
多重空间尺度的神经表征*
2024-03-23 07:10吴文雅
生物化学与生物物理进展 2024年3期
吴文雅 王 亮**
(1)中国科学院心理健康重点实验室(中国科学院心理研究所),北京 100101;2)中国科学院大学心理学系,北京 100049)
空间表征是指对环境中物体位置和空间关系等空间知识的心理表征[1],是外界客观知识和信息的主观存储形式。空间环境包含多重尺度,小到一览无余的桌面,大到遍布世界的不同国家,对空间环境不同尺度的区分性表征对人和其他动物的生存都具有重要意义。本文结合近期研究对多重空间尺度的神经表征进行总结,对比不同尺度空间的差异,并展望未来空间尺度表征的重要问题和研究方向。
1 空间尺度的分类
心理空间根据尺度可分成4类,从小到大依次是图像空间(figural space)、单视点空间(vista space)、环境空间(environmental space)和地理空间(geographical space)[2](图1)。图像空间比人体小,不需任何移动就可以感知此类空间的属性,包括二维图片空间(pictorial space)和三维物体空间(object space),如报纸、水壶或洗手台。单视点空间与人体不相上下甚至更大,但是仍然可以在不需移动的情况下从一个位置(即“单视点”)觉知其空间属性,如电梯间、卧室或没有障碍物的旷场。环境空间比人体大,并且包含若干障碍物,不移动就无法统览全局,通过在屏障之间穿梭并整合信息才可以形成对该空间的全面认知,如有多栋建筑物的街区。地理空间远大于人体,必须通过符号化的抽象表征将其缩减或凝练为上述的图像空间,才能为人类所感知和学习,如省份、国家乃至宇宙。
不同空间尺度的表征往往服务于特定的功能需求。图像空间属于小尺度或近尺度(near scale)空间,一般与操作性行为有关,比如拧开水壶的盖子或把水壶拿到桌面上,虽然都有动作的参与,但是可能涉及到孤立物体或空间内多个物体,其中需要对多个物体进行操作的尺度属于“可触及尺度(reachable scale)”[3]。地理空间则属于超大尺度(huge scale)空间,只能通过抽象的思考才能认知,而不能真实体验。这两类空间尺度因为过小或过大,而不需要或无法实现具身导航过程。然而介于二者之间的单视点空间和环境空间则是空间导航领域重点关注的对象,这两类空间是导航行为发生的背景和基础, 也被称为“ 可导航空间(navigable space)”。

Fig. 1 Examples of psychological space at different scales图1 不同尺度心理空间的实例
多尺度空间表征的形成过程按先后顺序一般都包括感觉输入、记忆存储和调用阶段。首先需要从多模态感觉线索(如视觉、听觉、前庭觉、自体运动线索等)中感知空间信息,随后分别在短时记忆和长时记忆中创建并存储空间表征,最后这些表征将被使用和操作以指导之后的空间行为[4]。对不同尺度空间的区别表征对人和其他生活在复杂环境中的各种动物而言都具有重要意义,动物在觅食、迁徙和求偶等过程中都需要调用多尺度空间知识[5-7],而人类在进行路线规划、有目标的空间导航等活动时也同样离不开空间知识的跨尺度灵活运用,因老化或脑损伤导致的空间能力受损会明显影响患者的日常生活[8]。
2 近尺度空间的操作性表征
近尺度空间是视觉体验的主要成分,日常生活的大部分时间人们都生活在触手可及的空间范围内,如办公室桌面等,这种近尺度空间内的活动一般都是用手执行任务并且会体验到相应的视觉、触觉等知觉输入,因此也可称为可操作空间(operable space)[9]。近尺度空间是人们与物理世界直接发生交互的中心,对这种空间的表征也因此受到具体交互内容的影响,特定近尺度空间的功能比其实际的物理位置更能直接预测相似性平级关系[10],表明近尺度空间表征是以行为功用作为指导原则的。
后侧副沟(posterior collateral sulcus)、顶下沟(inferior parietal sulcus) 和顶上小叶(superior parietal lobule)3 个脑区对近尺度空间尺度的表征表现出系统性的偏好[3]。在近期的一项功能磁共振成像(functional magnetic resonance imaging,fMRⅠ)实验中,Josephs和Konkle[3]让被试分别观看单个物体、近尺度空间以及场景的图片,3种图片都引起了视皮层的显著激活,但是皮层的不同部分会对每种刺激类型表现出特定的偏好,其中近尺度空间会引起后腹侧皮层、枕顶叶以及上顶叶的明显激活,与单个物体以及场景引起的皮层激活模式差异明显。这3个脑区虽然对近尺度空间敏感,但同时也会受到近尺度空间中可操作的物体数量的影响,当空间中只包含单个物体或没有物体时,这些区域活动微弱,而当包含多物体阵列时这些区域的活动会显著增强,由此表明近尺度空间表征注重操作性和互动性[11]。
服务于操作性行为的表征需要保证足够精确,譬如只有精准地定位茶杯和水壶的位置以及二者之间的关系,才能实现用茶壶往杯子内倒水。近尺度空间的精确表征需要满足两个要求,分别是视觉坐标和运动坐标[12],要对空间内某个物体施加操作,首先需要感知物体在环境中的位置坐标,其次要基于这种视觉坐标发起运动。对物体位置的视觉空间表征往往会存在一定的误差,根据对误差的相关性分析,Yousif 和Keil[13]发现,视觉坐标的表征采取的是极坐标系,而不是笛卡尔坐标系。早期有研究认为,后顶叶参与视觉坐标和运动坐标之间的转换,但并未明确运动坐标采取的是什么类型的坐标系[14]。
3 可导航空间的梯度表征
相比于近尺度空间的简单结构和可操作性,大尺度可导航空间的复杂性大大增强,这种复杂性体现在如下方面。a. 尺度增大伴随着度量信息的量级增加,需要消耗更多的认知资源选择和存储这些信息。b. 大尺度空间内的导航需要整合不同局部区域的空间信息,实现从单视点空间到环境空间的认知转换,进而形成对空间的整体认知,在此过程中会需要参考框架的转换[15],具体包括自我中心(egocentric)和环境中心(allocentric)两种参考框架,二者分别以自身和外界物体作为空间表征的参考。c. 导航过程需要处理多种可能的意外情况(如原有的路径阻断),实现灵活的绕路和捷径选择[16]。
鉴于上述复杂性,有必要区分性地表征大尺度空间,由于空间尺度的变化是从小到大的连续模式,所以相应的神经表征也应当呈现出这种梯度性。已有研究表明,海马和皮层对大尺度可导航空间的表征确实存在明显的梯度。
3.1 海马长轴梯度
海马和近邻的内嗅皮层参与空间表征的实验证据是多种类型的空间细胞,如海马中编码特定空间位置的位置细胞[17]、内嗅皮层中为空间位置提供背景标度的网格细胞以及编码朝向的头向细胞等[18-19]。基于这些细胞,人类海马长轴或啮齿类海马背腹轴的空间尺度表征呈现出表征粒度上“前粗后细”的梯度变化。
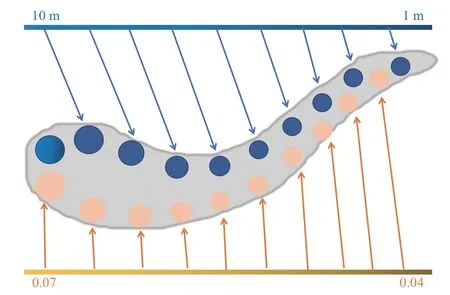
Fig. 2 Spatial gradient along the hippocampal longitudinal axis图2 海马长轴的空间梯度
对环境尺度的差异化表征主要依赖位置细胞发放域在海马长轴上的分离。人类海马前部(对应于啮齿类海马的腹侧)的位置细胞表现出更大的位置域,所能表征的空间信息更为粗糙;相反,海马后部(或背侧)位置细胞的位置域相对较小[20-21](图2)。为了确定空间尺度与海马锥体细胞活动的关联,Kjelstrup等[22]让大鼠在18 m长的线性轨道上来回奔跑,同时记录纵向水平上海马细胞活动,发现CA3 细胞的位置域沿着海马长轴有明确的偏好性分布,从背侧CA3细胞的1 m位置域到腹侧细胞的10 m 位置域,呈现对空间尺度的连续梯度表征。与此同时,海马结构近邻的内嗅网格细胞也沿着背腹轴表现出梯度性[23],从最背侧的50 cm 网格域到最腹侧的3 m网格域,网格细胞表征的空间尺度沿着内嗅皮层的背腹轴逐渐变大,这种空间标度规模的增大与海马表征的逐步粗糙化相照应[24]。
海马长轴上空间尺度的梯度性表征对应着功能的分化。Nadel 等[25]的一项fMRⅠ研究发现,当被试思索详细的空间信息时,海马后部会出现更强的激活,而当思索粗糙的背景信息时,海马前部则会被优先激活,由此表明海马后部对于精确的空间表征和空间行为至关重要,而海马前部则参与编码事件发生的背景。近期颅内电生理研究发现,人类右侧海马theta 频带的能量会受到当前位置和导航目标间距离的调节[26],随着目标接近度提升,theta能量会显著下降,并且这种效应沿着海马长轴从前往后增强,在海马后部最强,由此表明海马存在对目标距离的多尺度空间表征。此外,空间梯度性也能从时间上反映出来,Brunec等[21]研究发现,相比于后部,海马前部具有更强的时间上的自相关性,表明这个区域随着时间的推移信号变化更慢,因此空间信息的更新也更稀疏。
然而,排列在海马长轴上的位置细胞以及分布于内嗅背腹侧的网格细胞二者对空间尺度的梯度表征可能无法直接迁移到大尺度环境,因为两种细胞在大尺度环境下的发放模式可能迥异于在实验室环境下的情况。近期Eliav等[27]对大尺度环境中飞行的野生蝙蝠进行海马CA1 位置细胞活动的无线记录,这项研究颠覆了人们以往对位置细胞的经典认知,发现单个位置细胞在200 m的大尺度环境中表现出多重位置域和多尺度表征,并且这些不同位置域之间的尺寸差异甚至可达到20 倍以上,表明大尺度环境下位置细胞具有极强的位置域调节张力和潜能,这种多尺度编码能够以更高的精确度表征大尺度环境,而且并不需要后天的暴露和学习经验,是先天具备的。在同一实验内,研究者还设计了6 m的小尺度环境,发现在此环境中飞行的蝙蝠其背侧CA1位置细胞只有1~2个位置域,并且最大和最小位置域之间的比值相对更小,表明小尺度环境下位置细胞的位置域调节能力更弱或更刻板僵化,几乎无法实现多尺度编码。
除了研究生态效度更高的大尺度环境下海马CA1位置细胞的特征以外,Ginosar等[28]还创新地对以往研究鲜少涉足的3D空间展开探索,发现3D网格细胞也具有多重发放域,同样改变了人们对网格细胞的经典认知。研究者记录埃及果蝠在3D 房间中飞行觅食过程中内侧内嗅皮质(medial entorhinal cortex,MEC)的活动,发现网格细胞的多重网格域并非基于2D正六边形形成的正六棱柱,而是大致呈球形,并且不同网格域间有特征性距离加以间隔,表现出局部有序性,而非2D 网格细胞表现出的全局有序性。
学生思考一段时间后,给出回答:在研究二次函数性质的基础上,想到可以研究三次函数的奇偶性,比如是奇函数;对称性,包括对称轴、对称中心等;单调性;函数的范围,即函数的定义域、值域等;函数的零点;函数的周期性;函数是否有极值点,如果有,有几个……
大尺度环境下的海马位置细胞和3D 环境下的内嗅网格细胞分别表现出多重发放域,这表明海马长轴和内嗅背腹轴多细胞共同实现的对环境尺度的梯度表征,在面临大尺度和高维环境时可能是无能为力的,但却在单细胞层面实现了多尺度的表征。
3.2 皮层梯度
大脑的感觉运动区域和跨模态区域之间存在一条以皮层为核心的组织轴,反映了从直接的知觉和动作加工到信息的抽象和整合之间连续变化的功能梯度[29]。对于空间表征这一特定领域而言,皮层也表现出了这种功能梯度。
单视点空间或视觉场景(scene view)的加工主要在场景选择性区域(scene-selective regions),此区域会在人们观看场景时发生强烈反应,一般包括3 个功能子分区,分别是旁海马位置区(parahippocampal place area,PPA)、枕叶位置区(occipital place area, OPA) 和 压 后 皮 层(retrosplenial complex,RSC)[30-32]。在解剖上,PPA位于颞枕皮层内侧下部,在海马旁回后部和舌前回之间,OPA位于顶枕皮层,围绕枕横沟,RSC位于内侧顶叶皮层[33]。
OPA 主要负责加工局部场景的空间布局信息[34]、环境的功能性约束边界(如透明障碍物)以及到边界的距离等[35-36];PPA则参与加工场景的诸多特征,如视觉特征、几何特征及凸显的地标等[37]。这两个区域足以表征每一个离散的单视点空间,但是正如Gibson[38]指出,在不同位置间转移涉及不同单视点空间的渐次开闭。随着对环境的探索把所有单视点空间依序加以组织后,就会对由障碍物分隔开的环境空间形成全景表征,这个过程需要RSC参与“缝合”这些独立的单视点空间。
RSC 负责整合来自OPA 和PPA 的局部表征,将局部场景与全局空间进行关联[39],神经电生理记录显示,当在包含多个尺度上的循环轨道上穿梭时,大鼠的压后皮层神经元会表现出周期性的激活模式,表明压后皮层可能参与提取大尺度空间中的子空间并编码这些子空间之间的关系,进而辅助路径整合[40]。fMRⅠ研究发现,相比于倒序呈现,路径上位置点的依序出现会引发RSC 更强烈的活动[41],表明RSC 参与编码路线的拓扑结构。也有研究者提出不同观点,认为场景选择区域中有分别表征场景分类和导航的两个系统,其中PPA参与场景分类[42],而OPA 和RSC 参与导航过程,并且OPA 支持的是以视觉信息为主导的导航(visually guided navigation),RSC支持的是基于地图的导航(map-based navigation)[33],即OPA 采取自我中心表征,而RSC 则采取环境中心表征,RSC 在将视点依赖的空间表征转化成独立于观察者本身的“地图式”表征中发挥关键作用[43]。
内侧颞叶、内侧顶叶和外侧顶枕皮层这3个脑区从后部到前部都表现出对空间尺度变化的梯度表征,并且三者的后部与场景选择区域均有部分重叠,由此开始向前逐步扩大表征尺度[44]。Peer等[43]让被试判断6 种空间尺度内(分别是房间、建筑、社区、城市、国家、大洲)物体间的距离,同时进行fMRⅠ扫描,发现内侧颞叶偏好建筑、社区这类中小尺度,内侧顶叶偏好城市、国家这类大中尺度,而外侧顶枕皮层则偏好房间和大洲这两个可导航空间中最小和最大的尺度。这3个皮层区域都接近场景选择区域,进一步分析显示,三者的后部都起始于场景选择区域,从这里的局部单视点空间表征逐步向前扩大表征尺度,并且随着尺度的增大空间表征也逐渐从具体转变为抽象和符号化,这从被试的口头报告也能形象地反映出来。
近期Dong 等[45]的核磁研究发现,海马旁回偏好对中等尺度(建筑)的表征,前扣带回则倾向于对大中尺度空间(建筑和街区)有更强的激活,顶回和颞回在大尺度空间(城市、国家和大洲)中有更强的激活,与上述研究结果基本一致。不过除了上述尺度依赖性的脑区以外,此研究同时也发现枕中回会同时参与多个尺度的空间表征,表现出无标度性(scale-free)。
4 抽象社会关系空间的二分表征
已有研究表明,社会关系尺度的远近在皮层上存在二分化的表征。Roseman 等[47]将社交尺度(或情感亲密程度)分成最亲密的人、密友、朋友、有联系的人和不联系的熟人五类,尺度越大代表情感亲密度越低,让被试分别对不同社交尺度内的成员进行亲密度的判断,同时进行fMRⅠ扫描,结果发现,颞顶叶联合区、颞中回、楔前叶以及背内侧前额叶偏好表征最亲密的情感关系,而内侧颞叶、压后皮层以及腹内侧前额叶对其他所有的社交尺度表现出偏好。社交尺度的这种二分表征将社会关系中少数最亲密的人与其他人区分开来,突出这些人在个体社会生活中更可能为其提供支持和帮助,发挥更为关键的作用。
大脑对物理导航空间尺度的表征模式可能也会迁移到社会认知领域的尺度表征上,具体表现为相似或重叠的脑网络[48]。Roseman等[47]的研究还发现,社交尺度表征与物理上可导航空间尺度表征之间的相似性,即情感上亲密的社会关系表征(如最亲密的人、密友)与物理上小尺度的空间表征(如房间、建筑)在激活的脑反应上表现出显著的相关性,而情感上疏远的社会关系表征(如不联系的熟人)则与物理上大尺度的空间表征(如街区、城市)在激活的脑反应上表现出显著的相关性。
5 研究展望
5.1 多尺度空间表征的时间进程和研究范式
目前对多尺度空间表征的研究大部分采取的都是空间分辨率更好的fMRⅠ技术,从而考察不同尺度空间表征所在的脑区差异,并没有关注在特定尺度空间下偏好该尺度的相应脑区活动的时间进程、有关尺度大小的信息何时引起人的注意、不同空间尺度的这一时程特征会否有差别等问题。
虽然从房间、街区到国家大洲等多个尺度的空间所对应参与表征的脑区目前已有众多发现,但是并不知道人脑在观看到一张空间图片时,是先提取出该空间的尺度信息,然后输送到负责该尺度的脑区进行加工,还是先进行无尺度差别的加工,然后再交由各个专一于特定尺度的脑区进行“润色”。如果是首先识别出空间的尺度信息,那么就需要未来的研究寻找识别尺度信息的脑区及其参与识别的时间动态性。
此外,导航和空间表征的实验室研究一般都采取较小的空间尺度,尤其以单视点空间最为常见,比如水迷宫、旷场,这与人类和某些动物(如蝙蝠)的真实导航过程差异很大,人类导航行为最常发生的空间是环境空间而非简单的单视点空间,而蝙蝠的导航则发生在几百公里的大尺度空间中[49]。由于实验室模拟真实导航环境多重尺度存在难度,目前有关空间尺度的研究往往是呈现相应尺度的空间图片,让被试进行该尺度下的距离判断[44-45],被试的真实导航参与感并不强,未来研究可以考虑采取虚拟现实技术变化空间尺度,提高研究的生态效度。Ulanovsky课题组[27-28]对蝙蝠的单细胞记录研究刷新了人们对位置细胞和网格细胞的认知,也提示了高生态效度的必要性,然而目前对于能够在更大尺度(如国际、洲际甚至宇宙层面)内进行导航规划的人类而言,还没有更多的研究去探索其背后的神经机制,这也是未来需要关注的方向。
5.2 时间尺度表征的梯度性
空间和时间信息是构成情景记忆的两个重要部分,可导航空间尺度在海马、皮层区域表征所表现出的梯度性可能也同样适用于时间维度。Monsa等[50]的fMRⅠ实验发现,顶叶的活动具有时间尺度的敏感性,展现出从外侧向内侧的梯度表征,时间尺度从小到大,即外侧顶叶对小尺度的时间范围(如小时、天)选择性更高,而内侧顶叶则偏好大尺度的时间范围(如月、年),但是在海马长轴却没有发现对时间尺度表征的梯度性。然而,近期电生理研究发现,沿海马长轴从前往后(即从海马头到海马尾),theta 振荡的尖峰(peak)频率逐渐增大,表明海马长轴对时间尺度存在梯度表征,越靠近后部的海马区域所表征的时间尺度越小[26]。海马长轴的时间尺度表征可能是由空间尺度的表征迁移而来,还需要未来研究进一步探索。
由此可见,大脑对时间尺度和空间尺度的表征存在重叠的区域,也存在特异的区域。然而,对于特定的情景记忆而言,时间尺度和空间尺度的信息需要整合起来,未来研究需要关注时间尺度表征的梯度性和空间尺度表征的梯度性如何整合或交互。
5.3 不同空间尺度下的认知地图
可导航空间的表征能够为导航行为提供帮助,在功能上类似于地理学意义上制作的地图,因此也被称为认知地图(cognitive map)[51]。目前对于认知地图的存在形式存在多种理论争议,欧式假设认为,认知地图采取绝对统一的欧式度量结构,保留了物理空间中已知位置间包括距离、角度等在内的几何特征。Widdowson和Wang[52]发现,无论空间曲率如何(欧式空间、双曲空间或球面空间),被试进行指向任务时总会与欧式方向匹配,据此表明路径整合和空间更新系统是基于欧式几何学运作的。然而也有研究者通过“虫洞”实验证明欧式假设的局限性[53]。
另一方面,拓扑图假设认为,认知地图仅仅编码粗糙的拓扑图结构,包含节点网络和节点间的连接边,一般而言节点表示位置,而边代表位置间的路径关系。这种“位置图”(place graph)捕获了位置间的连接性,但不需要将位置嵌入到全局一致的坐标系统中(coordinate free)。由于单纯的拓扑图结构不包含任何度量性信息,Warren[54]提出有标签图假设,在图结构的基础上辅以边的权重和边之间的夹角。
对认知地图存在形式的这些理论争议本质上可能是空间尺度的影响,不同研究中所用的实验范式差异导致导航发生的背景和空间表征的对象存在较大差异,比如有的研究采取单视点空间,有的研究则采取复杂的环境空间或更大尺度的地理空间,不同空间尺度下的认知地图很可能具有不同的形式。直观上,大尺度空间由于认知负载大而需要简化,更可能采取拓扑图结构表征,而近尺度空间和单视点空间对精确性要求更高,相应地需要采取欧式表征。未来研究需要设计相应实验范式直接考察空间尺度对认知地图存在形式的影响及其神经机制。
猜你喜欢
矿产勘查(2020年8期)2020-12-25
中国现代医药杂志(2020年3期)2020-05-08
中国生物医学工程学报(2019年6期)2019-07-16
魅力中国(2016年42期)2017-07-05
中外医疗(2016年15期)2016-12-01
河南电力(2016年5期)2016-02-06
软件导刊(2015年8期)2015-09-18
新闻前哨(2015年2期)2015-03-11
中国水利(2015年5期)2015-02-28
湖南大学学报·自然科学版(2014年10期)2014-11-20
