而改变林分生长[7]。合理的间伐和修枝通过调整林分和枝条间的密度,能优化林分结构,降低林木竞争压力,促进保留木和枝条的生长发育,缩短林木培育时间[8]。
桉 树 是 桃 金 娘 科(Myrtaceae) 桉属(Eucalyptus)、伞房属(Corymbia) 和杯果木属(Angophora)植物的统称,是国际上主要的速生人工林树种,其覆盖面积超过2 500 万hm²。到2019 年,我国桉树人工林已覆盖546.74 万hm²的面积,在我国人工林总面积中居第三位[9]。它对维护我国国家战略木材安全、促进经济发展和维持生态平衡等方面作出了巨大贡献。尾巨桉(Eucalyptus urophylla×Eucalyptus grandis)是 尾 叶桉(Eucalyptus urophyllaS.T. Blake)和 巨桉(Eucalyptus grandisW. Mill ex Maiden)杂交种,有着干形好、生长发育迅速、抗逆性强的优点[10],在20 世纪80 年代在我国成功杂交获得尾巨桉后,其逐渐作为华南地区最主要的速生桉树杂交种之一[11]。现有不少研究表明,不同强度的间伐和修枝对尾巨桉人工林林分生长的影响有差异[12-13]。但这些研究多是单独研究间伐或修枝的效果,少有考虑间伐和修枝一并进行而产生的综合效应,对尾巨桉人工林培育的指导作用略显不足。鉴于此,本研究以尾巨桉无性系为研究对象,以4 种不同强度的间伐和修枝组成16 种处理,分析不同处理对尾巨桉人工林林分生长的影响,并对不同间伐和修枝及其交互效应进行综合评价,构建并充分验证尾巨桉林分生长增量指标对间伐和修枝响应的BP 人工神经网络预测模型,为尾巨桉人工林的大径材培育提供科学合理的理论依据和实践参考。
1 材料与方法
1.1 试验地概况
试验地位于广东省遂溪县岭北镇南方国家林木种苗示范基地(21°26′ N,110°09′ E),属于北热带湿润大区雷琼区北缘,海洋性季风气候,年平均气温23.1℃,全年降雨量约为1 567 mm,雨季是5 月—9 月,年相对湿度为80.4%,海拔101.56~136.47 m,土壤类型主要是浅海沉积物砖红壤、玄武岩砖红壤、砂页岩红壤,pH 值4.5~5.3,有机质含量大于10 g·kg-1。试验林地共计3.34 hm2,处于低山丘陵地带,样地坡度在5°~10°。2016 年4 月造林,造林密度1 250 株·hm-2,挖机挖穴(70 cm × 70 cm × 60 cm),基肥1 次(500 g·穴-1),第2 年开始追肥2 次,每年追肥1 次(500 g·株-1)。在试验林地中布设48 块样地,每块样地面积400 m2,相邻样地间设置保护行。幼林最初3 a 每年除草2 次,3 a 后每年除草1 次。
1.2 试验设计与指标测定
2018 年8 月,选样地内28 月龄的尾巨桉为研究对象,采用双因素试验设计,以间伐株数占样地初始株数的百分比确立4 种间伐强度,分别是60%(强度间伐)、40%(中度间伐)、20%(弱度间伐)和0%(不间伐对照),对应样地保留林分密度为400、600、800 和1 250 株·hm-2。以修枝后无枝条树干高占平均树高的百分比为标准确立4 种修枝强度,分别是38.18%(5 m)、42.39%(6 m)、48.18%(7 m)和0%(不修枝对照),两者共组成16 种处理,每种处理3 块样地依据设计进行间伐和修枝处理。在处理后立即对标准地进行林分生长调查,对样地内每株尾巨桉进行每木检尺,测量胸径(DBH)、树高(H)、枝下高(He)和两个方向的冠幅,计算平均冠幅(C)、用广西林科院研制的材积公式计算单株材积(V)[14]:
依据单株材积计算每块样地的蓄积量(SV),作为样地林分基础生长数据,依据胸径统计情况划分样地尾巨桉径阶分布。由于林分基础生长数据中基础胸径大于8 cm 的尾巨桉占据99% 以上,满足小径材的出材标准[15],故将当次测量的蓄积量与间伐材的材积量之和作为当次测量样地出材量。
在之后的第12、24、44 和56 个月(对应林龄分别为3.3、4.3、6.0、7.0 a)进行相同调查组成4 个时期的生长指标,与林分基础生长对应指标的差值作为林分生长累积增量指标,包括树高累积增量(Hi)、平均冠幅累积增量(Ci)、胸径累积增量(DBHi)、单株材积累积增量(Vi)。因此该双因素试验的方差分析数学模型为:
其中yTPk是4 种增量因子在不同效应影响下的观测值,μi是样本均值,αT和βP分别是间伐和修枝的主效应值,γTP是间伐和修枝各自水平的交互效应,eTPk是随机误差。
依据模型(1),分别对前3 个时期不同增量因子受间伐和修枝的主体效应和交互效应进行显著性检验和多重比较。由于第56 个月的指标数据量远小于前3 个时期,故第4 个时期指标仅用于BPNN 模型预测性能的验证。
1.3 数据分析
1.3.1 基本数据分析 数据采用WPS Office 2019 进行整理,Matlab R2019b 用于BP 神经网络(BPNN)的构建和内部测试,R-4.2.2 用于双因素方差分析(two-way ANOVA)、Duncan 多重比较和BPNN 的验证,Origin 2021 用于制作矢量图。
1.3.2 BPNN 的构建 首先是选择模型输入与输出因子,输入因子包括:间伐强度(T)、修枝强度(P)、林龄(A)。输出因子为林分生长增量因子,包括胸径、树高、冠幅和单株材积增量。选择的各个林分生长因子的意义不同,并且在数值上相差较大,为了消除数值量级差异对模型建立和预测的影响,需用公式(2)对数据进行归一化处理,转化为无量纲数据,该步骤将在Matlab 中自动完成。再将样本分为训练集和测试集,训练集占数据的80%,用于模型构建训练,测试集占20%,用于模型训练性能评估。
式中:Xs为归一化后的值;X为归一化前的值;Xmin为变量值下限;Xmax为变量值上限。
对训练好的BP 神经网络模型输入测试集的输入参数矩阵,将网络输出的林分生长增量训练值矩阵与测试集的实际值矩阵进行比较检验,结合检验结果,判断所建BPNN 预测林分生长增量的可行性。矩阵检验的方法包括均方根误差(RMSE),Kappa 一致性检验和Pearson 相关性分析。
RMSE是将BP 神经网络的训练值K和实际值O的均方差开根号,RMSE的定义见公式(3),体现的BP 神经网络训练的准确程度,值越小,训练值越接近于实际值,模型越有效,反之越差。Kappa检验是一致性检验的指标,也可以检验模型训练结果和实际结果是否一致。Kappa系数的大小决定了一致性的程度(见公式(4)),当Kappa系数在0~0.2 时,一致性程度较差,0.2~0.4 代表一致性程度一般,0.4~0.6 代表程度中等,0.6~0.8 代表程度较强,0.8~1 代表有很强的一致性。Pearson相关性分析的r值反映两个变量间的线性相关程度(见公式(5)),r越大相关性越高,反之越差。
式中:Ti是用于检验结果的每个实际数据,N是用于检验结果的数据个数;n是所有实际数据和训练数据之和;a和b分别代表的是训练数据和实际数据;mK、mO是实际值K和训练值O的平均值
本研究的数据关系为非线性,因此BP 神经网络采用单隐含层结构。隐含层神经元节点数的确定目前没有标准,本研究以比较法确定,鉴于确立原则之一是其数量小于输入因子数的2 倍[16],分别建立隐含层神经元节点数为1、2、3、4、5 的BPNN,在建立过程中对相同神经元节点数的BPNN 进行10 次重复训练,对训练完成的BPNN 输入测试集进行性能评估,具体方法为:(1)完成训练并求得不同模型不同因变量的RMSE、Kappa系数和相关系数r值;(2) 取10 次训练性能平均值作为评测值;(3) 以RMSE最低、Kappa值和r值最高的节点数为标准,确定训练效果最好的BPNN 模型。
2 结果与分析
2.1 间伐和修枝及交互效应对尾巨桉人工林林分生长的影响
2.1.1 间伐对尾巨桉林分生长的影响 图1 展示了3 种林龄下林分生长增量随间伐强度增加的变化趋势。样地胸径增量随着间伐强度的增大,呈先减少后增加的趋势,且此趋势随着林龄的增加变得越明显(图1A)。样地单株材积增量和树高增量的变化趋势与胸径增量变化趋势类似,不同的是不间伐的样地树高增量显著最高(图1B、D)。60%间伐在3.3、4.3 和6 a 这3 个林龄阶段均有着显著最高的胸径增量(1.72、3.78 和6.59 cm)和最高的单株材积增量(0.56、1.36 和2.8 m3)。间伐均显著提高样地的冠幅增量,其中20%和60%间伐的提升效果比40%间伐更明显(图1C)。尾巨桉林分的出材量随着间伐强度的增大而减少,且出材量的减少程度随着林龄增加而显著增大。综上,间伐促进冠幅增长;随着间伐强度增大,胸径增量、树高增量和单株材积增量均有着先减小后增大的趋势,且这种趋势会随着林龄增加而变得更明显,说明40%及以上强度的间伐更有助于胸径和单株材积的增长,但至少在6.0 a 内会显著降低出材量。
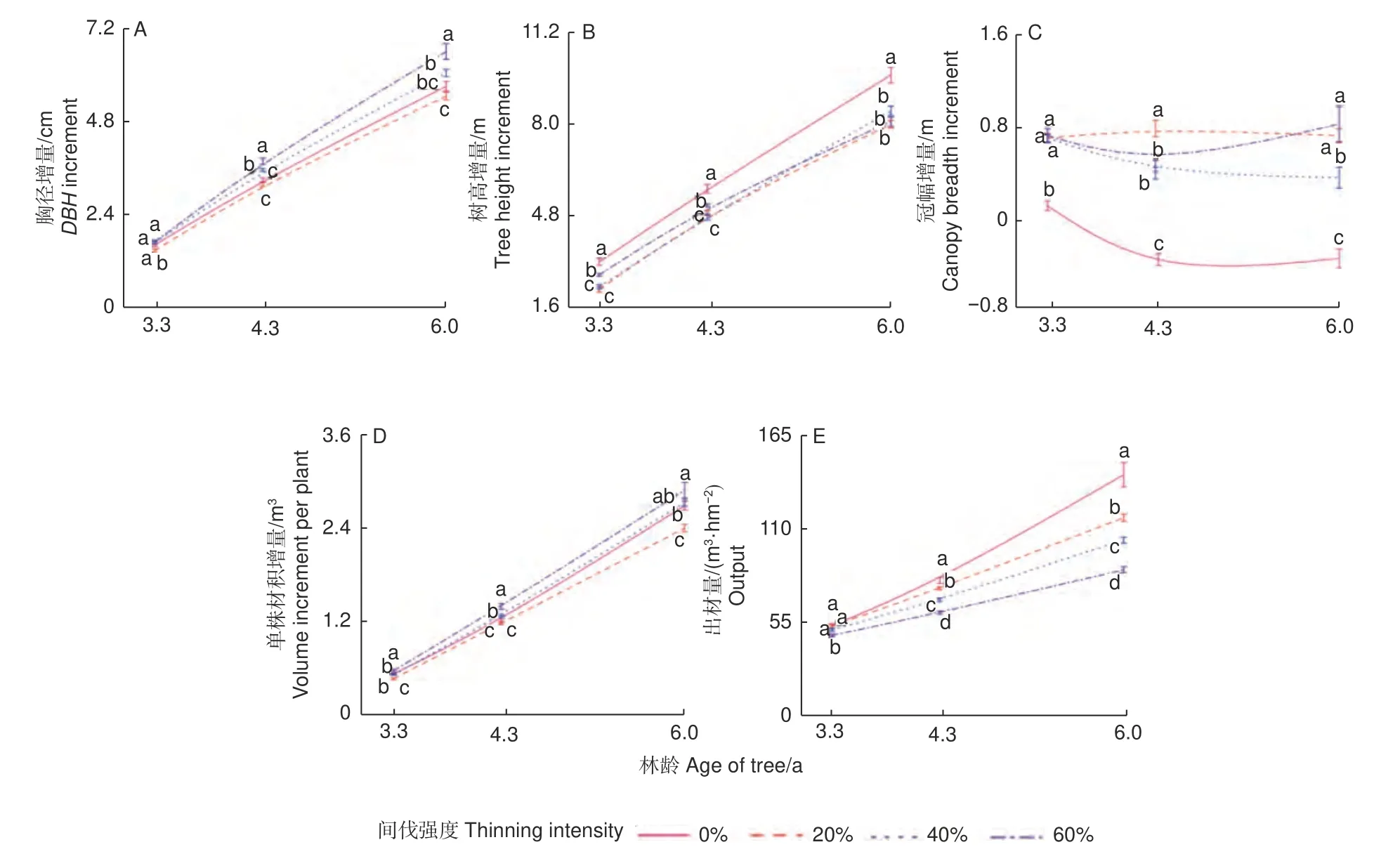
图1 3 种林龄时期不同间伐强度下林分生长增量结果Fig. 1 Results of stand growth increment under different thinning intensities at three stand ages
2.1.2 修枝对尾巨桉林分生长的影响 图2 展示了修枝对林分生长增量的影响趋势。胸径、树高和单株材积增量随修枝强度的增大,均呈现先增大再减小,之后再增大的趋势(图2A、B、D)。38.18%修枝在林龄为3.3、4.3 和6.0 a 阶段均有着最高的胸径增量(1.76、3.67 和6.25 cm)、树高增量(3.14 、5.39 和8.93 m) 和单株材积增量(0.58、1.30 和2.72 m3)。修枝显著降低了林分冠幅增量(图2C)。综合5 张图来看,图2B、C 的曲线比图2A、D、E 波动更大,说明修枝对尾巨桉林分生长的影响主要体现在树高和冠幅上。38.18%修枝能促进尾巨桉胸径和树高生长,是最适宜尾巨桉6.0 a 内林分生长的修枝强度,与不修枝相比,修枝对尾巨桉出材量没有影响,但显著降低了冠幅增量。
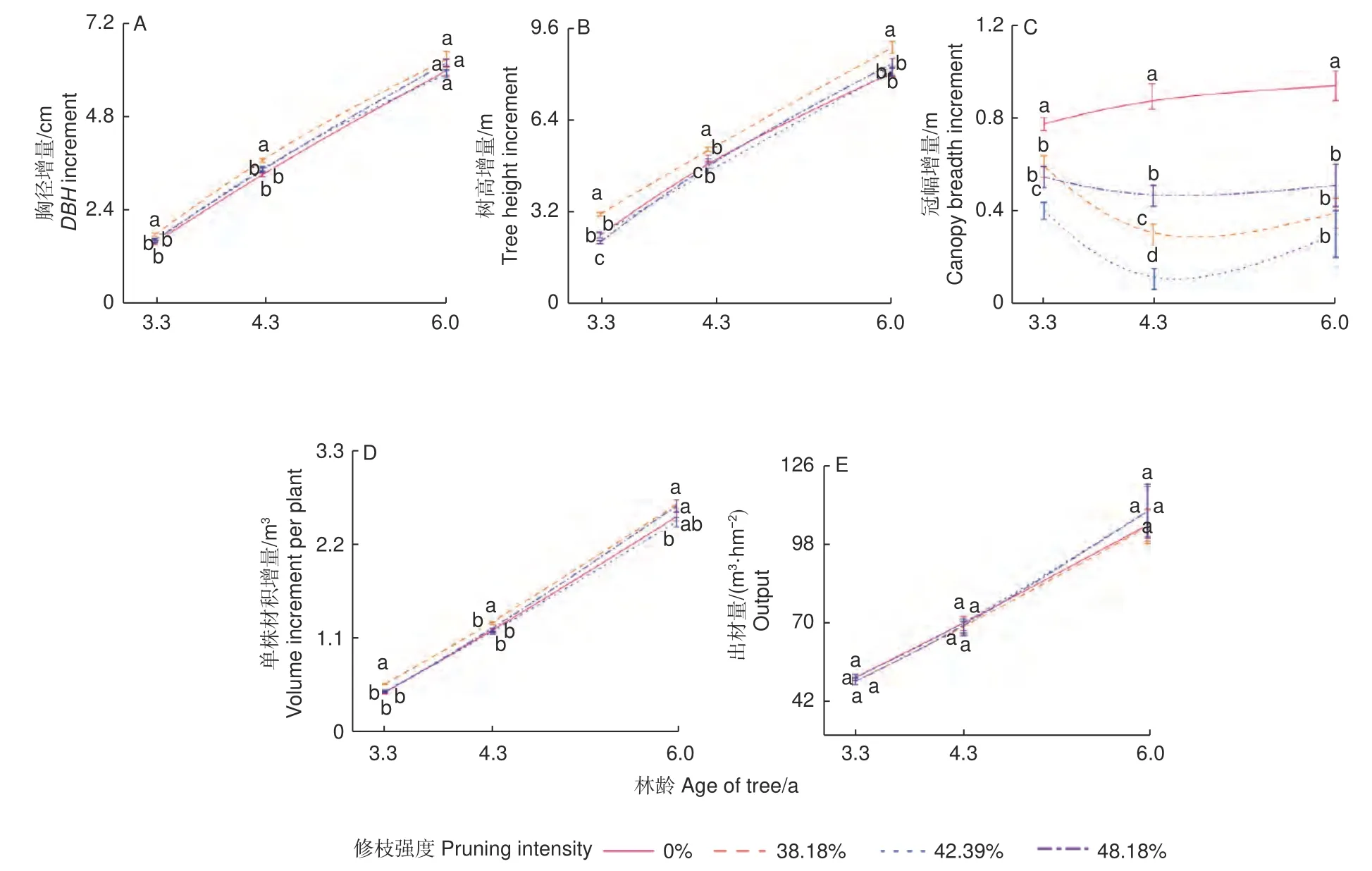
图2 3 种林龄时期不同修枝强度下林分生长增量结果Fig. 2 Results of stand growth increment under different pruning intensities at three stand ages
2.1.3 交互效应对尾巨桉林分生长的影响 通过双因素方差分析中交互效应的显著性检验得出,间伐和修枝的交互效应对胸径有显著影响,对树高、冠幅和单株材积的影响极显著(表1)。交互效应的结果,以颜色色系冷暖程度来表示不同间伐和修枝处理后的尾巨桉人工林胸径、树高、冠幅和单株材积增量平均值,不同处理颜色具有差异则代表相应的处理差异明显(图3)。结果表明,60%间伐 + 48.18%修枝处理最能促进胸径(3.00 cm)和单株材积(1.16 m3)的增长,且显著优于其他所有处理(图3A、D)。就树高生长而言,不间伐情况下修枝的促进效果显著优于不修枝处理,其中不间伐 + 38.18%修枝最优(图3B)。间伐比修枝更有助于冠幅生长(图3C),其中增量最大的是20%间伐 + 不修枝。综合来看,60% 间伐 +48.18%修枝处理最能促进尾巨桉胸径和单株材积生长,对冠幅生长也有显著促进作用,是促进尾巨桉林分生长的最佳抚育方式。

表1 基于间伐与修枝交互效应的方差分析Table 1 Variance analysis table based on the interaction effect of thinning and pruning
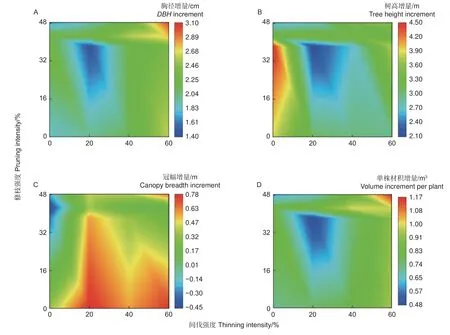
图3 交互效应对林分生长增量影响的热力图Fig. 3 Thermal map of the interaction effect on stand growth increment
2.2 间伐和修枝及交互效应对尾巨桉人工林径阶分布的影响
2.2.1 间伐对尾巨桉径阶分布的影响 图4A 展示了不同时期4 种间伐强度下尾巨桉各径阶株数占总株数的百分比。间伐前,各林分的尾巨桉株数高峰均位于径阶12 cm,不同的是与准备间伐的样地相比,对照林分有更多尾巨桉位于径阶10 cm(占32.37%)。3.3 a 林龄时,尾巨桉径阶高峰均是14 cm,该径阶株数占比分别是44.53%(不间伐)、58.74%(20% 间伐)、66.55%(40%间伐)和76.92%(60%间伐),所有林分的第二高峰均是12 cm,说明尾巨桉本身胸径早期生长发育迅速,间伐在短期内不足以对径阶分布产生影响。4.3 a 林龄时,不同间伐强度的径阶分布高峰出现差异,不间伐和20% 强度间伐径阶高峰均为14 cm,分别占37.41% 和51.89%,第二高峰是16 cm,分别占35.25% 和43.08%;40% 和60%强度间伐分布结果则恰好相反,径阶高峰位于16 cm 的分别占57.54%和70.36%,径阶位于14 cm的分别占36.49% 和17.79%,依据图4A 的初始径阶分布和表1 的胸径增量多重比较,40%和60%间伐的径阶分布随林龄增加逐渐向右偏移,中径材占比有增多的趋势,并且60%高强度间伐下这种趋势更加明显。6 a 林龄时,不间伐林分的径阶高峰为16 cm(占42.11%),间伐林分的径阶高峰都在18 cm,不同的是40%和60%间伐的第二高峰是20 cm,分别占15.08% 和28.28%,20%间伐的第二高峰则是16 cm(占40.52%)。由此表明,间伐从长期来看均利于林分径阶分布右偏,高强度的间伐更有助于优化尾巨桉林分结构,促进林木胸径生长,60% 强度间伐的效果最为明显。

图4 不同时期4 种间伐和4 种修枝强度下林分径阶分布Fig. 4 Distribution of stand diameters and steps under 4 thinning and 4 pruning intensities at different periods
2.2.2 修枝对尾巨桉径阶分布的影响 图4B 展示了修枝对尾巨桉径阶分布的影响。林分初始径阶分布高峰均位于12 cm,不修枝林分占比最高(85.77%)。4 个时期不同修枝下尾巨桉径阶分布的高峰、第二高峰情况基本一致,说明修枝对尾巨桉径阶分布的影响程度远不如间伐强。差异出现在林龄6 a 时,4 种修枝强度林地径阶分布高峰均是18 cm,但不修枝的林地在该径阶占比最高(65.63%),比修枝林地高了约10%,而修枝林地尾巨桉位于20 cm 和22 cm 的径阶占比之和恰好比不修枝林地多出约10%,表明修枝从长期来看促进尾巨桉径阶分布右偏。综上,修枝利于尾巨桉胸径生长,效果不如间伐,不同强度的修枝对尾巨桉林分径阶分布的影响没有显著差异。
2.2.3 交互效应对尾巨桉径阶分布的影响 通过计算不同处理在同一径阶下分布比例的方差,以该径阶占比在不同处理下的离散程度来衡量受交互效应的影响程度[17],方差越大交互效应的影响程度越高。本研究中,8 cm 和22 cm 径阶在4 个林龄下的方差很小,受交互效应影响程度很低,其他径阶受交互效应的影响均有不同程度的变异,因此有必要分析10 cm 到20 cm 共6 个径阶受交互效应的影响(表2)。间伐前2.3 a 林龄时,林木大多数径阶均位于12 cm(图5B)。3.3 a 林龄时,林木径阶在10、12 和14 cm 均有分布,间伐强度越高,径阶位于14 cm 的林木越多(图5A~C),60%间伐 + 48.18%修枝的样地林木位于14 cm的径阶占比最高(89.58%)。4.3 a 林龄时,40%及以上的间伐样地林木径阶更多位于16 cm,40%以下的间伐样地则多数位于径阶14 cm,不间伐的样地林木仍有部分径阶位于10 cm 和12 cm,而60%间伐 + 48.18%修枝的样地有47.06%的林木径阶位于20 cm(图5)。6 a 林龄时,少数不间伐和20% 间伐的样地径阶位于16 cm(图5D),40% 间伐和60% 间伐的样地林木多数径阶位于18 cm(图5E),在径阶20 cm 的分布中,60% 间伐 + 42.39%修枝处理的占比最高(50.00%,图5F)。综合图5 来看,在林龄高于2.3 a 时,间伐强度越大,样地林木位于更高径阶的占比越高,即与修枝相比,间伐更有助于尾巨桉林分径阶分布右偏。42.39%及以上的修枝强度与60% 强度间伐相结合的样地林木胸径生长迅速,是优秀的人工林抚育措施。

表2 8 个径阶在不同处理下占比的方差 Table 2 The variance of the proportions of the same diameter grade under different treatments
2.3 BPNN 的构建与性能分析
2.3.1 BPNN 的构建 基于Matlab R2019b 平台构建输入因子与林分生长增量因子的BP 神经网络模型,模型参数设置如下:激活函数采用“tansig”(输入层)和“purelin”(输出层);训练函数为“traninbr”;网络学习速率net.trainParam.lr=0.05;允许训练最大步数为1 500;期望误差为0.000 1。训练集内部设置训练数据占80%,验证数据和测试数据占10%,对5 种模型的训练性能评估结果见表3。
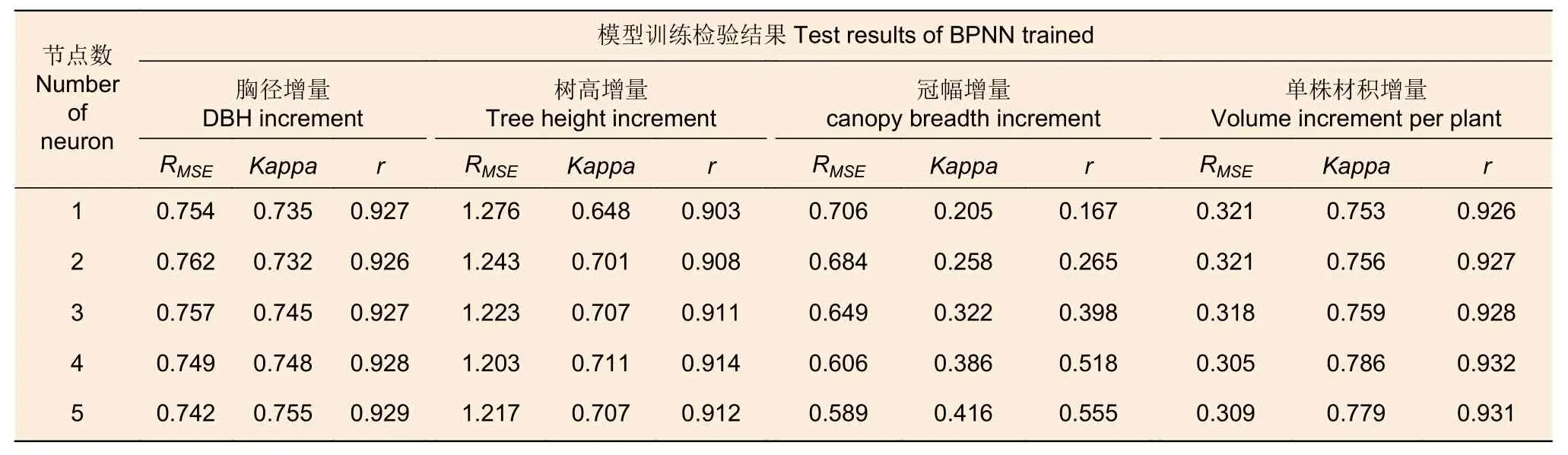
表3 不同隐含层节点数的BPNN 的训练性能结果 Table 3 Trained performance results of BPNN with different number of hidden neurons
2.3.2 BPNN 的选择 如表3 所示,除了胸径增量训练效果最优的是隐含层节点数为5 的BPNN,次优的节点数为4 以外,其余增量因子最优的均是节点数为4 的BPNN。BPNN 训练冠幅增量的Kappa 系数和相关系数小于其它因子,其中冠幅一致性等级明显低于其他因子。综上,BPNN 对冠幅增量的训练效果不如其它林分生长增量因子。隐含层节点数为4 的BPNN 训练效果最佳,设最佳预测模型的隐含层神经元分别为M1、M2、M3和M4,训练在第430 次时终止,此时根据权值矩阵和阈值矩阵结果确定传递函数。
输入层到隐含层的传递函数表达式为:
隐含层到输出层的传递函数表达式为:
最佳BPNN 结构见图6:Lm1为输入层到隐含层的偏移量,Lm2为隐含层到输出层的偏移量。
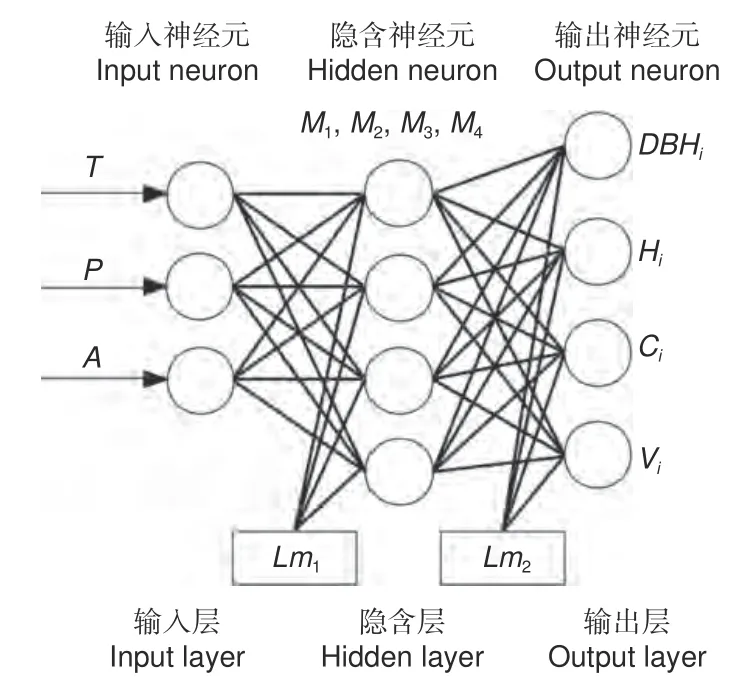
图6 BP 神经网络模型结构Fig. 6 Structure diagram of BP neural network model
将训练集中的20% 用于模型仿真测试数据,上述模型的网络输出仿真值与训练集的实际值进行仿真比较,决定系数R2反映BP 神经网络的仿真值对实际值的决定程度,其值越大说明内部仿真结果越准确。比较结果如图7 所示。
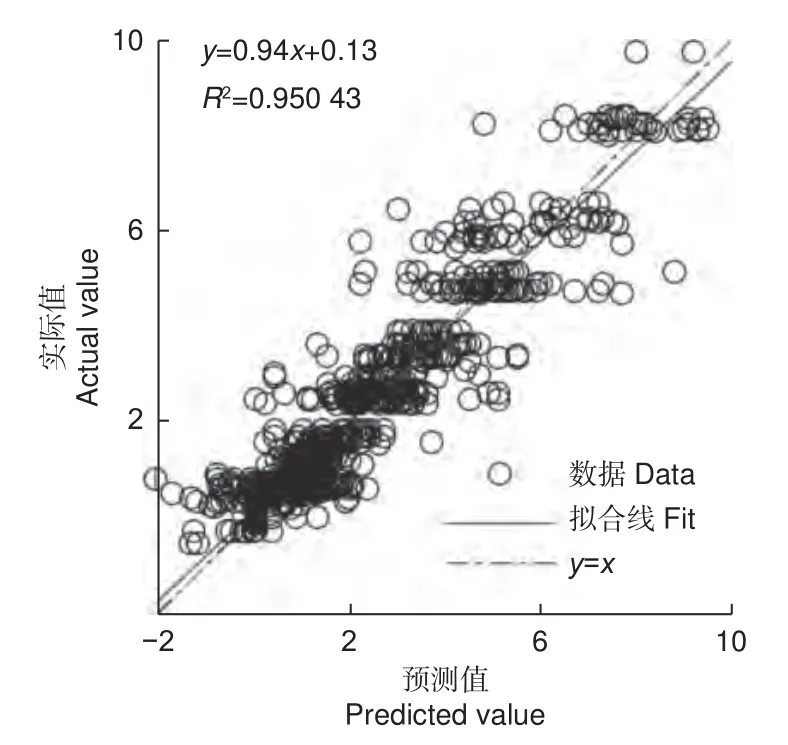
图7 基于贝叶斯正则化算法的林分生长增量因子预测理论值与实际值回归分析Fig. 7 Regression line of predicted and actual values of stand growth increment factor based on Bayesian regularization algorithm
从图7 可以看出,观测值和训练值贝叶斯正则化算法线性方程拟合R2为0.950 4,说明该模型在拟合程度较高,可以解释95.04%的训练值增量因子变异。将7 a 林龄的数据用于模型预测性能验证数据,以模型的网络输出预测值与不同间伐和修枝处理的数据平均值进行比较,以模型训练评估参数用于模型预测性能检验,结果如表4 所示。
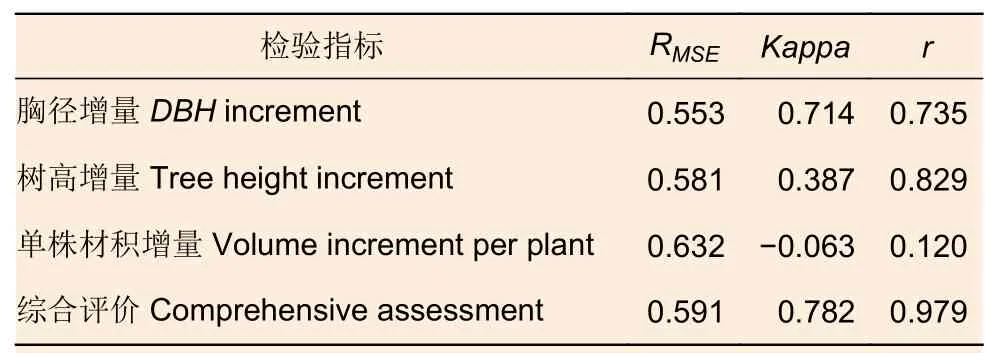
表4 模型预测检验结果Table 4 Test results of BPNN predicted
由表4 又得知,BPNN 模型对胸径和综合评价的预测结果很好,而对树高和单株材积的预测结果较差,可能因为第84 个月仅调查少部分尾巨桉的树高,而调查了林地绝大多数尾巨桉的胸径,导致树高数据量远小于胸径数据量,低于大样本统计的要求,由此可得,该模型至少可以对超过训练样本林龄12 个月内的尾巨桉胸径增长进行有效预测。综上,具有4 个隐含层神经元节点数的BPNN 模型是较为理想的尾巨桉林分生长增量对间伐和修枝的响应模型,可有效预测7 年内的尾巨桉林分生长。
3 讨论
抚育间伐和修枝是改善人工林林分生长与结构的重要措施[18]。持续4 a 的抚育结果表明,不同间伐和修枝处理对尾巨桉林分生长的影响不同。综合来看,更高强度的间伐更有助于尾巨桉胸径、树高和单株材积的生长,这与其他大多数树种的间伐结果基本一致,主要因为间伐减少了林木对土壤养分和水分的争夺,将资源集中在更少的林木和枝叶上,从而促进了林木生长,促进效果因树种、地域、间伐方式和间隔时间等而存在差异[16,19-20]。在本研究中,不同强度修枝对尾巨桉林分生长同样有不同程度影响,这与Pelkki 等人[21-22]的研究结论不一致,可能因为尾巨桉属于典型的速生树种,具有更发达的根系,在结合间伐抚育下,它对环境(气候、土壤条件、光照)的敏感性更高,在相同观察时长下,更优越的环境条件对桉树生长的促进作用更明显[23]。6 a 林龄时,38.18% 修枝后林分胸径、树高、冠幅和单株材积增量均高于42.39%修枝林分,说明对尾巨桉林分生长的促进效果并非随着修枝强度增大而增大,38.18%修枝是较适宜的修枝强度。以上考虑的都是单独使用一种抚育措施时的情况,交互效应结果表明,若两种措施同时进行抚育,最好的方式仍是较高强度间伐与较高强度修枝相结合,其促进林分生长的效果最佳。但无论哪种强度的间伐都会显著降低林分样地的出材量,因为从短期来看间伐样地蓄积量与间伐材积量之和远小于不间伐样地蓄积量,而有研究表明,随着间伐林单株材积增长的更加迅速,它们之间的差值会随林龄增加逐渐减小,并在某一年变为负数[24],因而间伐更有助于长期时间大径材的培育。
间伐能对尾巨桉径阶分布产生明显的影响,间伐后随着林龄的增加分布逐渐右偏,且间伐强度越大,右偏的程度越大,早期出现更大胸径的林木数量越多,说明随着时间推移,高强度间伐下可能出现尾巨桉大径材的概率越高,数量越多,修枝也能促使径阶分布右偏,但效果不如间伐,上述结论与Fontan 等[25]和Medeiros 等[26]的结果不同,因为间伐和修枝后的生长响应还受到树冠结构、光合作用、地上地下养分循环等的综合影响。因此,要培育大径材应该较高强度的间伐与适当修枝相结合进行抚育。
BP 神经网络模型是一种较为新颖的多元非线性模型,比传统模型的拟合优度更高,近年来多用于林分生长预测中[27-28],但多是基于自然因子对林木某一特征进行预测。鉴于间伐和修枝对林分生长的影响是非线性的,本研究建立林分生长因子对人为抚育措施响应的BP 神经网络模型,该模型具有精度高、避免选型等优点,但由于硬件条件不足,研究的输入因子未包含立地因子和气候因子,这些因子也会对林分生长产生影响,因此该模型有待进一步完善,同时BP 神经网络的整体训练过程属于科技黑箱,尚无法从模型得知间伐和修枝如何具体影响林分生长,其机理性研究有待探讨。
4 结论
尾巨桉林分生长受间伐、修枝强度和林龄的影响。间伐有助于林分生长和径阶分布右偏,且效果随着间伐强度的增大而增大,60%的间伐是本研究中最优间伐强度。修枝对林分生长和径阶分布的有一定的促进效果,但不如间伐,通常与间伐相结合来抚育,38.18% 修枝是相对更优的修枝强度。本研究建立了林分生长因子对间伐和修枝响应的BP 神经网络模型,并经过性能测试证明其可用于7 年内尾巨桉林分生长的整体反应预测,依据模型能更科学地选择促进尾巨桉林分生长的最适间伐强度和修枝强度。本研究方法及结果为之后尾巨桉营林提供参考,同时还可以为其他树种的间伐和修枝提供理论和实践依据。
猜你喜欢
甘肃林业科技(2021年3期)2021-03-30
防护林科技(2020年10期)2020-12-22
安徽农学通报(2020年17期)2020-10-09
东坡赤壁诗词(2019年3期)2019-07-05
山西林业科技(2018年4期)2018-03-19
防护林科技(2016年9期)2016-09-27
防护林科技(2016年5期)2016-09-05
贵州科学(2016年2期)2016-06-13
华南农业大学学报(2016年3期)2016-06-01
中南林业科技大学学报(2015年9期)2015-12-21
