
粤东6种阔叶树木材密度及其影响因子研究
2016-06-01 09:27:59徐明锋柯娴氡王永强苏志尧
华南农业大学学报 2016年3期
徐明锋, 柯娴氡, 张 毅, 王永强, 苏志尧
(华南农业大学 林学与风景园林学院,广东 广州 510642)
粤东6种阔叶树木材密度及其影响因子研究
徐明锋, 柯娴氡, 张毅, 王永强, 苏志尧
(华南农业大学 林学与风景园林学院,广东 广州 510642)
摘要:【目的】分析不同影响因子对木材密度的影响,为木材密度的良种选育和碳汇计量提供理论和数据支撑。【方法】在康禾自然保护区样地中,通过群落分析筛选出6个优势树种,测定其木材密度(包括生材密度和基本木材密度),通过多种统计软件分析不同树种、不同径阶和不同坡向对木材密度的影响,以及树木性状因子与木材密度的相关关系。【结果】木材密度在树种间的差异性极显著,生材密度、基本木材密度和含水率平均值最大的分别是米槠 Castanopsis carlesii 、红锥 C. hystrix 和米槠,其数值分别为1.10 g·cm-3、0.53 g·cm-3和122.84%。木材密度的径阶差异性分析结果显示,除了栲C. fargesii 以外,其他树种的木材密度在径阶间变化并不显著。木材密度在坡向上的差异性分析结果表明,6个树种的木材密度在坡向间的差异均不显著。非参数相关分析结果表明,胸径、树高与生材密度的相关性强,而与基本木材密度的相关性弱。【结论】红锥含水率低,基本木材密度高,具备优质木材的条件。木材密度随着径阶的增加有上升的趋势,但在坡向上没有规律性变化。生材密度、基本木材密度与胸径、树高的关系并不一致,可能是因为生材密度受到含水率增加的影响。
关键词:木材密度; 生材密度; 基本木材密度; 径阶; 坡向;阔叶树;影响因子
木材是一种应用广泛的原材料[1-2],木材密度是木材材性的重要指标[3],在木材生产加工过程中,对木材的强度、性能和木材产品质量有着直接的影响[4]。在森林生态系统进程中,木材密度也发挥了重要作用,既是树种的一个重要的功能性状,又是许多功能过程的核心,比如碳分配、碳增量、水运输、生物量等[5-6]。另外,木材密度是森林碳汇计量中一个重要的生物量估算因子[7],但是木材密度在一些天然林树种中仍缺少深入研究。
木材密度有着极其复杂的物理性质和变化规律[7]。树种差异对木材密度影响明显,不同树种间木材密度差异可达数倍[8],不同的树种和生活型对木材密度造成很大差异,比如,针叶树的木材通常较为疏松柔软,而阔叶树材则多为致密坚硬[1]。一些树木生长特性如生长速度、树龄、生长部位[9]、树高等对木材密度也有一定的影响。另外,外界环境因子可以通过改变树木生长环境中的水、肥、气、热,从而影响植物的木材密度变化[10-12]。但是,对于环境因子与木材密度的具体关系目前还未明确,比如,Patino等[13]的研究认为,在年降水量小于 3 000 mm 的亚马逊森林中,木材密度与降水之间表现正相关关系;而Wiemann等[14]对美洲地区森林的研究则认为木材密度与降水的关系并不显著。
目前,国内研究主要针对木材密度在不同树种性状上的变化情况,以及一些营林措施和遗传选育与木材密度的关系,而关于其他环境因子对木材密度影响的研究仍不够深入[10, 15-16]。本文在10 hm2固定样地每木检尺的基础上,筛选出6种粤东地区较为常见的阔叶树种,进行木材密度(包括生材和基本木材密度)测定,研究木材密度在种间、径阶、坡向的变化情况,为林木选育、森林生态效益评价提供数据支撑。
1研究区概况
研究区位于广东省东源县康禾省级自然保护区(23°25′49″~23°31′39″N,116°30′3″ ~116°37′44″E),该保护区处于低山丘陵地带,山体呈东北-西南走向,区内最低点海拔100 m,最高峰白石岗海拔893.3 m。自然保护区全部属东江一级支流“康禾河”流域范围。康禾保护区位于中亚热带和南亚热带交界处,属亚热带季风气候。保护区最高气温 39.3 ℃(1953 年),最低气温-3.8 ℃(1955 年),平均气温 21.1 ℃,年无霜期 343.3 d,年平均日照 2 003.6 h,最大年降雨量 2 756 mm (1997 年),最小年降雨量 1 051 mm (1991 年),年均降水量 1 912 mm,年均蒸发量 1 275.7 mm,年平均相对湿度77.0%。保护区岩石易于风化侵蚀,沟谷陡崖深幽,多呈“U”形。平原呈狭长带状分布于河流两岸及山间,地面平坦。区内成土母岩为花岗岩和砂岩,土壤以赤红壤为主,其次有山地红壤、山地黄壤。区内植物资源丰富,植物区系组成以泛热带和热带亚洲成分最为丰富,分别占30.5%和13.8%,生活型谱以高位芽植物占绝对优势,植被类型主要为亚热带常绿阔叶林,但因人为干扰活动,现有植被主要为次生性较强的常绿阔叶林和针阔叶混交林[17-19]。保护区中10 hm2固定样地林分类型为择伐胸径13 cm大树后自然更新的次生常绿阔叶林,择伐发生在1993年,调查时间在2012年,自然更新时间为19年。样地林分择伐后封山育林,排除强烈的人为干扰活动,林分中米槠Castanopsiscarlesii是建群种,另有小片的杉木Cunninghamialanceolata存于其中。
2研究方法
2.1外业调查及木材密度测定
在康禾保护区中,设置10 hm2固定样地,样地设置在山坡两边,山脊由东向西海拔减低,海拔最低处200 m,最高处375 m。10 hm2样地面积为500 m × 200 m,共分成250个20 m × 20 m样方,以样方为基本单位进行每木检尺调查,每木检尺从1 cm起测,调查的指标有种名、胸径、树高等,胸径采用胸径尺和游标卡尺进行测定,坡向是根据样地设置时使用的全站仪进行测定。用流体静力称衡法进行木材密度测定[20]。
2.2数据分析
2.2.1优势种筛选在PC-ORD 6.0软件中作出群落的优势种曲线,包括物种序位-多度曲线、物种序位-频度曲线;在STATISTICA 8.0软件作出物种序位-重要值曲线、物种序位-断面积曲线。通过序位曲线筛选出符合条件的优势树种,最后通过STATISTICA 8.0对筛选出的6个树种的胸径、树干、蓄积量进行分析。
2.2.2树种、径阶和坡向单因素方差分析运用STATISTICA 8.0软件对树种间、径阶间、坡向间的木材密度进行统计分析。对6个树种的基本木材密度、生材密度和含水率进行单因素方差分析和LSD检验;在区分树种的基础上再对径阶间和坡向间的基本木材密度、生材密度和含水率进行单因素方差分析和LSD检验。
2.2.3树木性状因子与木材密度的非参数Spearman相关分析在SASTISTICA 8.0中对胸径和树高等树种性状因子与基本木材密度、生材密度和含水率进行非参数Spearman相关分析,研究树种性状因子与木材密度之间的相关关系。
3结果与分析
3.1树种筛选
选取重要值大于0.01的物种进行优势种序位排序,从物种序位-重要值曲线(图1 A)可以看出,米槠的重要值远大于其他树种,为群落的建群种,次优势种为木荷Schimasuperba,其他优势树种主要有:罗伞树Ardisiaquinquegona、栲Castanopsisfargesii、鼠刺Iteachinensis、黄杞Engelhardtiaroxburghiana、豺皮樟Litsearotundifoliavar.oblongifolia、黄樟Cinnamomumporrectum、红锥Castanopsishystrix、多毛茜草树Aidiapycnantha、华润楠Machiluschinensis等。从物种序位-多度曲线(图1 B)和物种序位-断面积曲线(图1 D)可以看出,罗伞树、鼠刺、豺皮樟和多毛茜草树的多度序位高,但断面积序位低,原因是它们主要为灌木和小乔木,筛选中将它们排除。黄杞的重要值虽然也较高,但是在亚热带森林中不如黄樟和华润楠常见,另外其属于胡桃科,胡桃科在亚热带森林中分布也不如樟科和壳斗科那么广泛。综上所述,在样地中筛选出米槠、栲、红锥(壳斗科Fagaceae),木荷(山茶科Theaceae),黄樟、华润楠(樟科 Lauraceae)这6个树种。从物种序位-多度曲线(图1 B)可以看出,米槠多度最大,并且与排第2位的木荷有较大差距,米槠是群落的建群种;通过物种序位-频度曲线(图1 C)可以看出,频度最大是木荷,然后是米槠,说明木荷比米槠分布范围更广、更均匀,所选6个树种除了红锥以外,频度均超过150,即其他5个树种在一半以上的样方中都有出现;从物种序位-断面积曲线(图1 D)可以清楚地看出,米槠的断面积远大于第2位的木荷,同时,6个树种的断面积都居于前10位,米槠占总体的51.6%,6个树种断面积的总和超过总体的70%,可以看出6个树种的生物量、碳储量总和在群落中有着足够的优势。总的来说,通过物种序位曲线可以看出,米槠在米槠、木荷、栲、黄樟、红锥和华润楠:在A图中的序位依次为1、2、4、8、9和11;在B图中的序位依次为1、2、9、20、24和19;在C图中的序位依次为2、1、5、8、21和6;在D图中的序位依次为1、2、3、8、5和10。
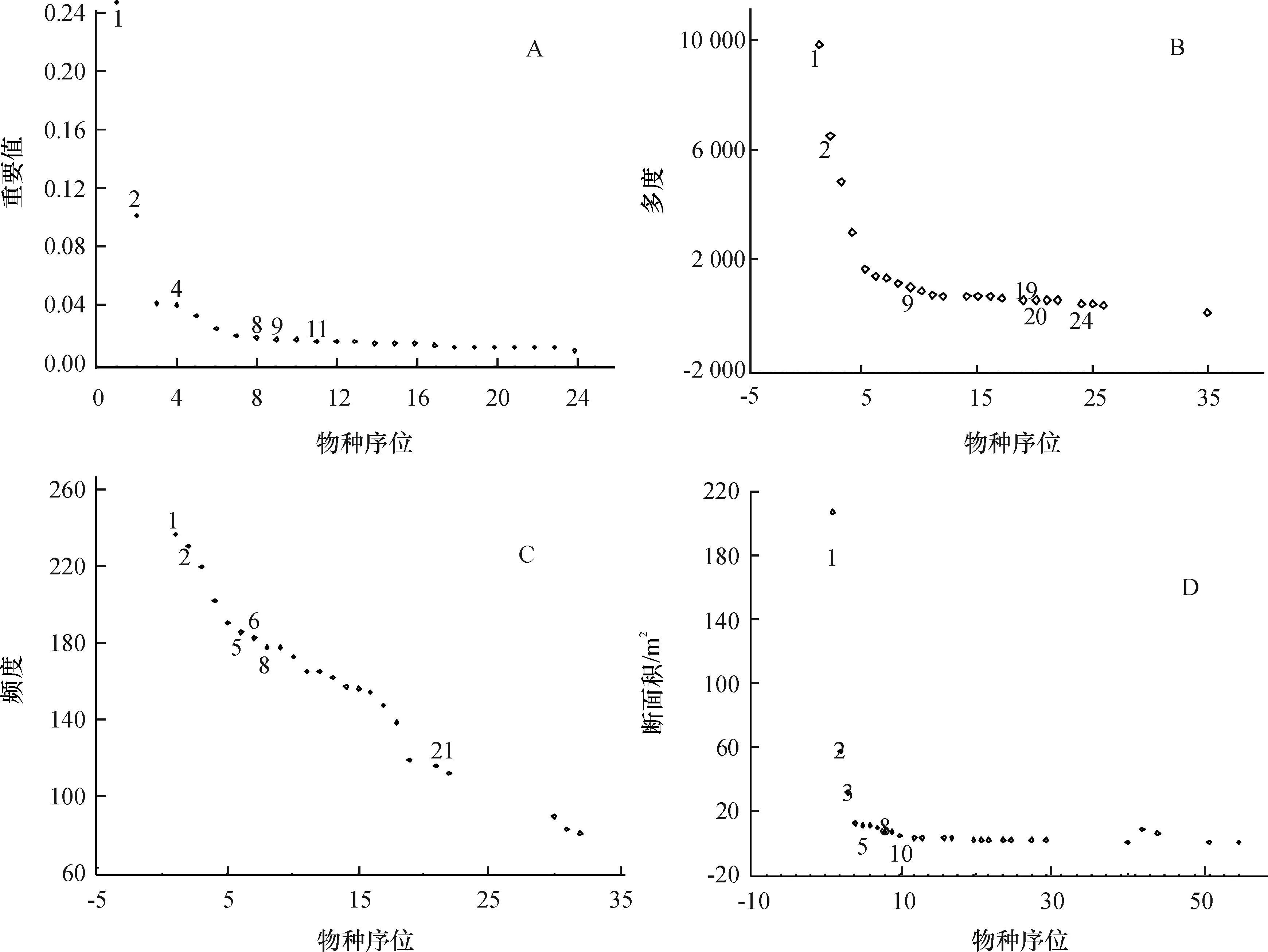
图1优势种序位曲线
Fig.1The sequence curves of dominant species
群落中相对其他树种有着足够的优势,是群落的建群种,木荷也有着较明显的优势,是排列第2的优势树种,而其他4个筛选树种的重要值也较大,虽然有个别的树种重要值更大,但主要是灌木和小乔木,其生物量和碳储量并不占据优势,综合各种考虑因素,最终筛选出6个优势树种进行更深入研究。
3.2木材密度在6个树种间的差异
对生材密度、含水率和基本木材密度在树种间的变化进行单因素方差分析和LSD检验,从表1可以看出,生材密度、含水率和基本木材密度在6个树种间都有极显著的差异。生材密度最大的是红锥,为1.18 g·cm-3,最小的是华润楠,为0.6 g·cm-3,平均值最大是米槠,最小是黄樟,变异系数最小是木荷,最大的是黄樟,米槠、栲、红锥这3种壳斗科的植物生材密度的最大、最小值和平均值都较大,木荷处于中等,而华润楠和黄樟这2种樟科植物的生材密度相对较小,同科植物生材密度变化不明显,不同科的植物生材密度有明显变化。含水率最大为黄樟,达到282.12%,最小也是黄樟,为36.35%,平均值最大和最小分别是米槠和红锥,变异系数最大和最小分别是黄樟和米槠,可以看出米槠的含水率普遍较其他树种高,而红锥含水率较其他树种低,同科植物在含水率上并不趋向一致,含水率低可能是红锥木质优良的原因之一。基本木材密度最大是黄樟(0.69 g·cm-3),最小也是黄樟(0.18 g·cm-3),平均值最大是红锥,最小是黄樟,变异系数最小是米槠,最大是黄樟,米槠、栲、红锥这3种壳斗科植物基本木材密度普遍偏大,而2种樟科植物普遍偏小,与生材密度变化一致。总的来看,红锥含水率低,基本木材密度高,具有较好的材质,而黄樟含水率高,木材密度低,其材质较差。3种壳斗科植物的种内木材密度变异相对较小,相对稳定,比较适合遗传选育,而2种樟科植物的种内木材密度变异较大,不利于遗传选育。
表1木材密度在树种间的变化
Tab.1Differences in wood density among tree species
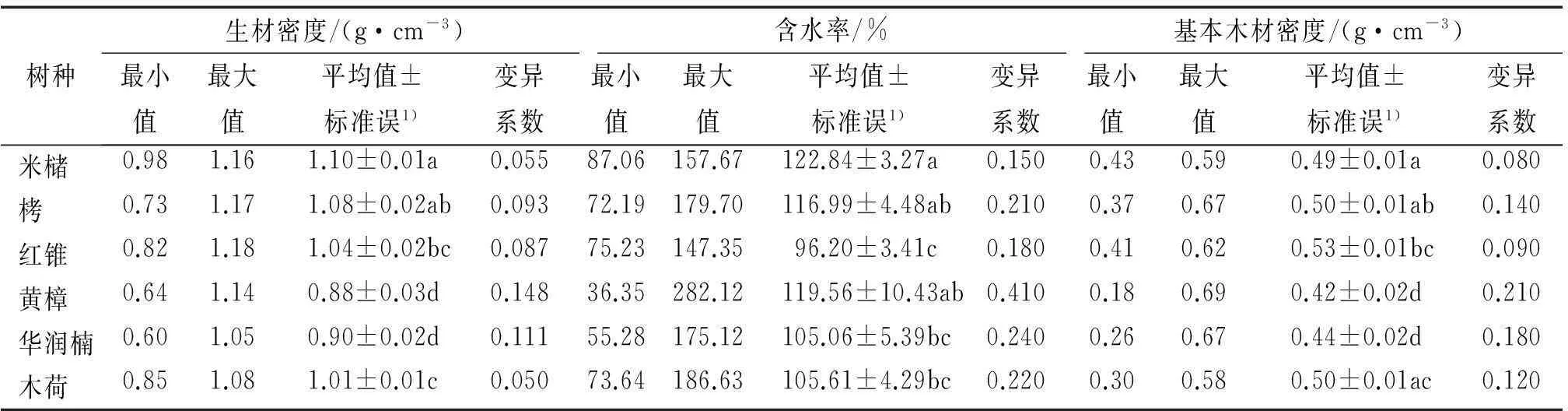
树种生材密度/(g·cm-3)含水率/%基本木材密度/(g·cm-3)最小值最大值平均值±标准误1)变异系数最小值最大值平均值±标准误1)变异系数最小值最大值平均值±标准误1)变异系数米槠0.981.161.10±0.01a0.05587.06157.67122.84±3.27a0.1500.430.590.49±0.01a0.080栲0.731.171.08±0.02ab0.09372.19179.70116.99±4.48ab0.2100.370.670.50±0.01ab0.140红锥0.821.181.04±0.02bc0.08775.23147.3596.20±3.41c0.1800.410.620.53±0.01bc0.090黄樟0.641.140.88±0.03d0.14836.35282.12119.56±10.43ab0.4100.180.690.42±0.02d0.210华润楠0.601.050.90±0.02d0.11155.28175.12105.06±5.39bc0.2400.260.670.44±0.02d0.180木荷0.851.081.01±0.01c0.05073.64186.63105.61±4.29bc0.2200.300.580.50±0.01ac0.120
1)同列数据后,凡是具有一个相同小写字母者,表示差异不显著(LSD检验,P>0.05)。
3.3径阶对木材密度的影响
用方差分析F检验以及LSD多重比较分析了木材密度在径阶间的差异。F检验结果得出生材密度在径阶间有显著差异的树种有栲和红锥,基本木材密度在径阶间有显著差异的树种有栲和华润楠,而6个树种的含水率在径阶间均无显著差异。从表2可以看出,米槠的生材密度和基本木材密度从径阶1~径阶4间都有缓慢上升趋势,在径阶5处下降。栲的生材密度和基本木材密度从径阶1~径阶5不断增加,而含水率不断降低。红锥的生材密度和基本木材密度总体呈上升趋势,在径阶4有一个波谷。3种壳斗科植物的生材密度和基本木材密度从小径阶向大径阶变化过程中总体呈上升趋势,其中,栲的基本木材密度和红锥的生材密度上升幅度最大,最为明显,径阶的增长意味着树龄的增大,可以推断,随着树龄的增大,3个壳斗科植物的生材密度和基本木材密度也有增加的趋势。黄樟的木材密度和含水率从径阶1~径阶4分别呈上升和下降趋势。华润楠的木材密度虽然总体上升,但波动较大。樟科植物木材密度总体变化趋势也是随径阶增加而变大,但变化较小,波动较大。木荷的生材密度和含水率总体有微弱上升,基本木材密度有微弱下降。总的来说,除了木荷的基本木材密度外,6个树种的生材密度和基本木材密度随径阶的增加都有上升的趋势,有的上升幅度较大,有的较小,有的波动较大,而含水率没有一定的变化规律。
3.4坡向对木材密度的影响
用单因素方差分析探讨坡向对木材密度的影响(表3),6个树种的生材密度、含水率和基本木材密度在阴坡和阳坡间的差异均不显著。其中,含水率的变异系数普遍偏大,基本木材密度次之,生材密度最小;从含水率来看,3种壳斗科植物阳坡的含水率均大于阴坡,木荷也有同样情况,2种樟科植物则相反,阴坡大于阳坡。而在生材密度和基本木材密度方面,6个树种在坡向的变化上并不一致,有的阴坡较大,有的阳坡较大。坡向反映了环境中的水热情况,因此间接表明了6个物种的生材密度、基本木材密度对水热条件不够敏感。
表26个树种木材密度在径阶间的差异1)
Tab.2Differences in wood density among diameter grades for six tree species
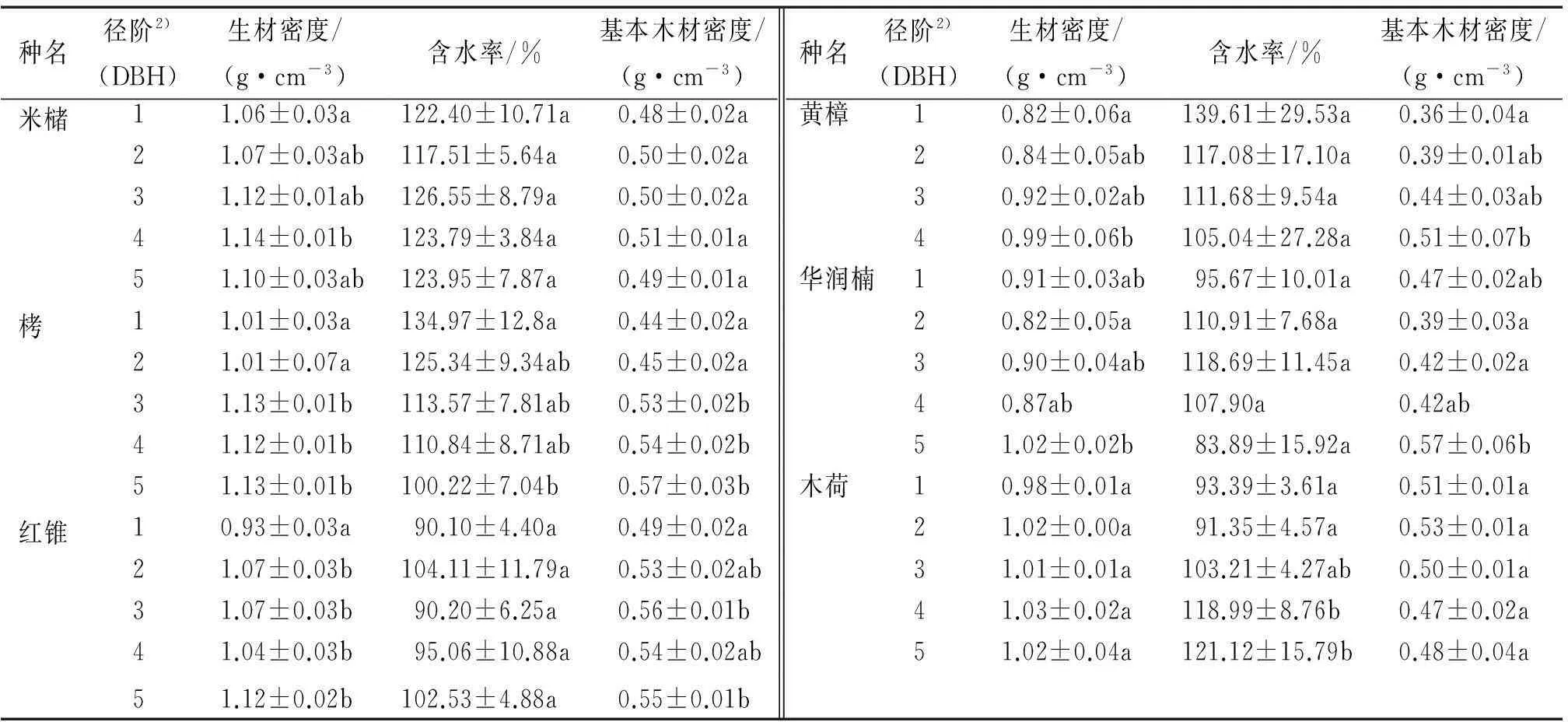
种名径阶2)(DBH)生材密度/(g·cm-3)含水率/%基本木材密度/(g·cm-3)种名径阶2)(DBH)生材密度/(g·cm-3)含水率/%基本木材密度/(g·cm-3)米槠11.06±0.03a122.40±10.71a0.48±0.02a黄樟10.82±0.06a139.61±29.53a0.36±0.04a21.07±0.03ab117.51±5.64a0.50±0.02a20.84±0.05ab117.08±17.10a0.39±0.01ab31.12±0.01ab126.55±8.79a0.50±0.02a30.92±0.02ab111.68±9.54a0.44±0.03ab41.14±0.01b123.79±3.84a0.51±0.01a40.99±0.06b105.04±27.28a0.51±0.07b51.10±0.03ab123.95±7.87a0.49±0.01a华润楠10.91±0.03ab95.67±10.01a0.47±0.02ab栲11.01±0.03a134.97±12.8a0.44±0.02a20.82±0.05a110.91±7.68a0.39±0.03a21.01±0.07a125.34±9.34ab0.45±0.02a30.90±0.04ab118.69±11.45a0.42±0.02a31.13±0.01b113.57±7.81ab0.53±0.02b40.87ab107.90a0.42ab41.12±0.01b110.84±8.71ab0.54±0.02b51.02±0.02b83.89±15.92a0.57±0.06b51.13±0.01b100.22±7.04b0.57±0.03b木荷10.98±0.01a93.39±3.61a0.51±0.01a红锥10.93±0.03a90.10±4.40a0.49±0.02a21.02±0.00a91.35±4.57a0.53±0.01a21.07±0.03b104.11±11.79a0.53±0.02ab31.01±0.01a103.21±4.27ab0.50±0.01a31.07±0.03b90.20±6.25a0.56±0.01b41.03±0.02a118.99±8.76b0.47±0.02a41.04±0.03b95.06±10.88a0.54±0.02ab51.02±0.04a121.12±15.79b0.48±0.04a51.12±0.02b102.53±4.88a0.55±0.01b
1)某物种同一指标不同径阶的该列数据后,凡是具有一个相同小写字母者,表示差异不显著(LSD检验,P>0.05);2)径阶1为5 cm≤DBH<10 cm,径阶2为10 cm≤DBH<20 cm,径阶3为20 cm≤DBH<30 cm,径阶4为30 cm≤DBH<40 cm,径阶5为DBH≥40 cm。
表36个树种木材密度在坡向间的差异性1)
Tab.3Differences in wood density among slope aspects for six tree species

树种坡向生材密度/(g·cm-3)含水率/%基本木材密度/(g·cm-3)平均值±标准误变异系数P平均值±标准误变异系数P平均值±标准误变异系数P米槠阴坡1.10±0.020.060.91122.30±4.460.140.870.49±0.010.080.99阳坡1.10±0.010.05123.38±4.930.150.49±0.010.10栲阴坡1.07±0.020.080.70111.86±5.900.200.260.51±0.020.160.47阳坡1.09±0.030.09122.11±6.680.210.49±0.020.12红锥阴坡1.02±0.030.100.2594.79±4.520.160.720.53±0.020.110.47阳坡1.06±0.020.0797.31±5.070.190.54±0.010.06黄樟阴坡0.91±0.040.120.42127.10±11.440.280.520.40±0.020.130.55阳坡0.86±0.040.16113.29±16.840.510.43±0.040.28华润楠阴坡0.87±0.030.090.34109.49±10.860.300.510.42±0.020.140.30阳坡0.91±0.030.13101.99±5.450.190.46±0.030.22木荷阴坡1.02±0.010.030.45104.90±5.590.210.870.50±0.010.100.72阳坡1.01±0.010.06106.33±6.700.240.49±0.020.12
1)P>0.05表示差异不显著(F检验)。
3.5树木性状与木材密度的相关关系
树木性状因子如胸径、树高等与木材密度的相关关系采用非参数Spearman相关分析进行处理,结果(表4)表明,除了华润楠的胸径和树高、米槠的胸径以及黄樟的树高,其他树种的胸径和树高与生材密度总体呈正相关关系,其中,栲、红锥和木荷的树木性状因子与生材密度的相关关系较强;基本木材密度方面,只有栲的树高和胸径以及黄樟的胸径与基本木材密度有显著正相关关系,其他关系不显著,这与生材密度不形成一致;含水率只与栲、木荷的胸径有显著关系,其他不显著。总的来看,栲的木材密度与胸径、树高的关系显著,随着胸径的增大,树高的增加,栲的生材密度和基本木材密度也随着变大,含水率降低;而华润楠的木材密度与胸径、树高的关系均不显著,没有明显变化规律。红锥、木荷的胸径、树高与生材密度有正相关关系,而与基本木材密度的关系却不显著,这可能是受含水率影响造成的。随着胸径和树高的增大,生材密度也增大,但生材密度增大的一部分原因可能是含水率的提高,而干物质却没有显著变化,因此生材密度增大了,基本木材密度却没有显著增大。另外,可能是生材密度增大部分被分为含水率和干物质增大两部分,各自增加的量变小了,因此含水率和基本木材密度与胸径、树高的相关性不显著。
表4树木性状与木材密度的相关关系1)
Tab.4Correlation between tree traits and wood density

树种指标生材密度含水率基本木材密度RPRPRP米槠胸径0.36070.05020.06000.75280.17010.3688树高0.4183*0.02140.21500.25400.02080.9132栲胸径0.4562*0.0113-0.4433*0.01420.6365**0.0002树高0.5837**0.0007-0.32930.07560.6002**0.0005红锥胸径0.5950**0.00170.24160.24460.35950.0775树高0.4404*0.02760.22000.29070.24750.2330黄樟胸径0.4420*0.0394-0.15560.48920.5495**0.0081树高0.17670.43160.05950.79270.26330.2364华润楠胸径0.18460.41070.00730.9741-0.00730.9741树高0.14320.52500.00230.99200.01430.9498木荷胸径0.4199*0.02090.5327**0.0024-0.28610.1253树高0.5390**0.00210.29380.1150-0.03410.8579
1) *表示0.05水平显著相关,**表示0.01水平极显著相关(Spearman’s方法)。
4讨论与结论
4.1不同树种木材密度对材质的影响
树种差异对木材密度的影响很大,不同树种木材密度差异可达数倍[8]。木材密度与植物的功能性状息息相关,关系着植物体的支撑能力、水分运输、木质强度和硬度等。不同树种其生活策略也有所差别,有的树种依赖快速生长在短时间达到林冠层,得到更多的光照和雨水,这样的树种一般生命周期较短,木材密度较低。而有些树种因为生长较慢,不需要短时间吸收太多的物质,而是通过长期的物质积累成长,其木材密度一般较高,生命周期较长。本研究样地群落的建群种为米槠,是华南地区一种较为常见的先锋树种,其他5种优势种也是亚热带森林出现频率较高的树种。通过树种间的木材密度比较得出,在筛选的6个树种中,生材密度和基本木材密度平均值最大的分别为米槠和红锥,最小的均为黄樟,生材密度和基本木材密度在树种间的变化幅度为20%和21%。而含水率平均值最小的是红锥,最大的是米槠,然后是黄樟。从生材密度、基本木材密度和含水率的变化情况可以看出,红锥的木材密度大,变异系数小,含水率低,已经具备作为优质木材的条件。而黄樟的木材密度小,变异系数大,含水率高,不具有优质木材的特性。蒋燚等[21]认为红锥的木材坚硬耐腐,色泽、纹理美观,可作为高级家具、造船、车辆、工艺雕刻、建筑装修等优质用材,是华南地区重要的乡土阔叶珍贵用材和高效多用途树种。
不同树种在木材密度上的差异,为制作各种各样的木材产品提供了保证。比如,高档的木材制品通常由致密的木材制作,但提供这类木材的树种通常生长期较长,因为相同体积下其积累的物质密度较高,需要更长的周期,因此产品价格较为昂贵。而造纸、低档木材应用等方面的木材需求数量较大,因此需要生长速度快的树种,这类树种因为生长过程中单位体积需要的物质量较少,因此能够快速生长。研究木材密度在树种间的变化,对于人工造林来说,可以根椐不同的用材目的筛选出合适的树种,而对于天然林,木材密度的树种间差异体现出不同树种的生活策略,这可为森林生态系统的运行机制提供科学参考和合理建议。
4.2不同树种木材密度对径阶和坡向的响应
在人工林木材密度的研究中,树龄是一个重要的影响因素[22],有研究认为,相同树种,树龄越大,其木材密度也越大[23]。但是在天然林中,树龄较难获得,树木的径阶则可直接测得,而径阶的变化与树龄密切相关,通过研究树木的径阶对其木材密度的影响来反映树龄与木材密度的关系是一种可行的办法。本研究中仅有栲的木材密度在径阶间有显著差异,其他树种差异性不够显著。除了木荷的基本木材密度外,6个树种的生材密度和基本木材密度随径阶的增加都有上升的趋势,但上升的幅度不同。木材密度随着径阶变大而变大的情况,原因可能有两种:一种是树木主动增加木材密度,在树木生长过程中,躯干承受的压力增大,需要强度和硬度更大的材质来负担,植物体主动提供更多的物质来增加木材密度,以适应这种变化;另一种是树木被动增加木材密度,随着树木的生长,躯干受到更大的压力,这种压力使得树木细胞变得越来越紧密,一些空隙也被挤压填补,木材密度增大。
地形因子对植物的生长、生理具有重要影响,特别是阳坡(南坡)、阴坡(北坡)雨热分配的差异,对植物的生理结构,包括其木材密度也具有一定的作用[24]。在本研究中,筛选的6个树种的生材密度、含水率和基本木材密度在阴坡和阳坡间的差异均不显著,6个树种木材密度在坡向的变化上并不一致,有的阴坡较大,有的阳坡较大。其原因可能有两种:一种是因为6个树种的木材密度对水热条件不够敏感;另外一种是因为坡向对木材密度的变化并不是主导因子,坡向对木材密度的影响较小,而体现不出一定的规律性。
参考文献:
[1]BERGMAN R, CAI Z, CARLL C G. Wood handbook: Wood as an engineering material[M]. Madison: United States Department of Agriculture, Forest Service, Forest Products Laboratory, 2010: 1-2.
[2]WU R. Microstructural study of sanded and polished wood by replication[J]. Wood Sci Technol, 1998, 32(4): 247-260.
[3]GJERDRUM P, EIKENES B. A model for spatial wood density gradients in Norway spruce stems and stochastic between-stem dissimilarities for basic and dry density[J]. Wood Sci Technol, 2014, 48(1): 71-84.
[4]MACHADO J S, LOUZADA J L, SANTOS A J A, et al. Variation of wood density and mechanical properties of blackwood (AcaciamelanoxylonR. Br.)[J]. Mater Design, 2014, 56: 975-980.
[5]MOLTEBERG D, HOIBO O. Modelling of wood density and fibre dimensions in mature Norway spruce[J].Can J Forest Res, 2007, 37(8): 1373-1389.
[6]POORTER L, WRIGHT S J, PAZ H, et al. Are functional traits good predictors of demographic rates? Evidence from five neotropical forests[J]. Ecology, 2008, 89(7): 1908-1920.
[7]罗云建. 华北落叶松人工林生物量碳计量参数研究[D]. 北京:中国林业科学研究院, 2007: 52-53.
[8]中国林业科学研究院木材工业研究所. 中国主要树种的木材物理力学性质[M]. 北京: 中国林业出版社, 1982.
[9]侯振宏. 中国林业活动碳源汇及其潜力研究[D]. 北京:中国林业科学研究院, 2010: 78-79.
[10]朱江玲,石岳,方乐祺,等. 中国主要树种木材物理力学属性的地理格局及其环境控制[J]. 中国科学(生命科学), 2015, 45(1): 56-67.
[11]徐金梅,吕建雄,鲍甫成,等. 祁连山青海云杉木材密度对气候变化的响应[J]. 北京林业大学学报, 2011, 33(5): 115-121.
[12]刘福涛. 立地对马占相思人工林生长和材性的影响[D]. 福州:福建农林大学, 2007: 25-29.
[13]PATINO S, LLOYD J, PAIVA R, et al. Branch xylem density variations across the Amazon Basin[J]. Biogeosciences, 2009, 6(4): 545-568.
[14]WIEMANN M C, WILLIAMSON G B. Geographic variation in wood specific gravity: Effects of latitude, temperature, and precipitation[J]. Wood Fiber Sci, 2002, 34(1): 96-107.
[15]鲍甫成,江泽慧. 中国主要人工林树种木材性质[M]. 北京: 中国林业出版社, 1998.
[16]成俊卿. 木材学[M]. 北京: 中国林业出版社, 1985.
[17]徐明锋,胡砚秋,李文斌,等. 土壤养分对亚热带天然林物种分布的影响[J]. 中南林业科技大学学报, 2014, 34(9): 91-97.
[18]潘秋荣,梁佰华,陈小芸. 广东东源康禾省级自然保护区野生药用植物资源调查[J]. 广东林业科技, 2012, 28(4): 25-30.
[19]林中大. 广东康禾自然保护区植物区系研究[J]. 广西林业科学, 2007, 36(2): 75-77.
[20]胡传双,苏志尧,云虹,等. 流体静力称衡法测定生长锥取样木材密度的改进方法[J]. 华南农业大学学报, 2010, 31(3): 105-108.
[21]蒋燚,李志辉,朱积余,等. 红锥家系木材密度等物理性状的遗传及变异性分析[J]. 中南林业科技大学学报, 2012, 32(11): 9-13.
[22]刘青华,张蕊,金国庆,等. 马尾松年轮宽度和木材基本密度的种源变异及早期选择[J]. 林业科学, 2010, 46(5): 49-54.
[23]NGUYEN H, FIRN J, LAMB D, et al. Wood density: A tool to find complementary species for the design of mixed species plantations[J]. Forest Ecol Manag, 2014, 334: 106-113.
[24]王秀花,陈柳英,马丽珍,等. 7年生木荷生长和木材基本密度地理遗传变异及种源选择[J]. 林业科学研究, 2011, 24(3): 307-313.
【责任编辑李晓卉】
Wood densities of six hardwood tree species in Eastern Guangdong and Influencing factors
XU Mingfeng, KE Xiandong, ZHANG Yi, WANG Yongqiang, SU Zhiyao
(College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou 510642,China)
Abstract:【Objective】 This study aims to provide the theoretical and data supports for the breeding of wood density and carbon sink accounting by analyzing the effects of different impact factors on wood density. 【Method】Six tree species were selected through the analysis of communities in sample plots of Kanghe nature reserve, and their wood densities were measured. This study analyzed the effects of different species, different diameter grades and different slope aspects on wood density, and the correlation between tree traits and wood density using a variety of statistical softwares. 【Result】 There were significant differences in wood density among different tree species. The highest averages of green densities (1.10 g·cm-3), basic densities (0.53 g·cm-3) and the moisture contents (122.84%) were found in Castanopsis carlesii, C. hystrix and C. carlesii respectively. There was no significant difference in wood density among different diameter grades for all species except C. fargesii. There was no significant difference in wood density among different slope aspects for all species. The nonparametric Spearman’s rank correlation analysis showed that both diameter at breast height and tree height were significantly correlated with green density but not with basic density. 【Conclusion】C. hystrix produces high quality wood with low moisture content and high basic density. Wood density has a tendency to rise with the increase of diameter grade,but has no regular change with the change of slope aspect. The relationship between diameter at breast height, tree height and green density, and the relationship between diameter at breast height, tree height and basic density are different probably because green density is affected by the moisture content.
Key words:wood density; green density; basic density; diameter grade; slope aspect; hardwood tree; influencing factor
中图分类号:S718.5
文献标志码:A
文章编号:1001- 411X(2016)03- 0100- 07
基金项目:广东省林业科技创新专项(2010KJCX012-02, 2011KJCX031-02)
作者简介:徐明锋(1988—),男,博士研究生,E-mail: mingfengxu2008@163.com;通信作者:苏志尧(1963—),男,教授,博士, E-mail: zysu@scau.edu.cn
收稿日期:2015- 09- 07优先出版时间:2016-04-15
优先出版网址:http://www.cnki.net/kcms/detail/44.1110.s.20160415.1555.038.html
徐明锋, 柯娴氡, 张毅,等.粤东6种阔叶树木材密度及其影响因子研究[J].华南农业大学学报,2016,37(3):100- 106.
猜你喜欢
防护林科技(2022年2期)2022-04-25 23:47:32
安徽农学通报(2020年17期)2020-10-09 10:46:23
湖南林业科技(2017年1期)2017-02-06 05:28:55
电脑知识与技术(2016年28期)2016-12-21 11:09:59
农业与技术(2016年19期)2016-12-12 03:18:40
出版科学(2016年5期)2016-11-10 06:47:04
防护林科技(2016年8期)2016-08-29 01:28:17
贵州科学(2016年2期)2016-06-13 00:58:58
读书(2016年5期)2016-05-21 22:36:29
武夷学院学报(2015年3期)2015-07-18 11:03:47
