
‘渝城1号’核桃矿质元素周年吸收规律研究
2024-04-12 02:51魏立本黄小辉王玉书唐佳佳杨华均冯大兰邹孝文
西南大学学报(自然科学版) 2024年4期
魏立本, 黄小辉, 王玉书, 唐佳佳,杨华均, 冯大兰, 邹孝文
1. 重庆市林业科学研究院/三峡库区森林生态保护与恢复重庆市重点实验室,重庆 400036;2. 城口县林业局,重庆 城口 405900
核桃(JuglansregiaL.)属胡桃科胡桃属落叶乔木, 果实含多种氨基酸和磷、铁、锰、锌等人体必需的矿质营养, 脂肪含量在60%以上, 其中不饱和脂肪酸占90%左右, 具有很高的营养价值和保健作用[1], 备受国内外消费者的青睐.我国核桃栽植面积较大, 尤其云南、四川、重庆等山区因退耕还林栽植了大量核桃, 然而, 受山区交通、地形等条件限制, 核桃施肥难度大, 成本高, 很多核桃园在栽植后基本不施肥, 不仅核桃产量低下, 更使土壤养分只出不进, 土壤肥力严重下降.有条件的核桃园为了提高产量大量施肥, 但缺乏科学的施肥技术指导, 肥料乱用滥施现象普遍, 不仅造成资源浪费, 还导致土壤退化、营养失衡.因此, 为减少化肥用量和提高肥效, 节约资源和保护环境, 我国各地皆在倡导核桃配方施肥[2-3].
科学地配方施肥需掌握目标作物的营养需求规律.由于树木生长周期长, 树体大, 对其营养需求规律方面的研究相对较难, 所以已有的报道, 如苹果[4-5]、茶树[6]、荔枝[7]等, 仅研究了矿质营养元素在不同生育期的变化规律, 对具体吸收量和分配特征等研究还相对较少.在20世纪90年代, 赵明范[8]对核桃叶片中各营养元素质量分数的季节性变化规律进行了分析, 初步发现核桃叶片的N, P, K质量分数随生育期变化逐渐降低, 随后还有学者[9-11]研究了晚实核桃叶片中矿质元素质量分数的动态变化.在营养分配方面, 梁智等[12]以5年生‘温185’早实核桃为试验对象, 研究各器官矿质元素质量分数及分配特性, 发现N, P, K三大元素在核桃各器官组织中的分配存在较大差异, 赵瑞芬等[13]对9年生‘辽宁1号’早实核桃的研究结果表明, 该品种核桃对N, P, K的吸收累积量比例也有很大不同.就目前来看, 国内已有的核桃矿质营养质量分数动态变化及分配特征等研究, 能为核桃施肥管理提供一定的科学指导, 但还远远不够.对于核桃这种产果量大的树种, 需肥种类和数量更多, 还需详细了解核桃各器官生物量构成、矿质营养累积特性及需求量等, 才能更好地指导对核桃的科学施肥.
‘渝城1号’是重庆市的主栽核桃品种, 具有树势强旺、适应性强的特点, 在重庆市多数区县均有分布, 总面积超过6.67万hm2, 但因施肥等重要管理技术不到位, 干果平均亩产量不到30 kg, 经济效益低下, 严重影响山区农民增收.因此, 本研究以‘渝城1号’核桃为研究对象, 采用彻底刨根和分解取样的方法, 分别在不同生育期采集核桃的器官样品, 分析其生物量和N, P, K, Ca, Mg质量分数, 探究核桃对5种矿质元素的周年吸收动态与需求规律, 以期为‘渝城1号’核桃的科学高效施肥提供依据.
1 材料与方法
1.1 试验材料与设计
试验地点位于重庆市城口县高观镇施礼村(31°50′57″N, 108°54′32″E), 海拔870 m, 年均气温13.7 ℃, 降雨量1 361.1mm, 无霜期235 d, 年均日照时数1 357 h.土壤为黄砂壤, 土层厚度60 cm, 土壤理化性质如表1所示.试验核桃树于2016年12月选用规格基本一致的1年生嫁接苗定植, 品种为‘渝城1号’.2017-2018年, 每年施有机肥(有机质46.4%, N 0.973%, P2O51.96%, K2O 2.23%)和复合肥(N∶P2O5∶K2O=15∶15∶15), 有机肥在11月上旬施入, 施肥量2 kg/株, 复合肥在次年4月上旬施入, 施肥量0.2 kg/株.施肥方式采用沟施, 沿树冠边缘开沟, 深度20 cm.2019年初, 选择生长均匀且健康的核桃树50株, 标记作为采样单株.在各生育期随机抽取5株进行全株挖掘, 按照根、茎、营养枝、果枝、叶、果实分开采集样品.具体生育期为: 休眠期(12月上旬-次年3月上旬)、展叶抽梢期(3月中旬-4月中旬)、开花坐果期(4月下旬-5月上旬)、果实膨大及硬核期(5月中旬-8月中旬)、果实成熟期(8月下旬-9月下旬)、落叶期(10月上旬-11月下旬).各生育期的采样时间为: T1(1月16日), T2(3月24日), T3(4月28日), T4(7月17日), T5(9月8日), T6(11月3日).所有样品洗净后, 进行烘干、粉碎, 用于生物量和矿质元素的测定.

表1 试验地土壤理化性质
1.2 测定指标及方法
生物量测定: 采用烘干法, 在80 ℃烘箱烘至恒质量, 用千分之一电子天平测定其干质量.
矿质元素含量测定: 样品经硫酸-高氯酸消煮后, 分别采用凯式定氮法、钼锑钪比色法和火焰光度计测定N, P, K质量分数; 经硝酸-高氯酸消煮后, 采用原子吸收法测定Ca, Mg质量分数.
矿质元素累积量的计算及曲线拟合参照张文君[14]的方法, 具体如下:
相关矿质元素累积量计算公式:
a矿质元素总累积量(g)=b干物质累积量(g)×c矿质元素含量(%)
针对生长曲线特征, 用Logistic曲线拟合核桃的矿质元素累积量随生长天数增长的动态变化, 方程模型为:
Y=A/(1+Be-kX)
式中:Y为各矿质营养元素的累积量;X为当年生长的天数(按采样当天计算);k为生长速度参数;A为矿质元素积累量极限参数;B为常数尺度.方程拟合参数采用非线性最小二乘法估算.
1.3 数据分析
利用Microsoft Excel进行数据统计, Origin 2021软件进行作图, 用SPSS 20.0统计软件进行方差分析和Logistic曲线的拟合, 采用LSD进行多重比较, 显著水平为p<0.05.
2 结果与分析
2.1 核桃各器官干物质累积量动态变化
如图1所示, 在整个生育期内, 各器官干物质累积量均表现出相似的增长趋势.根、茎、营养枝、果枝、叶的干物质从T2开始逐渐增加, 直到T5后树体生长基本停止, 干物质基本不再增加.核桃果实在T3后逐渐生长, 随着果实膨大, 干物质迅速增加, 直到T4后, 核桃果实的干物质累积量明显高于其他器官.从整个生育期看, 核桃开花坐果到果实膨大这一阶段是干物质快速增加的时期, 增加的干物质主要储存在果实、根和茎中.

图1 核桃各器官干物质累积量动态变化(平均值±标准差)
2.2 核桃各器官N质量分数及分配特征
由表2可以看出, 核桃根、茎、营养枝、果枝的N质量分数总体均呈先升后降的趋势, 其中茎、营养枝、果枝的N质量分数均在T2时最高, 随后逐渐下降, 在T5时降至最低.根的N质量分数在T4时最高.叶和果实N质量分数随着核桃的生长逐渐下降, 在T5时降至最低.在N的分配上, 在T3前, N更多地分配在根、茎、叶中, 尤其以根部的累积量最大.在T3后, N在果实中的分配急剧增加, 在T5时, 果实中N的累积量达到5.63 g, 占全株的44.43%.
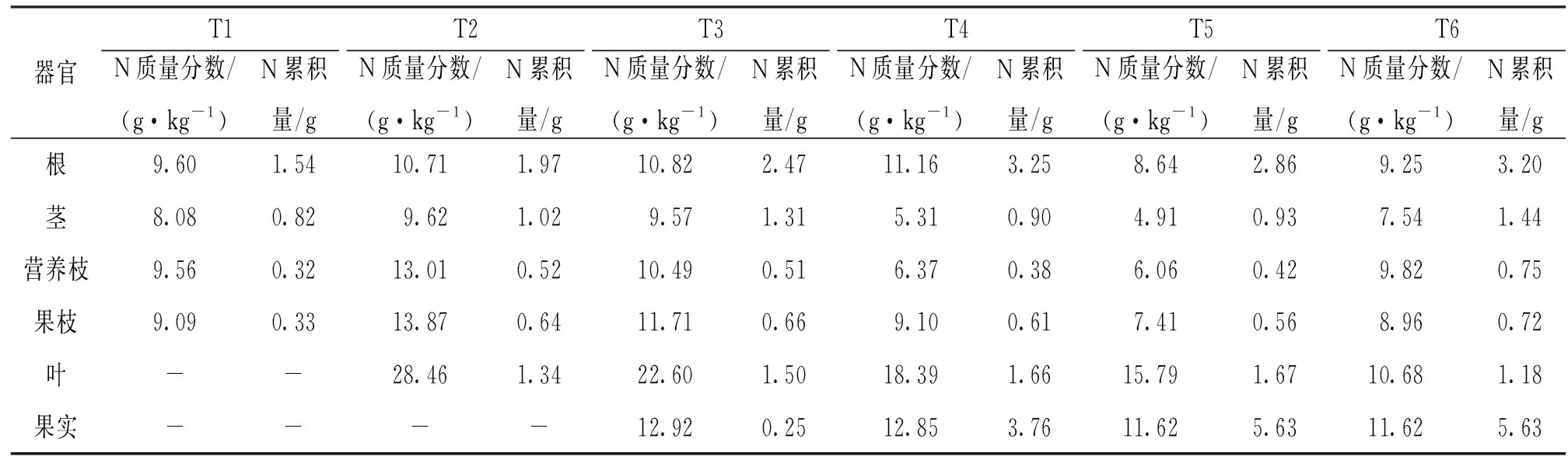
表2 核桃各器官N质量分数及累积量动态变化
2.3 核桃各器官P质量分数及分配特征
由表3可以看出, 与N的变化趋势相似, 核桃根、茎、营养枝及果枝的P质量分数总体呈先升后降的趋势, 在T2时最高, 然后逐渐下降, 直到T5时降至最低.叶和果实的P质量分数随生育期的推进呈下降趋势, 在T6时降至最低.P的分配上, 在T3前, P主要分配于根和叶中, 并以根的累积量最大.在T3后, P在果实中的分配逐渐增多, 直到T5时, 果实中的P累积量达到1.12 g, 占全株的40.73%.
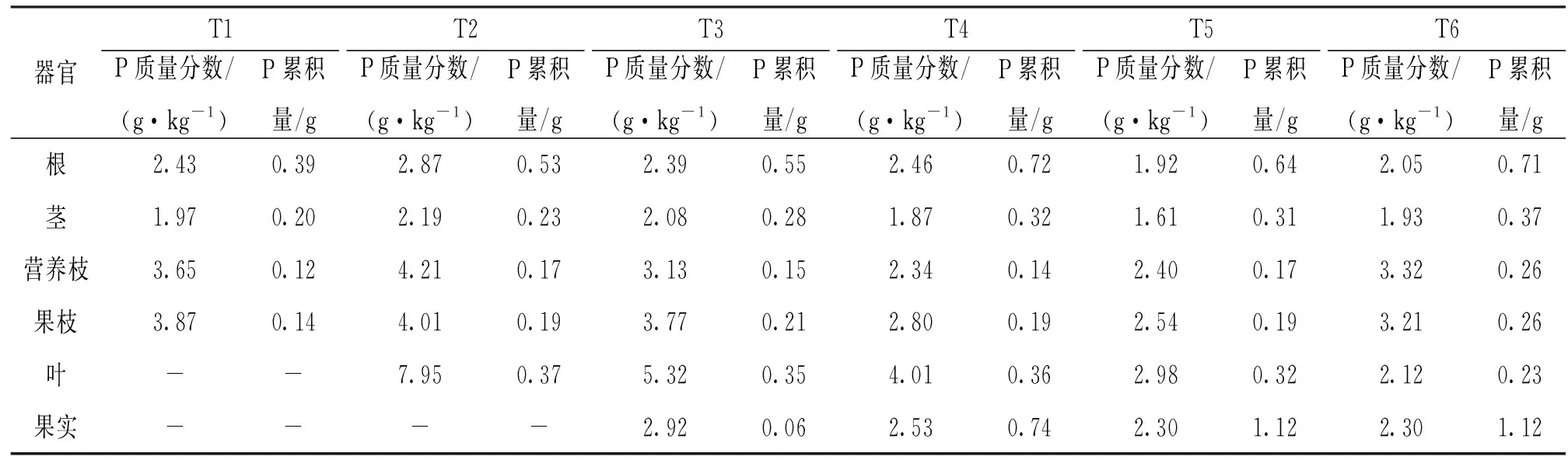
表3 核桃各器官P质量分数及累积量动态变化
2.4 核桃各器官K质量分数及分配特征
由表4可以看出, 核桃根、茎、营养枝、果枝的K质量分数与N, P的变化趋势相似, 总体呈现出先增加后降低的趋势, 其中茎、营养枝、果枝K质量分数在T2达到最高, 根部的K质量分数在T4达到最高.叶和果实中K质量分数的变化趋势一致, 随生育期的推进逐渐下降, 直至T6时降至最低.K的分配上, 在T3前, K在根和叶中分配较多, 尤其在根中的累积量最大.进入T3后, K在果实中分配增加.在T5时, 果实中K的累积量达到最高, 为6.62 g, 占全株的62.22%.

表4 核桃各器官K质量分数及累积量动态变化
2.5 核桃各器官Ca质量分数及分配特征
由表5可以看出, 整个生育期内, 核桃根和果枝的Ca质量分数呈先降后升的变化趋势, 在展叶抽梢期降至最低.相反, 茎和营养枝的Ca质量分数呈先升后降的变化趋势, 在果实膨大期达到最高, 随后迅速下降, 并在成果实熟期和落叶期保持相对稳定.叶片和果实的Ca质量分数变化趋势相反, 其中叶片的Ca质量分数随生育期的推进不断升高, 而在果实中则不断下降.果实膨大期之前, Ca在核桃各器官中的分配以根、茎较多, 尤其是根部的累积量最大.果实膨大期后, 随着果实的逐渐成熟, 果实中的Ca累积量逐渐增多, 这一阶段Ca更多地累积在根、叶和果实中.由于Ca的移动性很弱, 在落叶期无法回流至树体, 所以叶片中的Ca质量分数为全株最高, 占全株的24.82%.

表5 核桃各器官Ca质量分数及累积量动态变化
2.6 核桃各器官Mg质量分数及分配特征
由表6可以看出, 整个生育期内, 根和茎的Mg质量分数随生育期的推进呈现明显的波动变化.营养枝和果枝的Mg质量分数均呈现先降后升的变化趋势, 并在落叶期达到最高.相反, 叶片的Mg质量分数呈先升后降的变化趋势, 在果实成熟期降至最低, 而果实中的Mg质量分数随着生育期推进不断降低.从休眠期到开花坐果期, 核桃各器官中的Mg元素主要累积在根部.开花坐果期后, 叶片和果实的Mg累积量逐渐增多, 并且该时期果实和叶片中的Mg累积量明显高于其他器官.总体上, 在果实成熟期, 果实的Mg累积量达到全株最高, 为0.77 g, 占全株的30.20%.

表6 核桃各器官Mg质量分数及累积量动态变化
2.7 核桃对N, P, K, Ca, Mg的吸收累积量动态变化
从全株角度看, 核桃N, P, K, Ca, Mg累积量动态变化对不同元素的吸收累积规律不尽相同.从图2可以看出, 核桃对各类矿质元素的吸收累积量变化趋势大致相同, 均表现为休眠期到开花坐果期缓慢上升, 而开花坐果以后, 随着果实的生长, 树体中N, K, Ca的吸收累积量快速上升, 在果实膨大期后逐渐减缓, 直至落叶期仍略有增加.在整个生育期内, 各矿质元素的吸收累积量从大到小依次为: N, Ca, K, P, Mg(p<0.05).通过对核桃N, P, K, Ca, Mg的吸收累积量进行Logistic曲线拟合分析可知(表7), 五者的R2均较高, 尤其N和P均达0.99以上, 拟合程度较优.根据拟合曲线得出核桃对N, P吸收量最大的时间接近, 均在开花坐果期, 相应的累积量分别为7.09 g/株和1.67 g/株, 而对K, Mg的吸收量最大的时间在果实膨大期前, 相应的累积量为6.21 g/株和1.60 g/株.对Ca的吸收量最大的时间在果实膨大期, 相应的累积量为7.77 g/株.
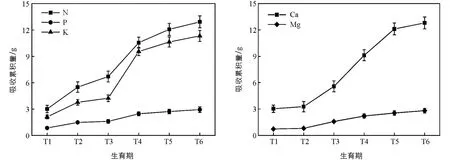
图2 核桃N, P, K, Ca, Mg累积量动态变化(平均值±标准差)

表7 核桃N, P, K, Ca, Mg累积量的Logistic方程及其特征值
3 结论
纵观‘渝城1号’核桃的整个生育期, 各器官中的N, P, K质量分数均在当年开始生长(展叶抽梢期)时最高, 尤其叶片中的营养元素质量分数显著高于其他器官, 原因是该阶段核桃吸收的养分主要用于叶片展开和迅速扩大.因此, 在核桃的展叶抽梢期前应及时进行N, P, K等大量元素的供应, 以防因叶片发育不良影响光合作用, 导致树体衰弱, 影响随后的开花坐果.叶片展开后, 随着叶片和新梢的持续旺长及花果的发育, 各器官的N, P, K质量分数逐渐下降.在开花坐果到果实膨大期间, 核桃茎、营养枝、果枝和叶片的N, P, K质量分数降幅最大, 但果实的N, P, K质量分数变化不大, 说明果实的生长需消耗大量矿质营养, 而茎、枝、叶的减少部分则转移到了果实中.核桃整个树体对N, P, K的吸收累积增长最快(生长拐点)的时间也在开花坐果期和果实膨大期前.可见核桃在开花坐果以后, N, P, K肥的及时补充很有必要, 这也是保证幼果膨大及提高产量、品质的关键, 这与袁紫倩等[15]对薄壳山核桃的研究结论一致.核桃果实采收后到落叶前期, 根、茎、营养枝、果枝的N, P, K质量分数出现回升现象, 而叶片中仍在继续下降, 原因是N, P, K在植物体内具有较强的移动性, 此时从叶片回流至树体内进行营养储存, 保证树体来年的营养供应, 这也是果树的普遍现象[16-17], 所以果实采收后也应当注意N, P, K肥的适当补充.此外, 整个生育期内, 核桃的根、果实中N, P, K元素的累积量最高, 尤其开花坐果后, 果实中的累积量急剧上升, 其中K的累积量占全株60%以上, 推测这与果实发育过程中淀粉和脂肪合成需要大量K有关.
Ca, Mg是植物必需的中量元素, 需求量仅次于N, P, K.但由于核桃果实对Ca的吸收量较大, 加上整个树体各器官Ca质量分数均较高, 所以‘渝城1号’核桃对Ca的需求量显著高于P, K, Mg, 在树体中Ca的总累积量接近于N.从展叶抽梢期到果实成熟期, 核桃叶片、营养枝和果枝的Ca, Mg质量分数均呈上升趋势, 并且整个树体在开花坐果期到果实膨大期对Ca, Mg的吸收累积增长最快, 表明该阶段树体对Ca, Mg的需求较大, 因此为保证核桃生长和果实品质, 应在开花坐果期到果实膨大期补充适量的钙镁肥.果实成熟后, 各器官Ca, Mg质量分数均趋于稳定, 而叶片的Ca质量分数仍在继续上升, 原因可能是Ca的移动性很弱, 很难从叶片转移到树体中, 而Mg是可移动元素, 随着叶片衰老, 不断从叶片向其他器官回流, 所以叶片的Mg质量分数有明显下降.在Ca, Mg的累积分配上, 整个生育期内根部的Ca累积量最多, 这可能也与Ca的移动性弱有关.在展叶抽梢期和开花坐果期, Mg在根部的累积量最多, 但随后更多地转移到了果实中, 这与Mg容易从成熟组织转移至生长活跃的器官有关, 尤其在幼果膨大时, 还会加速果实周围叶片和枝条的Mg元素向其转移[18].
综上, 开花坐果期到果实膨大期是核桃干物质量增加最快和矿质元素吸收量最大的时期, 因此该阶段是核桃施肥的关键时期.在开花坐果期应注意补充N, P肥, 在幼果期应补充K, Mg肥, 在果实膨大期应补充Ca肥, 以达到肥料高效利用和核桃对矿质元素高效吸收的目的, 进而促进核桃增产.
猜你喜欢
园艺与种苗(2023年6期)2023-07-03
草地学报(2022年9期)2022-09-29
农业科技通讯(2022年6期)2022-06-21
新疆农业科学(2020年2期)2020-03-12
中成药(2018年11期)2018-11-24
山东农业科学(2018年7期)2018-10-22
现代园艺(2017年21期)2018-01-03
江苏农业科学(2014年9期)2014-11-15
安徽农学通报(2014年3期)2014-03-11
食品工业科技(2014年9期)2014-03-11
