
盐胁迫条件下竹柳光合作用日变化特征研究
2024-03-20 05:28:16赵育鹏
安徽农业科学 2024年4期
摘要 以竹柳盆栽苗木为试验材料,分析其在不同盐分浓度下的净光合速率、气孔导度、蒸腾速率、胞间CO2浓度等光合生理指标的日变化特征。结果表明:竹柳在对照与轻度胁迫条件下,净光合速率的日变化特征表现为双峰型;在中度和重度胁迫条件下,为单峰型曲线,变化趋势表现为先上升后下降。气孔导度在对照与轻度胁迫条件下的日变化特征表现为双峰型;在中度和重度胁迫条件下为单峰型曲线,呈逐渐下降的趋势。蒸腾速率的日变化特征与气孔导度类似。各胁迫处理胞间CO2浓度均较对照高,且随胁迫程度的加剧,胞间CO2浓度也随之上升。
关键词 竹柳;盐胁迫;净光合速率;气孔导度;蒸腾速率;胞间CO2浓度;日变化
中图分类号 S718 文献标识码 A
文章编号 0517-6611(2024)04-0087-03
doi:10.3969/j.issn.0517-6611.2024.04.018
开放科学(资源服务)标识码(OSID):
Study on Photosynthetic Diurnal Variation of Salix matsudana Under Salt Stress
ZHAO Yu.peng
(Shanxi Academy of Forestry and Grassland Sciences, Taiyuan, Shanxi 030012)
Abstract The Salix matsudana were used as experimental material to study the effect of salt stree on the photosynthetic diurnal variation.The results showed that the photosynthetic rate of Salix matsudana showed a bimodal type in the standard contrast and mild stress conditions,while showed a single peak type in the moderate and severe stress condition with a trend first rise then falling.The stomatal conductance showed a bimodal type in the standard contrast and mild stress conditions,while showed a single peak type in the moderate and severe stress conditions with a trend of gradual decline.The daily variation trends of transpiration rate was similar with the stomatal conductance.The intercellular CO2 concentration under stress treatment were higher than the standard contrast,and increased with the salt stress levels.
Key words Salix matsudana;Salt stress;Photosynthetic rate;Stomatal conductance;Transpiration rate;Intercellular CO2 concentration;Diurnal variation
作者简介 赵育鹏(1988—),男,山西陵川人,工程师,从事森林生态研究。
收稿日期 2023-03-13;修回日期 2023-04-14
盐碱土是地球陆地上分布广泛的一种土壤类型,约占陆地面积的25%[1]。土壤的盐渍化是一个世界性的资源与生态问题[2],是制约农林业生产的主要因素之一[3],同时对生态环境造成巨大威胁[4-6]。一般植物在盐碱条件下生长不良甚至难以成活,耐盐碱植物的引种选育和利用研究一直是人们关注的热点,其在盐碱地造林绿化中发挥生态效益的同时,还能降低土壤含盐量,起到改良土壤的作用[7-8]。
竹柳是杨柳科(Salicaceae)柳属(Salix)乔木,其枝干笔直如柳,叶片狭长如竹,具有适应性强、生长快、耐盐碱、耐寒、耐旱等特点[9-10],是盐碱地造林、湖泊滩涂造林、园林绿化、环境生物修复的理想树种,具有极大的推广价值和广阔的发展前景[11-16]。笔者通过竹柳在不同盐分浓度胁迫下的光合生理指标来分析其光合作用变化特征,旨在为竹柳在盐碱地区的开发利用提供基础依据。
1 材料与方法
1.1 试验材料
选取1年生竹柳实生苗为供试材料,于4月下旬至5月上旬定植于塑料花盆中,采用腐殖质土、珍珠岩、草炭体积比为1∶1∶3的混合基质,基质用多菌灵消毒处理,定植后置于温室大棚中。于7月底,选择长势相对一致的苗木进行试验。
1.2 试验设计
采用完全随机区组设計,设置盐分胁迫浓度为0(标准对照)、1‰(轻度胁迫)、4‰(中度胁迫)、7‰(重度胁迫),浓度用质量分数表示为NaCl占土壤重的比例,每个浓度处理10株苗木,3次重复。盐胁迫试验期间,采取多次施盐、逐级递增的施盐方式,同时对苗木进行定期定量浇水、防治病虫害等常规养护管理。为防止浇水时盐分流失,在花盆下垫塑料托盘,将浇水时流出的溶液及时倒回花盆内[17]。
1.3 光合日变化的测定
仪器采用Li-6400便携式光合作用测量仪,于8月份选择1~2个晴朗天气,06:00—18:00每隔2 h测定1次,选取自生长点向下第3~4片完全展开叶进行测定。读取叶片瞬时净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等值。每个处理3次重复进行统计分析[18]。试验结束后,用软件导出数据,用Excel做统计分析及绘图。
2 结果与分析
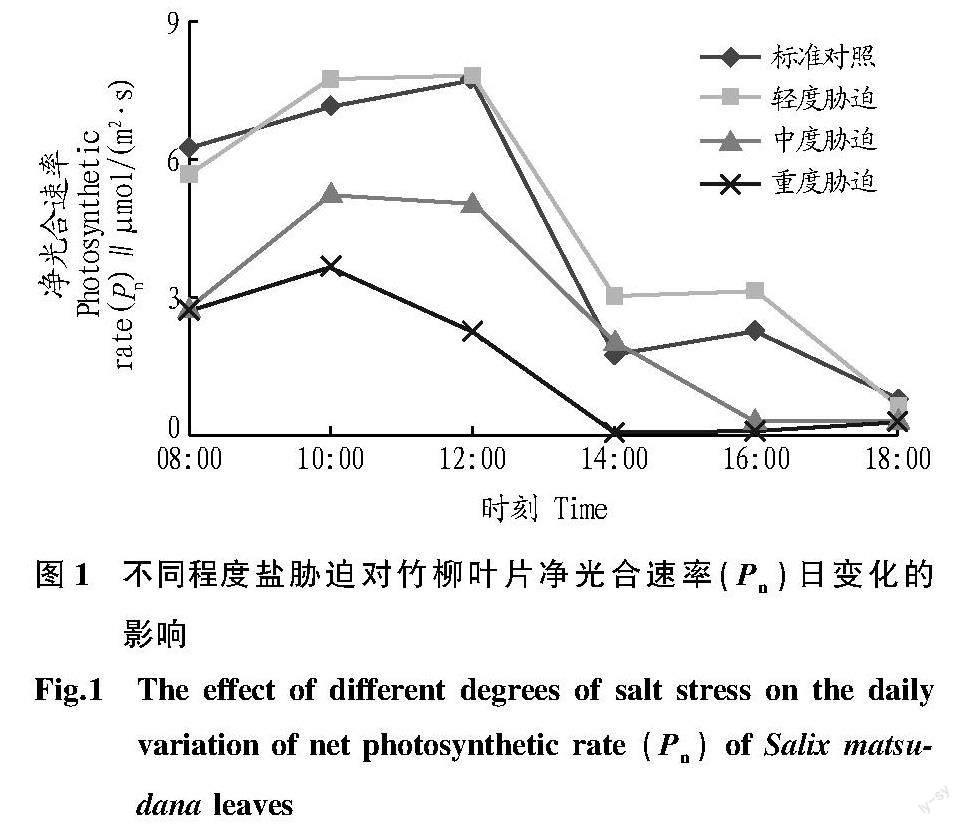
2.1 净光合速率(Pn)的日变化
由图1可知,竹柳在对照与轻度胁迫条件下,净光合速率日变化总体呈双峰型趋势,但变化趋势不尽相同;中度与重度胁迫均为单峰型曲线,变化趋势表现为先上升后下降。对照与轻度胁迫处理的净光合速率峰值出现在中午时段,此后迅速下降,在16:00左右出现另一个峰值;中度与重度胁迫处理的峰值都出现在10:00,此后逐渐下降。在所有胁迫条件下,最大值出现在轻度胁迫条件下的12:00,最大值为7.855 4 μmol/(m2·s),最小值出现在重度胁迫处理。标准对照、轻度胁迫、中度胁迫、重度胁迫处理全天平均光合速率分别为4.310 7、4.683 5、2.632 1、1.517 0 μmol/(m2·s),轻度胁迫平均光合速率最高,重度胁迫最低。
对照与各处理峰值之间比较,中度胁迫、重度胁迫与对照的峰值不同,轻度胁迫变化趋势与对照基本一致;在08:00之前,各处理净光合速率均低于对照,此后,轻度胁迫略高于对照外,并维持较长时间净光合速率高峰,其他处理均低于对照,并随胁迫程度增加而急剧下降;重度胁迫出现明显的“午休”现象,所有处理在午后净光合速率都明显下降,对照与轻度胁迫在下午有所回升,都在18:00均降到很低水平(图1)。
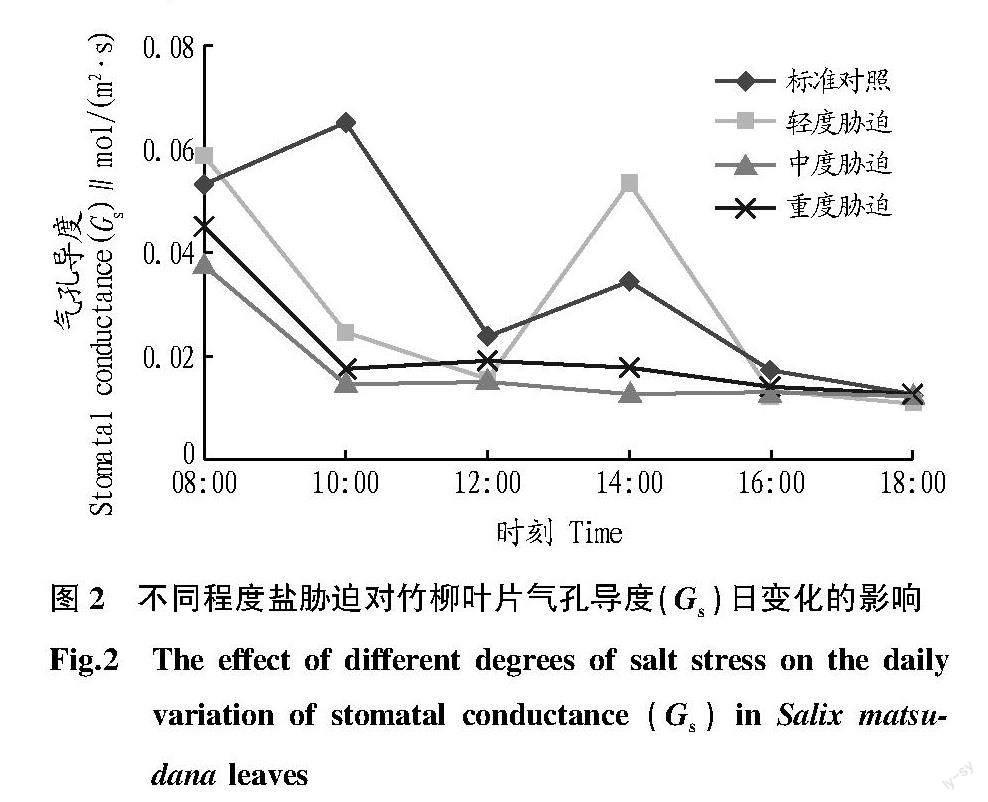
2.2 气孔导度(Gs)的日变化
由图2可知,竹柳在对照条件下,气孔导度的日变化呈双峰型;中度与重度胁迫处理为单峰型曲线,呈逐渐下降趋势。对照的气孔导度峰值分别出现在10:00与14:00;轻度胁迫处理在14:00出现一个峰值并处于较高水平,其余处理除了08:00外均处于较低水平,在12:00左右都发生“午休”现象。对照、轻度胁迫、中度胁迫、重度胁迫处理全天平均气孔导度分别为0.034 6、0.029 3、0.017 7、0.021 2 mol/(m2·s),对照的平均气孔导度值最高,中度胁迫与重度胁迫处理较低。各处理在16:00以后处于较低水平。
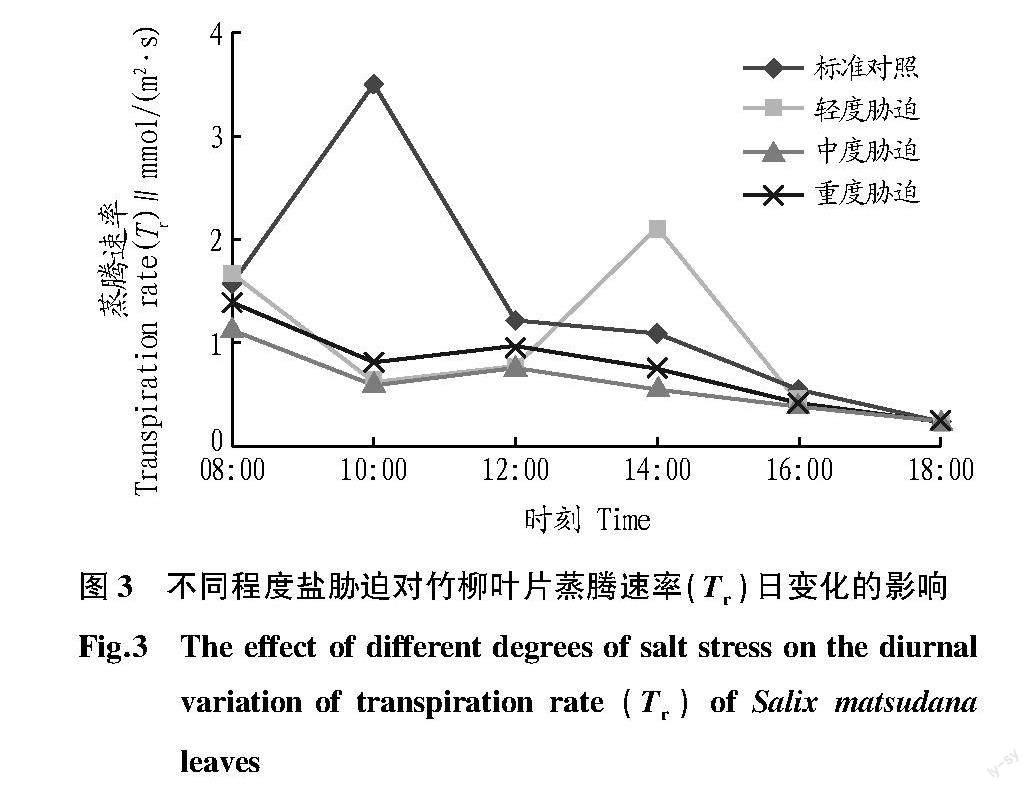
2.3 蒸腾速率(Tr)的日变化
从图3可以看出,竹柳蒸腾速率的日变化趋势与气孔导度的日变化趋势相似,对照与轻度胁迫处理的峰值分别出现10:00与14:00,分别为3.499 1、2.095 5 mmol/(m2·s),其余处理均处于较低水平。对照、轻度胁迫、中度胁迫、重度胁迫处理全天平均蒸腾速率分别为1.357 6、0.965 0、0.618 8、0.764 8 mmol/(m2·s),这与气孔导度的日平均值变化趋势一致,对照平均蒸腾速率最高,中度胁迫与重度胁迫处理较低。各处理在14:00以后均迅速下降。
2.4 胞间CO2浓度(Ci)的日变化
由图4可知,竹柳胞间CO2浓度除对照外,其他处理在10:00维持在较高水平,轻度胁迫和重度胁迫处理在14:00最低,之后又逐渐上升;而对照在16:00后迅速上升,且低于其他处理;重度胁迫处理在12:00之前都高于其他处理,12:00以后与中度胁迫处理的胞间CO2浓度高于轻度胁迫与对照。对照、轻度胁迫、中度胁迫、重度胁迫处理全天平均胞间CO2浓度分别为281.926 5、337.742 7、354.084 6、372.821 0 μmol/mol。总体来说,盐胁迫处理胞间CO2浓度比对照高,各处理全天胞间CO2浓度总体都呈上升趋势,且随胁迫程度的加剧,胞间CO2浓度也随之上升。
3 结论与讨论
该研究通过对竹柳各项光合指标日变化的测定得出:竹柳净光合速率日变化趋势在正常生长环境条件下呈双峰型曲线,净光合速率日变化对不同程度盐胁迫的响应有明显差异。中度胁迫与重度胁迫处理较对照下降快,各处理在10:00—12:00均达峰值,轻度胁迫全天净光合速率平均值比对照高8.65%,说明轻度盐胁迫能刺激提高竹柳的光合速率,且随着盐胁迫程度的增加,净光合速率达峰值的时间逐渐提前,说明竹柳属于喜光植物,适度的增加光照强度能促进其光合作用,但同时也下降较快。这可能是由于竹柳受到盐分浓度的影响,叶片的光合作用场所叶绿体受到影响,改变了其正常的光合作用規律。
竹柳的气孔导度变化趋势与蒸腾速率相似,对照与轻度胁迫时为单峰型曲线,但峰值出现时间不一致;中度胁迫与重度胁迫条件下表现为逐渐下降的曲线,这是由于早晨气温较低、空气相对湿度较高,各处理的气孔导度随时间逐渐下降,且在中午出现“午休”现象。各盐胁迫处理的胞间CO2浓度高于对照,说明竹柳在盐胁迫条件下,刺激气孔光合CO2同化能力,而限制非气孔光合CO2同化能力。如果气孔导度对盐胁迫不敏感,在盐胁迫条件下仍会维持较高水平的气孔导度,胞间CO2浓度不会成为限制因素,这时光合速率仍下降,则是由非气孔因素受限所致[19]。通常认为,中午由于叶片温度的上升,使叶片-空气蒸汽差值增加,引起气孔导度下降,是导致植物光合作用“午休”的主要原因[20]。
通过对竹柳光合生理指标在不同盐分浓度胁迫下的日变化趋势分析发现,对照和轻度胁迫处理下的光合速率、气孔导度及蒸腾速率值均比中度和重度胁迫高,说明轻度盐胁迫对竹柳的光合作用有一定促进作用,在轻度盐胁迫条件下竹柳可以维持正常的光合生理特性,能够正常生长,其具有一定的耐盐性,但是随着盐胁迫程度的不断加剧,竹柳的生长发育明显受到抑制。综上,竹柳可作为轻度盐碱地的栽植树种,同时,在盐碱地造林中,可按照林地盐分含量对其进行分类,科学选择造林树种,达到适地适树,最有效地发挥盐碱地的生产潜力,充分利用盐碱地资源,实现盐碱地的可持续发展[21]。
参考文献
[1]张川红,沈应柏,尹伟伦,等.盐胁迫对几种苗木生长及光合作用的影响[J].林业科学,2002,38(2):27-31.
[2]MUNNS R.Comparative physiology of salt and water stress[J].Plant,cell and environment,2022,25(2):239-250.
[3]JUN S S,CHOI H J,LEE H Y,et al.Differential protection of photosynthetic capacity in trehalose.and LEA protein.producing transgenic plants under abiotic stresses[J].Jouranal of plant biology,2008,51(5):327-336.
[4]江钰娜,方威,吴瑜玮,等.盐胁迫对15种乔木柳插条表型和生理指标影响[J].西北林学院学报,2021,36(1):117-125.
[5]DEINLEIN U,STEPHAN A B,HORIE T,et al.Plant salt.tolerance mechanisms[J].Trends in plant science,2014,19(6):371-379.
[6]VAN ZELM E,ZHANG Y X,TESTERINK C. Salt tolerance mechanisms of plants[J].Annual review of plant biology,2020,71:403-433.
[7]白善军,李玉娟,张健,等.沿海滩涂竹柳直插造林技术[J].现代农业科技,2010(13):241.
[8]王文成,郭艳超,李克晔,等.盐胁迫对竹柳种苗形态及生理指标的影响[J].华北农学报,2011,26(S1):143-146.
[9]张健,李玉娟,张树清,等.美国竹柳生物特性与繁殖试验简报[J].上海农业科技,2009(6):116,118.
[10]崔立勇,佟庆,梁成斌.竹柳造林地选择、造林时间和方法[J].现代园艺,2014(10):59-60.
[11]唐胶,彭祚登,贾清棋,等.城市排水污泥对竹柳扦插苗生理特性及N、P、K含量的影响[J].福建农林大学学报(自然科学版),2021,50(5):611-618.
[12]麻浩,程志义,钱芳,等.竹柳种植新技术及其在荒漠和盐碱地中的应用[J].江苏农业科学,2020,48(12):119-122.
[13]孙佳,夏江宝,苏丽,等.黄河三角洲盐碱地不同植被模式的土壤改良效应[J].应用生态学报,2020,31(4):1323-1332.
[14]刘向鸿,席忠诚,曹思明.氮、磷、钾肥不同配比施肥对竹柳生长的影响[J].林业科技通讯,2016(10):22-24.
[15]徐克顺.竹柳生物学特性及扦插育苗技术[J].安徽农学通报,2011,17(20):121-122.
[16]邵志广.NaCl处理对竹柳苗光合特性的影响[J].安徽农业科学,2014,42(19):6139-6141.
[17]杨升.滨海耐盐树种筛选及评价标准研究[D].北京:中国林业科学研究院,2010.
[18]吾木提汗·卡克木,海利力·库尔班,陈其军,等.盐胁迫条件下骆驼刺与绿豆光合日变化特征及午休现象的成因[J].干旱区研究,2012,29(6):1039-1045.
[19]CHAVES M M,FLEXAS J,PINHEIRO C.Photosynthesis under drought and salt stress:Regulation mechanisms from whole plant to cell[J].Annals of botany,2009,103(4):551-560.
[20]PONS T L,WELSCHEN R A M.Midday depression of net photosynthesis in the tropical rainforest tree Eperua grandiflora:Contributions of stomatal and internal conductances,respiration and Rubisco functioning[J].Tree physiology,2003,23(14):937-947.
[21]劉平,潘文利,魏忠平,等.盐胁迫下白蜡和小胡杨光合生理日变化的响应研究[J].北方园艺,2012(1):75-78.
猜你喜欢
节水灌溉(2021年5期)2021-05-26 06:49:38
中国农业科技导报(2020年12期)2020-03-15 10:50:08
江苏农业科学(2016年8期)2017-02-15 19:41:06
山东农业科学(2016年12期)2017-01-21 14:20:16
中国科技博览(2016年1期)2016-04-25 07:47:28
浙江林业(2015年5期)2015-12-12 03:39:39
吉林林业科技(2015年6期)2015-09-17 01:56:20
农民致富之友(2014年6期)2014-04-29 10:07:00
现代营销·经营版(2013年5期)2013-05-14 14:55:04
赤峰学院学报·自然科学版(2012年19期)2012-10-14 02:53:58
