
中华绒螯蟹不同家系幼蟹生长性能及性早熟比较试验
2024-04-08 03:35:06高志宝张婷李旭光邓燕飞周军
水产养殖 2024年4期
高志宝,张婷,李旭光,邓燕飞,周军*
(1.江苏海洋大学海洋科学与水产学院,江苏 连云港 222005;2.江苏省淡水水产研究所,农业农村部淡水虾蟹遗传育种与养殖重点实验室,江苏 南京 210017)
中华绒螯蟹(Eriocheir sinensis),又名河蟹,隶属于节肢动物门,软甲纲,十足目,弓蟹科,绒螯蟹属,广泛分布于我国东南沿海以及长江、瓯江、辽河等内陆水域,是我国重要的经济养殖蟹类,2022 年全国河蟹养殖面积53 万hm2,产量80 万t[1]。河蟹养殖周期主要包括苗种繁育、幼蟹培育和成蟹养殖3 个阶段,其中幼蟹质量直接决定着成蟹的养殖产量与效益。幼蟹培育阶段,1 龄幼蟹性腺如提前发育达到性成熟,会导致性早熟。性早熟1 龄蟹生长速度快,个体大,性腺发育早,与正常幼蟹抢食饵料,但体质量增长缓慢甚至停滞,绝大多数在翌年死亡,无法用于后续成蟹的养殖,给养殖生产带来巨大的损失[2]。因此,控制降低1 龄蟹性早熟比例,是幼蟹培育阶段的关键。
河蟹性早熟,受到内源种质遗传和外源因素双重调控[3]。目前对于河蟹性早熟的研究,主要集中在温度、盐度、溶解氧等环境因素,以及饵料、养殖密度、地理群体等养殖条件对性早熟的影响方面,而有关内源遗传对1 龄蟹性早熟的影响研究,尚存在较大争议[4]。在同一养殖条件下培育蟹种,部分幼蟹个体仍然会性早熟,暗示性早熟的发生与内源遗传有关。现开展中华绒螯蟹不同家系幼蟹生长性能及性早熟的比较试验,分析影响中华绒螯蟹生长性能与性早熟的关键因素,为河蟹遗传选育提供参考依据。
1 材料与方法
1.1 时间与地点
2022 年5—12 月。试验地点位于江苏省镇江扬中市江苏省淡水水产研究所基地。
1.2 材料
选取8 个150 m2长方形池塘。池塘为平底,每池用10 kg 漂白粉消毒,清除野杂鱼、水生昆虫等。在进排水口设立孔径0.18 mm 网栅,过滤鱼卵等敌害生物。所有网箱均放置在水体联通的池塘围隔中,保证养殖水环境条件均一。4 月中旬应开始种植水草,以水花生为主,伊乐藻为辅,其中采用毛竹和线绳设置水花生带,带宽80~100 cm,沿池塘四周种植伊乐藻,养殖前期水草覆盖率为40%,中后期为70%。河蟹亲本选自江苏省淡水水产研究所扬中基地的成蟹养殖池塘。雌、雄亲本均于2021 年11 月,运至江苏省淡水水产研究所盐城射阳中华绒螯蟹繁育基地,进行人工繁殖,定向构建8 个全同胞家系,分别为ES1—ES8,每对家系亲本(雌雄1∶1)分别在100 L 的网箱中单独交配和育苗。
1.3 试验方法
1.3.1 幼蟹培育
将培育出来的大眼幼体,运送至江苏省淡水水产研究所扬中基地进行蟹种培育试验,按1 个家系投放0.5 kg 大眼幼体,1 个家系1 个池塘进行定向蟹种培育。幼蟹培育全程采用相同配合饲料投喂,饲料日投喂量约为池内蟹体质量的5%,投喂配合饲料粒径,随着蟹种的生长逐步加大,每天投喂1,次,全池泼洒。根据天气和摄食情况,适时调整饲料投喂量。每天早晚巡塘,观察幼蟹的摄食、活动、蜕壳等情况,检查池埂是否渗漏、拦隔设施是否严密、防止幼蟹逃逸等。严防野杂鱼苗进入培育池,及时清除进入培育池中的青蛙、黄鳝等;及时捞除池中飘浮的脏污,清除池埂杂草,做好塘口档案记录。
1.3.2 生长性能与性早熟测定
在90、120 和150 日龄,分别采集每个家系30 只个体(雌雄各15 只),剔除断肢个体,采用电子游标卡尺测量幼蟹壳长和壳宽(精确到0.01 mm),采用电子天平称量幼蟹体质量(精确到0.1 g)。10 月份以后,采用地笼集中捕捞规格蟹种和性早熟个体。性早熟判别主要依据外部副性特征,其中雄蟹主要依据交接器是否突出和硬化、大螯绒毛覆盖面积和长度;雌蟹主要依据腹脐形状、腹脐绒毛长度和壳的颜色。结合性腺解剖,雌蟹卵巢发育到II~III期,雄蟹精巢发育到III 期,筛选确定性早熟个体。后期将池内水花生分段集中,先用网箱集中捕捞,再用地笼冲水诱捕,最后清塘捕捞,统计每个家系的幼蟹产量和性早熟蟹比例。
1.4 数据分析
采用Graphpad Prism 8.3 软件对试验数据进行统计分析。统计幼蟹不同生长阶段壳长、壳宽和体质量数据,结果用(平均值±标准误)表示。对数据进行方差齐性检验,采用单因素方差Tukey 法,进行多重比较分析,P<0.05 为差异显著,P<0.01 为差异极显著。
2 试验结果
河蟹不同家系幼蟹生长阶段的壳长、壳宽和体质量随机抽样测试结果见表1。河蟹不同家系正常与性早熟蟹种体质量比较见图1。河蟹不同家系正常与性早熟蟹种数量比较见图2。
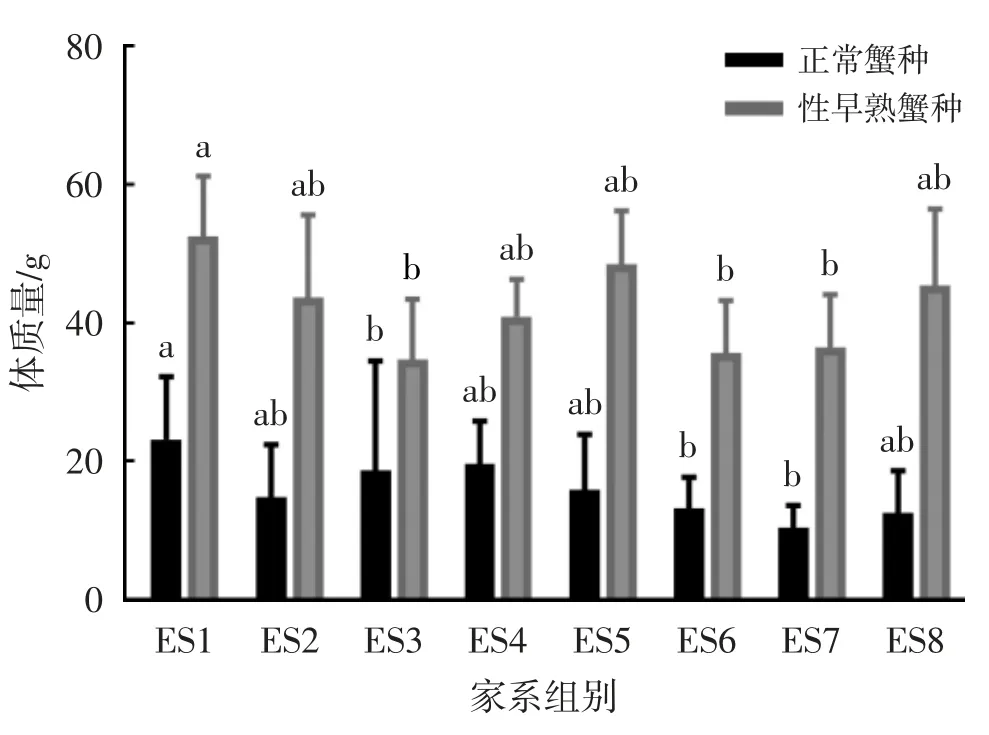
图1 河蟹不同家系正常与性早熟蟹种体质量比较
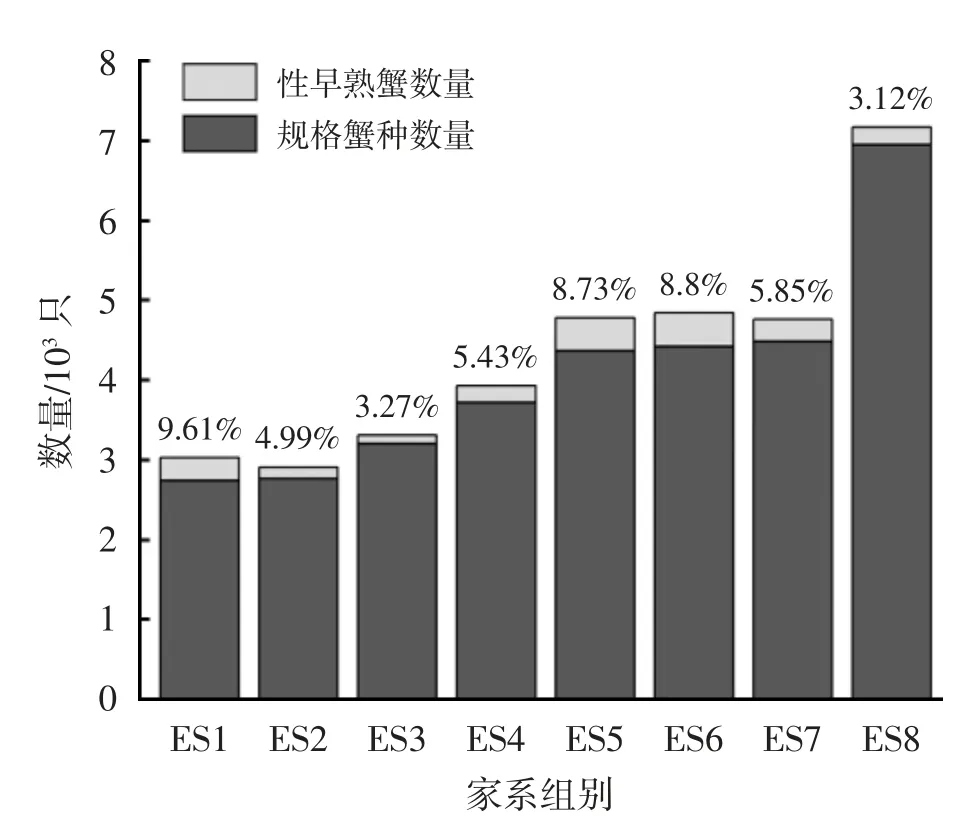
图2 河蟹不同家系正常与性早熟蟹种数量比较
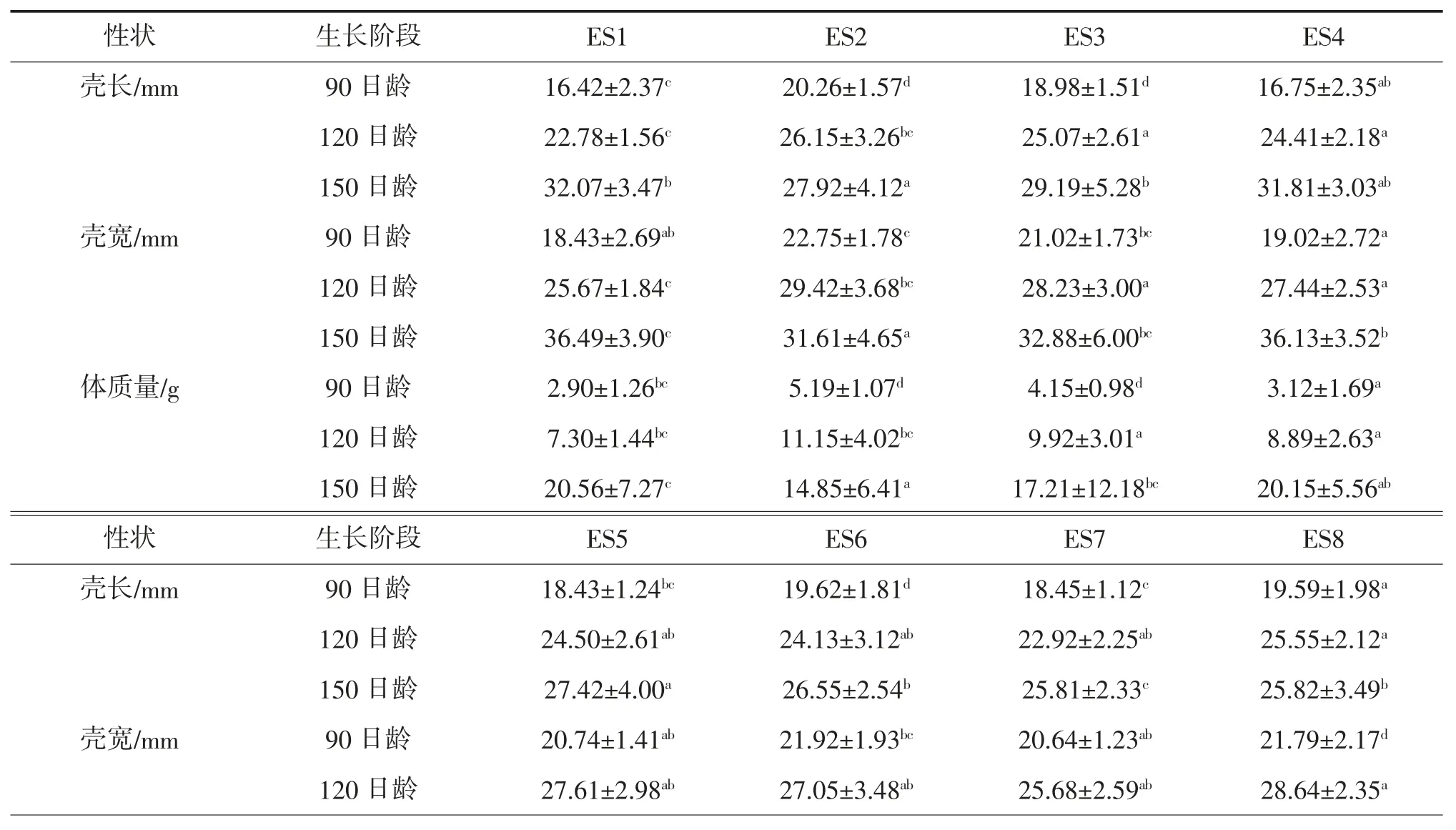
表1 河蟹不同家系间幼蟹生长阶段的壳长、壳宽和体质量随机抽样测试结果①
3 讨论
3.1 河蟹不同家系的生长性能
本试验结果表明,河蟹不同家系间的生长性状存在差异,这与文献[5]关于长江水系河蟹家系蟹种阶段生长性能的研究结果相似。为辨识内源与外源因素对河蟹生长的影响,本试验整个蟹种培育阶段,不同家系间养殖水温、pH 值、氨氮等指标相同,养殖密度相同、蟹种规格均一。因此确定河蟹不同家系间的生长性状差异,主要由内源遗传因素导致。
家系选育是目前遗传育种常用的方法之一,通过建立家系,比较不同家系生长性状,可获得性状稳定的家系[6]。郑岩等[7]在对1 龄性早熟河蟹关于早熟性状遗传能力方向的研究时发现,早熟蟹回捕率与扣蟹规格呈极显著正相关,与扣蟹回捕率、总回捕率呈极显著负相关。王海宁等[8]报道了1 龄早熟河蟹家系后代的体质量和甲壳宽,均显著低于2 龄成熟家系,选育河蟹家系的生长性能和免疫抗病生理指标优于未选育群体。
3.2 河蟹不同家系的性早熟差异
相同养殖环境条件下的不同家系河蟹性成熟比例存在差异,表明种质遗传因素贡献度较大。本试验中,河蟹不同家系性早熟数量比例为3.12%~9.61%,这与许郑超等[9]研究的性早熟数量比例为2.22%~7.19%相近。河蟹性成熟包括生理性成熟、功能性成熟和形态性成熟,首先是性腺发育进入生理性成熟,然后螯肢绒毛、腹节、步足刚毛、体色第二性征的形态性成熟,最后是交接器骨质化、降河洄游交配的功能性成熟[10]。影响甲壳类性腺发育进程的因素,主要包括光照、温度、营养和遗传等。Li等[11]研究表明,弱光照强度和间歇饥饿,不仅可以有效降低幼蟹的性早熟率,还可以激发幼蟹的补偿生长潜能。杜海燕等[12]比较分析了积温、饵料和遗传3 个因素对性早熟的影响,结果表明,河蟹不同家系的组间变异明显大于组内变异,种质遗传因素对幼蟹性早熟的影响最大。本试验中,养殖环境相同,养殖密度与饵料投喂相似,不同家系河蟹性早熟比例差异的最大变量,主要来自不同家系的种质遗传背景。文献[13]通过组织化学和分子生物学等方法,筛选发现性早熟个体能量代谢通路中关键基因的突变,提高了能量摄入和转化效率,加速了性腺发育。而本试验中,部分家系性早熟比例较高,是否与能量代谢通路有关,还有待于进一步研究。
猜你喜欢
当代水产(2022年8期)2022-09-20 06:46:34
今日农业(2021年1期)2021-03-19 08:35:38
农业与技术(2018年14期)2018-11-12 11:22:08
湖北畜牧兽医(2018年1期)2018-01-26 15:38:15
渔业致富指南(2016年12期)2016-11-11 07:07:21
渔业致富指南(2016年12期)2016-11-11 07:07:15
广东海洋大学学报(2015年4期)2016-01-13 08:39:30
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:04
中药与临床(2015年5期)2015-12-17 02:39:29
首都医科大学学报(2015年4期)2015-12-16 13:00:08
