
月均气温对三叶青林下容器栽培苗光合特性及其块根总黄酮含量的影响
2024-04-08 07:22章健敏林国卫娄俊
浙江农业科学 2024年3期
章健敏,林国卫,娄俊
(1.玉山县气象局,江西 上饶 334700;2.上饶师范学院 生命科学学院,江西 上饶 334001;3.上饶市广信区气象局,江西 上饶 334100)
三叶青 (TetrastigmahemsleyanumDiels et Gilg)为葡萄科崖爬藤属植物,主要分布于浙江、江西、福建、湖南、湖北、广东、广西等地,为我国特有珍稀药用植物[1]。三叶青新鲜或干燥块根有清热解毒、消肿止痛、化痰散结的功效,常用于小儿高热惊风、百日咳、毒蛇咬伤等治疗[2];三叶青干燥全草也具有清热解毒、消肿止痛、活血祛风等功效,也可用于流行性感冒、肝炎、泌尿系统结石、跌打损伤等治疗[3]。光合作用是绿色植物基本的生命活动之一,也是植物次生代谢物形成的基础[4]。研究表明,温度可显著影响植物的生长发育,限制其光合作用,从而决定后续植物次生代谢物的含量,是一个重要的生态因子[5]。在全球气温变化加剧的背景下,极端气温频繁出现对植物造成逆境胁迫及导致的经济损失已引起世界广泛关注,也是我国农林业生产中最主要的气象自然灾害之一[6]。目前,对三叶青的研究主要集中在扦插育苗[7]、化学成分[8]、药理作用[9]、组培快繁[10]等方面,而关于三叶青光合生理的研究仅限于设施栽培条件[11]、不同光强处理[12]、不同氮形态[13]和生物炭肥[14]等影响因子,关于三叶青总黄酮含量的影响因子也仅限于产地[15]、生长环境[16]、干燥与储存方式[17]、取材部位与处理[18]、光质[19]、遮阴[20]、海拔高度[21]等,而关于气温对三叶青光合生理及其块根总黄酮含量的影响尚未见报道。本研究以三叶青林下容器栽培三年生扦插苗为研究对象,在2015—2019年研究五府山月均气温对其光合生理及其块根总黄酮含量的影响,旨在找出三叶青林下容器栽培的光合生理规律及其块根总黄酮含量变化,为三叶青的林下容器生态栽培提供理论依据。
1 材料与方法
1.1 材料
五府山竹林下三叶青三年生控根容器栽培苗[22]。
1.2 方法
1.2.1 五府山月均气温对三叶青容器栽培苗光合特性和叶绿素荧光参数的影响
2015—2019年每年每月5日、10日、15日、20日和25日选取五府山竹林下三叶青三年生控根容器栽培苗顶端形状相近的叶片,用Li-6400光合仪测定光合特性参数和遮光(采用锡箔纸进行遮光)条件下的叶绿素荧光参数。测量前,需先使用锡箔纸对待测叶片进行30 min的暗适应处理。然后先用检测光照射测定叶片的初始荧光(Fo),再用饱和白光光照1 s测最大荧光(Fm)。最后,打开内源光光化30 min,测定光下最小荧光(F′o)、光下最大荧光(F′m)及稳态荧光(Fs)[23]。光合特性参数包括净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等参数及环境CO2浓度(仪器进气口的CO2浓度),并计算气孔限制值(Ls)、水分利用效率(WUE)和瞬时羧化速率(CUE),其公式分别为Ls=1-Ci/Ca,WUE=Pn/Tr,CUE=Pn/Ci。CUE用于估测RuBPCase(1,5-二磷酸核酮糖羧化酶)的活性。使用开放式气路,CO2浓度为400 μmol·mol-1。同日,选择生长健壮、长势相近的五府山竹林下三叶青三年生控根容器栽培苗顶部和中间部分的叶片,采用便携式叶绿素测定仪SPAD-502Plus对五府山竹林下三叶青三年生控根容器栽培苗叶片叶绿素含量 SPAD 值进行测定[24]。上述每日试验重复3次。每月5日、10日、15日、20日和25日测定的光合特性、叶绿素荧光参数和叶绿素含量SPAD 值,取其平均值,算出五府山竹林下三叶青容器栽培苗光合特性、叶绿素荧光参数和叶绿素含量SPAD 值的每月平均值。
1.2.2 五府山月均气温对三叶青容器栽培苗块根总黄酮含量的影响
2015—2019年每年每月5日、10日、15日、20日和25日选取五府山竹林下三叶青三年生控根容器栽培苗块根,采用阮明颖[15]的方法测定其总黄酮含量。上述每日试验重复3次。每月5日、10日、15日、20日和25日测定的总黄酮含量,取其平均值,视为五府山竹林下三叶青容器栽培苗块根的每月平均总黄酮含量。
1.2.3 五府山2015—2019年每日气温的调取和数据统计分析
利用五府山2015—2019年区域站气温数据,算出2015—2019年逐年及5年的各月平均气温,结合五府山竹林下三叶青容器栽培苗光合特性、叶绿素荧光参数和叶绿素含量SPAD 值的每月平均值以及五府山竹林下三叶青容器栽培苗块根的每月平均总黄酮含量。
利用SPSS 20.0软件,采用单因素方差分析法(one-way ANOVA)进行2015—2019年五府山月均气温下三叶青林下容器栽培苗光合生理及其块根总黄酮含量的显著性分析。
2 结果与分析
2.1 2015—2019年五府山月均气温对三叶青林下容器栽培苗叶绿素含量的影响
2015—2019年五府山月均气温对三叶青林下容器栽培苗叶绿素含量的影响见表1。由表1可知,2015—2019年,五府山1月的气温最低,2—6月气温逐渐提升,7—8月气温达到最高值,9—12月气温逐渐下降。由表1可知,在2015—2019年的每年1—12月,五府山三叶青林下容器栽培苗叶绿素含量的SPAD值也随着气温的变化而显著发生变化;在1—6月气温逐渐提高,五府山三叶青林下容器栽培苗叶绿素含量的SPAD值也显著提高(P<0.05);气温在7—8月达到最高值,五府山三叶青林下容器栽培苗叶绿素含量的SPAD值也达到最大值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗叶绿素含量的SPAD值也随之显著下降(P<0.05)。

表1 2015—2019年五府山月均气温对三叶青林下容器栽培苗叶绿素含量的影响Table 1 Effect of monthly average temperature in Wufu Mountain from 2015 to 2019 on chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg seedlings grown in containers under the forest
2.2 2015—2019年五府山月均气温对三叶青林下容器栽培苗光合特性的影响
2015—2019年五府山五年月均气温对三叶青林下容器栽培苗光合特性的影响见表2。由表2可知,在2015—2019年的每年1—12月,五府山三叶青林下容器栽培苗光合特性也随着气温的变化而显著发生变化。在1—6月气温逐渐提高,五府山三叶青林下容器栽培苗净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、气孔限制值(Ls)和瞬时羧化速率(CUE)也显著提高(P<0.05);气温在7—8月达到最高值,净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、气孔限制值(Ls)和瞬时羧化速率(CUE)也达到最大值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、气孔限制值(Ls)和瞬时羧化速率(CUE)也随之显著下降(P<0.05)。此外,在1—6月气温逐渐提高,五府山三叶青林下容器栽培苗胞间CO2浓度(Ci)和水分利用效率(WUE)也显著下降;气温在7—8月达到最高值,胞间CO2浓度(Ci)和水分利用效率(WUE)也达到较低值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗胞间CO2浓度(Ci)和水分利用效率(WUE)也随之显著提高(P<0.05)。
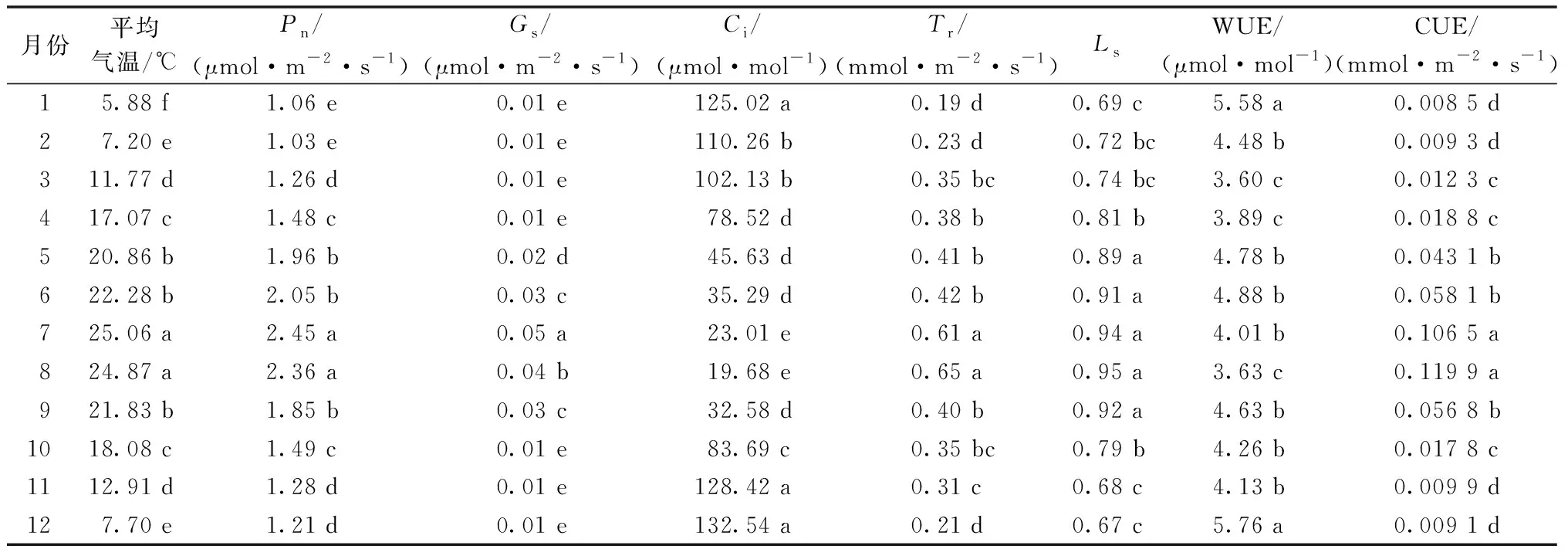
表2 2015—2019年五府山月均气温对三叶青林下容器栽培苗光合特性的影响Table 2 Effect of monthly average temperature in Wufu Mountain from 2015 to 2019 on photosynthetic characteristics of Tetrastigma hemsleyanum Diels et Gilg seedlings grown in containers under the forest
2.3 2015—2019年五府山月均气温对三叶青林下容器栽培苗叶绿素荧光参数的影响
2015—2019年五府山五年月均气温对三叶青林下容器栽培苗叶绿素荧光参数的影响见表3。由表3可知,在2015—2019年的每年1—12月,五府山三叶青林下容器栽培苗叶绿素荧光参数也随着气温的变化而变化。在1—6月气温逐渐提高,五府山三叶青林下容器栽培苗初始荧光(Fo)、最大荧光(Fm)、光下最小荧光(F′o)、光下最大荧光(F′m)、稳态荧光(Fs)、PSⅡ最大光能转化效率(Fv/Fm)、PSⅡ潜在光化学效率(Fv/Fo)、PSⅡ实际光化学效率(ΦPSⅡ)、开放的PSⅡ反应中心捕获激发能效率(F′v/F′m)、光化学淬灭系数(qP)也显著提高,而非光化学淬灭系数(NPQ)显著下降(P<0.05);气温在7—8月达到最高值,初始荧光(Fo)、最大荧光(Fm)、光下最小荧光(F′o)、光下最大荧光(F′m)、稳态荧光(Fs)、PSⅡ最大光能转化效率(Fv/Fm)、PSⅡ潜在光化学效率(Fv/Fo)、PSⅡ实际光化学效率(ΦPSⅡ)、开放的PSⅡ反应中心捕获激发能效率(F′v/F′m)、光化学淬灭系数(qP)也达到最大值,非光化学淬灭系数(NPQ)达到最低值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗初始荧光(Fo)、最大荧光(Fm)、光下最小荧光(F′o)、光下最大荧光(F′m)、稳态荧光(Fs)、PSⅡ最大光能转化效率(Fv/Fm)、PSⅡ潜在光化学效率(Fv/Fo)、PSⅡ实际光化学效率(ΦPSⅡ)、开放的PSⅡ反应中心捕获激发能效率(F′v/F′m)、光化学淬灭系数(qP)也随之显著下降,而非光化学淬灭系数(NPQ)显著上升(P<0.05)。
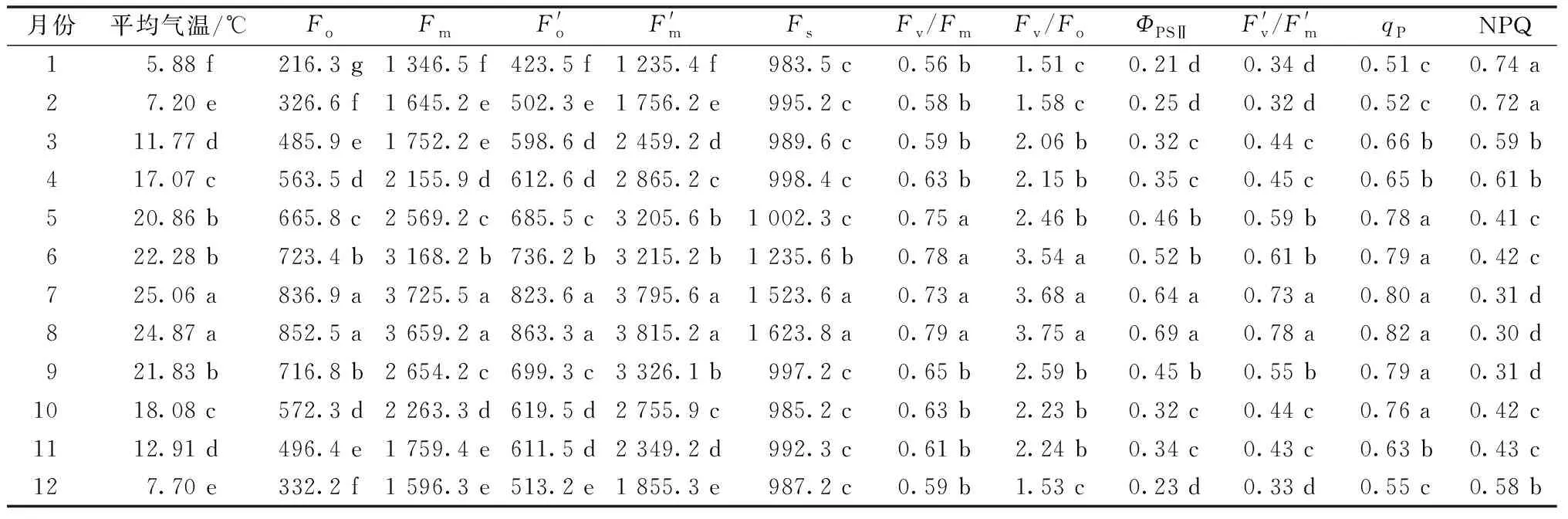
表3 2015—2019年五府山月均气温对三叶青林下容器栽培苗叶绿素荧光参数的影响Table 3 Effect of monthly average temperature in Wufu Mountain from 2015 to 2019 on chlorophyll fluorescence parameters of Tetrastigma hemsleyanum Diels et Gilg seedlings grown in containers under the forest
2.4 2015—2019年五府山月均气温对三叶青林下容器栽培苗块根总黄酮含量的影响
2015—2019年五府山五年月均气温对三叶青林下容器栽培苗块根总黄酮含量的影响见表4。由表4可知,在2015—2019年的每年1—12月,五府山三叶青林下容器栽培苗块根总黄酮含量也随着气温的变化而变化。在1—6月气温逐渐提高,五府山三叶青林下容器栽培苗块根总黄酮含量也显著提高;气温在7—8月达到最高值,块根总黄酮含量也达到最大值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗块根总黄酮含量也随之显著下降(P<0.05)。
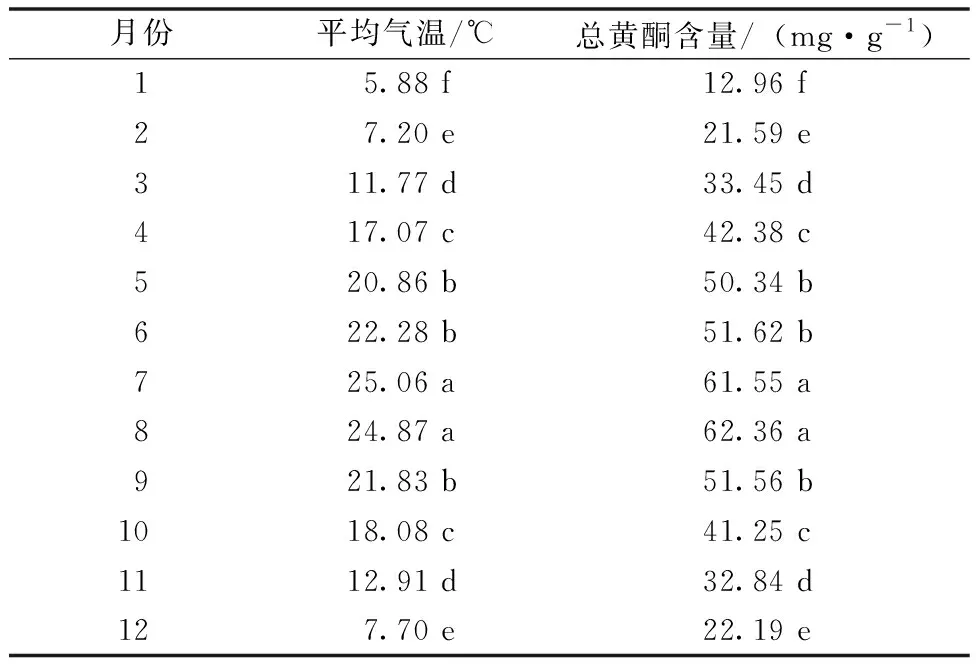
表4 2015—2019年五府山月均气温对三叶青林下容器栽培苗块根总黄酮含量的影响Table 4 Effect of monthly average temperature in Wufu Mountain from 2015 to 2019 on total flavone content of root tuber of Tetrastigma hemsleyanum Diels et Gilg seedlings grown in containers under the forest
3 讨论
植物的光合作用受外界多重因子的影响,其中温度是影响植物光合作用的重要因子之一[25]。有研究表明,温度会显著影响植物的叶绿素含量[26]。温度对龙葵幼苗叶绿素含量的影响,发现随着温度降低,龙葵幼苗叶绿素含量显著降低。黄艳花等[27]研究了温度对裂片石莼叶绿素含量的影响,发现培养的裂片石莼的叶绿素含量随着温度升高而显著增加。王璐等[28]探究了温度升高对甜椒叶绿素含量的影响,发现升温可显著增加其叶绿素含量。本试验结果与上述结果一致。在2015—2019年,随着1—6月气温逐渐提高,五府山三叶青林下容器栽培苗叶绿素含量的SPAD值也显著提高(P<0.05);气温在7—8月达到最高值,五府山三叶青林下容器栽培苗叶绿素含量的SPAD值也达到最大值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗叶绿素含量的SPAD值也随之显著下降(P<0.05)。
温度也会显著抑制植物的光合特性[29]。刘念奇等[30]研究了根区温度对嫁接黄瓜苗光合特性的影响,发现随着根区温度的降低,黄瓜叶片Pn显著降低。王珣等[31]分析不同温度下甜高粱叶片光合作用的气孔限制和非气孔限制特征,发现随着温度的升高,Pn不断增大,35 ℃时Pn分别是25 ℃、30 ℃时的1.55和1.18 倍。宗毓铮等[32]研究了气温升高对北方冬小麦旗叶光合特性的影响,发现气温升高 2 ℃会使小麦孕穗期净光合速率显著增加24.7%,而对小麦拔节期与灌浆期净光合速率无显著影响。王璐等[28]探究了温度升高对甜椒光合生理的影响,发现温度升高可使甜椒Pn、Gs、Tr和WUE极显著增加,温度升高后Ci降低。本试验结果与上述结果一致。在2015—2019年,随着1—6月气温逐渐提高,五府山三叶青林下容器栽培苗净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、气孔限制值(Ls)和瞬时羧化速率(CUE)也显著提高;气温在7—8月达到最高值,净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、气孔限制值(Ls)和瞬时羧化速率(CUE)也达到最大值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、气孔限制值(Ls)和瞬时羧化速率(CUE)也随之显著下降(P<0.05)。此外,在1—6月气温逐渐提高,五府山三叶青林下容器栽培苗胞间CO2浓度(Ci)和水分利用效率(WUE)也显著下降(P<0.05);气温在7—8月达到最高值,胞间CO2浓度(Ci)和水分利用效率(WUE)也达到最低值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗胞间CO2浓度(Ci)和水分利用效率(WUE)也随之显著提高(P<0.05)。
也有研究表明,温度会显著影响植物的叶绿素荧光参数[33]。刘念奇等[30]研究了根区温度对嫁接黄瓜苗叶绿素荧光参数的影响,发现随着根区温度的降低,黄瓜叶片Fv/Fm、qP和NPQ均显著降低。马欠等[34]研究了温度对小球藻和铜绿微囊藻生长及叶绿素荧光特性的影响,发现较高的温度有利于小球藻和铜绿微囊藻Fv/Fm的提高。张振文等[35]研究了温度对木薯叶片叶绿素荧光及 Rubisco 酶的影响,发现温度升高可显著提高木薯叶片的Fv/Fm和F′v/F′m。黄艳花等[27]研究了温度对裂片石莼叶绿素荧光特性的影响,发现培养的裂片石莼的叶绿素荧光参数(Fv/Fm、F′v/F′m、qP)随着温度升高而显著增加。本试验结果与上述结果一致。在2015—2019年,随着1—6月气温逐渐提高,五府山三叶青林下容器栽培苗Fo、Fm、F′o、F′m、Fs、Fv/Fm、Fv/Fo、ΦPSⅡ、F′v/F′m、qP也显著提高(P<0.05),而NPQ显著下降(P<0.05);气温在7—8月达到最高值,Fo、Fm、F′o、F′m、Fs、Fv/Fm、Fv/Fo、ΦPSⅡ、F′v/F′m、qP也达到最大值,NPQ达到最低值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗Fo、Fm、F′o、F′m、Fs、Fv/Fm、Fv/Fo、ΦPSⅡ、F′v/F′m、qP也随之显著下降(P<0.05),而NPQ显著上升(P<0.05)。究其原因,可能是随着气温降低,三叶青林下容器栽培苗叶片PSⅡ的部分失活或受到一定伤害,原初光能转换效率下降,电子传递活性减弱,有效抑制了光合碳代谢电子供应效率,从而影响三叶青林下容器栽培苗的光合作用,并随温度的降低抑制作用加剧。但关于不同温度对不同基因型棉花叶片叶绿素荧光的研究结果以及王浩等[4]关于气温对葡萄叶片光合荧光的研究结果与本研究结果不一致,随着温度不断升高,不同基因型棉花叶片的Fv/Fm表现为先升高后降低,而Fo则表现为先降低后升高。高气温下葡萄叶片光系统Ⅱ(PSⅡ)实际光化学效率不会显著上升反而显著降低,NPQ则显著上升。
还有研究表明,生态环境特别是气温对植物的有效药用成分的积累也有显著影响。薛焱等[36]比较了不同生态环境对冷蒿总黄酮积累的影响,发现冷蒿总黄酮含量与年均气温呈正相关,年均气温是影响冷蒿总黄酮积累的主要地理因子。本试验结果与上述结果一致,在2015—2019年,随着1—6月气温逐渐提高,五府山三叶青林下容器栽培苗块根总黄酮含量也显著提高(P<0.05);气温在7—8月达到最高值,块根总黄酮含量也达到最大值;9—12月气温逐渐下降,五府山三叶青林下容器栽培苗块根总黄酮含量也随之显著下降(P<0.05)。
猜你喜欢
阅读(快乐英语中年级)(2022年11期)2022-05-30
阅读(科学探秘)(2020年8期)2020-11-06
读者·校园版(2019年24期)2019-12-10
中国果业信息(2019年1期)2019-01-05
中成药(2018年2期)2018-05-09
现代园艺(2017年19期)2018-01-19
生物学教学(2017年9期)2017-08-20
农业工程学报(2016年6期)2016-05-17
小朋友·聪明学堂(2015年8期)2015-11-30
云南中医学院学报(2014年4期)2014-07-31
