
3种佐剂对猪瘟病毒E2蛋白免疫效果的影响
2024-03-18 07:09:42陈玉梅刘运超祁艳华周景明王爱萍
动物医学进展 2024年3期
陈玉梅,刘运超,祁艳华,梁 超,周景明,王爱萍*
(1.郑州大学生命科学学院,河南郑州 450001;2.河南省农业科学院动物免疫学重点实验室,河南郑州 450001)
猪瘟(Classical swine fever,CSF)是由猪瘟病毒(Classical swine fever virus,CSFV)引起的一种急性传染病[1-2]。近年控制猪瘟应用最广泛的是减毒活疫苗[3],经典的CSFV疫苗,如C-strain、GPE-strain、LPC-strain和LK-VNIVViM strain,虽然不具有区分感染和接种疫苗动物的能力,但提供了对CSFV感染的强大保护。FlagT4Gv、TWEJ2、FLC-LOM-Berns等标记疫苗接种家猪后可控制CSF的传播并降低急性死亡率,但疫苗的广泛使用也导致了猪瘟的复杂性,如临床症状从典型转向非典型,免疫抑制和持续感染的现象普遍存在,这也给控制CSF带来了困难[4]。
囊膜糖蛋白E2是CSFV病毒颗粒的主要成分,有多种功能,如病毒对细胞的吸附、保护性免疫反应的激发和病毒毒力[5]。研究显示,强毒CSFV的Erns(E0)、E1或E2糖蛋白的糖基化降低了对猪的病毒毒性[1]。而糖蛋白E2和E0是感染动物中激发抗猪瘟抗体的主要靶点,因此基于E2和E0靶点的疫苗研究很多,用杆状病毒系统表达糖蛋白E2和E0,并测试了它们在兔体内的保护性免疫,发现E2蛋白诱导的特异性抗体、中和抗体和细胞免疫应答水平较高,而仅接种E0不能诱导保护性免疫反应[6]。通过腺病毒载体表达E0和E2基因,构建了重组腺病毒E0-E2(rAd-E0-E2)疫苗,并确定了最小免疫剂量和免疫持续时间[7]。制备E2-CD154亚单位候选疫苗,其在怀孕母猪中是安全有效的,并且在出生后第63天,传递给后代的被动免疫仍然具有保护性[8]。用于CSFV E2纯化的肽配体进行分子对接,从转基因水稻种子中表达纯化E2蛋白,对小鼠免疫抗体的评估表明,接种后第42天抗体阻断率达到86.18%和90.68%[9],而通常情况下阻断率Blocking%≥50%的样本可认定为产生保护。
体外表达的CSFV E2蛋白可以刺激机体的保护性免疫应答,产生高滴度的特异性抗体和中和抗体,有效抑制病毒感染[10]。但由于CSFV E2蛋白亚单位疫苗只有一种免疫原,与全病毒相比,其T细胞免疫应答水平较弱,产生的保护性抗体滞后,限制了其推广应用。佐剂是一种非特异性免疫增强剂,其本身不能让机体产生免疫应答,但其用在疫苗中便能激发强烈的体液、细胞免疫,可以诱发机体针对特定抗原产生更高效、更长期的免疫应答。兽用疫苗常用的佐剂有无机佐剂、有机佐剂、合成佐剂以及油佐剂等[11]。其中氢氧化铝(明胶)为常用的无机佐剂,明胶吸附的抗原呈颗粒状,可增强抗原呈递细胞(Antigen-presenting cells,APC)对抗原的摄取,氢氧化铝佐剂主要诱导血清抗体反应;50V佐剂剂型为油包水(W/O)乳化液,主要是植物来源的甘露醇和油酸高度精炼制成,在猪体内W/O乳剂主要功能是促进抗体产生、淋巴细胞增殖和临床保护;MontanideTM ISA 201 VG(201佐剂)为双相佐剂,水包油包水(W/O/W)剂型[12],主要成分为矿物油,抗原被包裹在内部水滴和外部水相中,外水相中的抗原容易被免疫系统利用,而内部水滴中的抗原则缓慢释放。
本文研究铝佐剂、50V佐剂和201佐剂分别与CSFV E2蛋白联合制备亚单位疫苗免疫Balb/c小鼠,评价不同佐剂对CSFV E2蛋白刺激机体T/B细胞免疫反应,筛选出高效的免疫佐剂,为提高CSFV亚单位疫苗的免疫效果,揭示不同佐剂类型对CSFV E2蛋白亚单位疫苗免疫在小鼠体内诱发免疫反应的差异,为CSFV E2蛋白亚单位疫苗的研制提供理论依据。
1 材料与方法
1.1 材料
1.1.1 载体与菌种 pFast Bac I、pFast Bac I- gp67-E2、NL-E2等质粒和菌种均为郑州大学分子免疫学实验室保存。
1.1.2 细胞系和病毒株 Sf 21细胞、PK-15细胞有郑州大学分子免疫学实验室培养并保存;CSFV石门强毒株、CSFV疫苗毒株等均由郑州大学分子免疫学实验室增殖及保存。
1.1.3 实验动物 4~6周龄雌性Balb/c小鼠,购自河南省郑州市实验动物中心。
1.1.4 主要试剂BamHⅠ和XhoⅠ内切酶,NEB公司产品;RPMI 1640培养基、IL-2、IL-4、IL-10、IFN-γ ELISA KIT及Hematoxylin-Eosin/HE Staining Kit,北京索莱宝科技有限公司产品;Sf 900TMⅡ SFM昆虫细胞无血清培养基(10902104)、CellfectinTMⅡ Reagent转染试剂(10362100),赛默飞世尔科技公司产品;佐剂MONTANIDETMISA 50 V2,法国赛比克(Seppic)公司产品;Anti-6×His-tag antibody,Proteintech;HRP-G@M-IgG,Abcam公司产品;DH10Bac感受态细胞(BC112-01)、Trans5α感受态细胞(CD201-01),北京全式金生物技术有限公司产品;胰酶、胎牛血清(FBS),Gibco公司产品;Ni-NTA亲和层析填料,Norvagen公司产品;其他常规试剂均为分析纯。
1.1.5 主要仪器 酶标仪(iMark)、洗板机(1575)和凝胶成像系统 ,美国伯乐公司产品;多功能酶标仪,美国MD公司产品。
1.2 方法
1.2.1 CSFV E2蛋白的纯化及鉴定 以pFast Bac I-gp67-E2重组质粒转染DH10Bac感受态细胞制备粘粒NL-E2,以粘粒NL-E2转染Sf 21细胞分别制备P1代、P2代和P3代感染病毒;以P3代阳性感染病毒为毒种扩大培养,表达CSFV E2重组蛋白,采用Ni2+柱亲和层析咪唑梯度洗脱纯化制备重组E2蛋白,用SDS-PAGE及Western blot进行鉴定。
1.2.2 佐剂配伍及免疫程序 将纯化获得的重组E2蛋白分别与铝佐剂、50V佐剂和201佐剂配伍制备亚单位疫苗,具体配伍方式见表1。分别选取30只健康的Balb/c小鼠随机分成6组,每组5只(表1),背部皮下多点注射;首免后21 d加强免疫1次。免疫后0、14、21、28、35 d断尾采血收集血清。

表1 每剂疫苗抗原含量
1.2.3 间接ELISA测定抗体效价及抗体消长规律 用碳酸盐缓冲液(0.05 mol/L、pH9.6,CBS)将E2蛋白稀释成2 μg/mL,按100 μL/孔加入96孔酶标板内,4℃包被过夜;用磷酸盐吐温缓冲液(Phosphate buffered saline tween-20,PBST)洗板3次,加入5%脱脂奶粉37℃封闭2 h;弃去封闭液,PBST洗板2次,甩干,一抗用待检小鼠血清,按2倍倍比稀释,即1∶100、1∶200……1∶102 400,每孔加入50 μL稀释后血清,同时,设1∶500稀释的小鼠阳性血清为阳性对照,37℃ 反应1 h;二抗使用1∶5 000稀释的HRP-标记的羊抗鼠IgG,37℃ 反应30 min,TMB显色液显色,测定抗体效价。
1.2.4 阻断ELISA检测 采用IDEXX猪瘟抗体检测试剂盒,对小鼠血清进行检测(将小鼠血清稀释20倍用于检测),具体操作参照试剂盒说明书。
1.2.5 淋巴细胞分离增殖和细胞因子测定 免疫后第35天采集抗凝血,处死小鼠分离脾脏,用索莱宝的淋巴细胞分离试剂盒分离淋巴细胞,然后使用CCK-8进行淋巴细胞增殖实验,同时使用索莱宝中的小鼠IL2、IL4、IL10、IFN-γ ELISA KIT对免疫后35 d小鼠血清进行检测,分析IL2、IL4、IL10和IFN-γ的表达情况。
2 结果
2.1 CSFV E2蛋白的纯化与鉴定
以杆状病毒表达系统重组表达CSFV E2蛋白,用亲和层析纯化制备重组E2蛋白,经SDS-PAGE及Western blot鉴定获得的重组E2蛋白纯度大于95%,糖基化的E2蛋白分子质量为48~62 ku,可以与猪瘟病毒E2蛋白单克隆抗体发生特异性反应(图1),为进一步进行免疫效果评价奠定基础。

M.蛋白分子质量标准;1.SDS-PAGE鉴定;2.Western blot鉴定
2.2 特异性抗体效价测定
用间接ELISA检测小鼠抗体应答情况,发现在首免后3周(即免疫后第21天)和二免后2周(即免疫后第35天),A组铝佐剂组抗体效价分别为1∶1 600和1∶51 200,B组50V佐剂组抗体效价分别为1∶12 800和1∶204 800,C组201佐剂组抗体效价分别为1∶6 400和1∶102 400,D组阳性对照组抗体效价分别为1∶6 400和1∶102 400,E组无佐剂组E2蛋白免疫后抗体效价分别为1∶1 600和1∶3 200,F组PBS阴性对照组无特异性抗体产生(图2)。

A.一免后21 d血清的抗体效价;B.一免后35 d血清的抗体效价
2.3 抗体消长规律测定
测定免疫后0、14、21、28、35 d的抗体消长规律,结果显示,在免疫后第14天各疫苗免疫组特异性抗体已经产生,并随时间推移而升高,在第21天(二免以后1周)至28 d抗体水平进一步升高直至第35天,特异性抗体有一个明显的提升,且有佐剂的A、B、C、D组明显高于无佐剂的E组,F组PBS阴性对照组无特异性抗体产生(图3)。
A.抗体消长规律检测;B.免疫后14、21、28、35 d抗体效价测定
2.4 抗体阻断率的检测
测定免疫后14、21、28、35 d的抗体阻断率,结果在免疫后第21天,B、C和D组抗体阻断率大于50%,在免疫后第28天A组抗体阻断率大于50%,而无佐剂的E组抗体阻断率则明显低于A、B、C和D组,F组PBS阴性对照组无阻断抗体产生(图4)。

图4 血清抗体阻断率检测
2.5 不同佐剂对E2蛋白刺激机体T细胞的免疫应答
淋巴细胞经ConA或灭活猪瘟病毒刺激后,用CCK-8试剂盒检测淋巴细胞增殖活性,经ConA或灭活猪瘟病毒刺激,与PBS对照组相比,淋巴细胞活性均无显著差异(图5)。分别用ELISA检测免疫血清中细胞因子IFN-γ、IL-2、IL-4、IL-10等的表达变化,发现IL-2表达量在10.52~28.46 pg/mL,各免疫组与阴性对照组相比没有显著差异(图6);IL-4表达量在498.4~892.23 pg/mL,与F组相比,A、B、C组差异显著,其中B组50V佐剂组表达量最高为892.23 pg/mL,D、 E组差异不显著(图6);IL-10表达量在31.20~59.46 pg/mL,各免疫组与阴性对照组相比差异均不显著(图5);IFN-γ表达量在44.13~56.47 pg/mL,各免疫组与阴性对照组相比均没有显著差异(图6)。
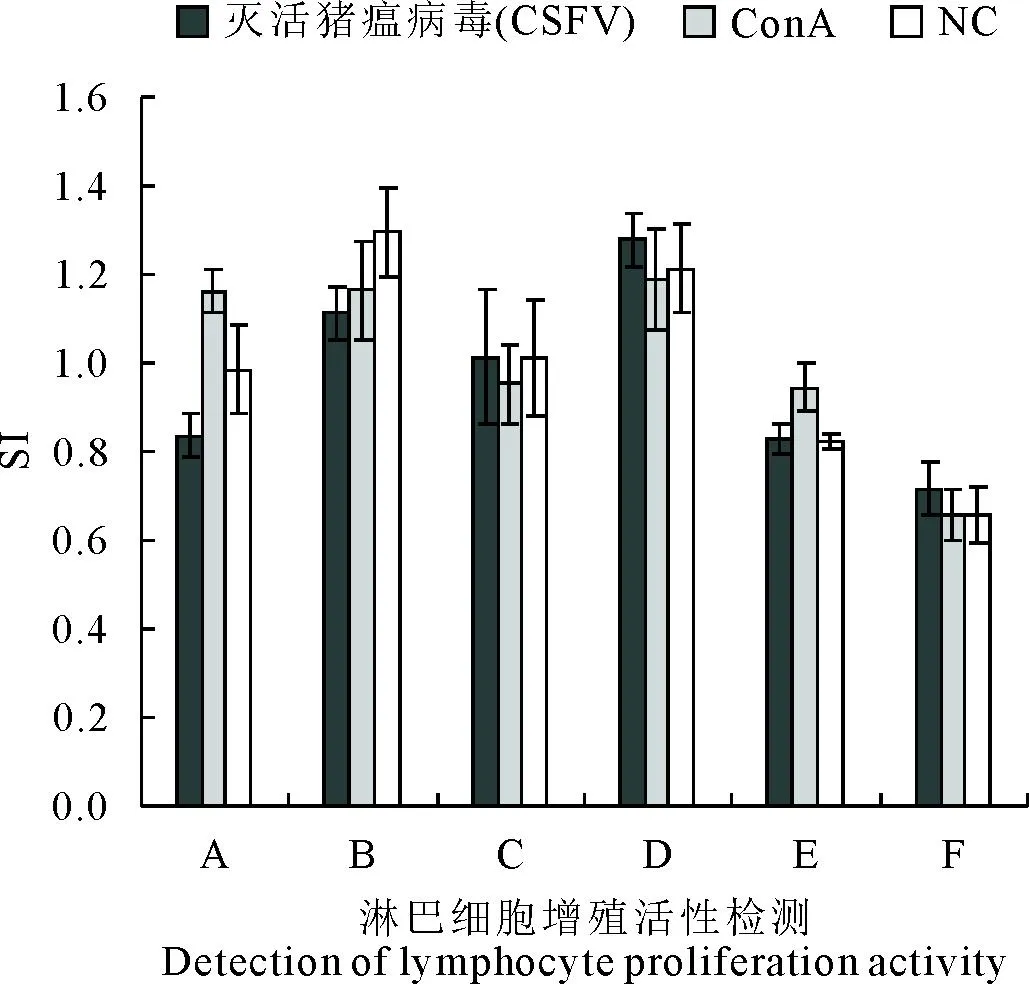
图5 免疫后35 d淋巴细胞增殖活性检测

图6 免疫血清细胞因子检测
3 讨论
囊膜糖蛋白E2是感染动物中激发抗猪瘟抗体的主要靶点,基于CSFV E2蛋白的疫苗研究较多,有利用毕赤酵母表达系统表达E2蛋白[13]、利用杆状病毒表达系统表达E2蛋白[6,14],有在中国仓鼠CHO-dhfr-细胞中表达的[15],还有利用水稻、烟草等植物表达系统生产E2蛋白,接种植物源E2蛋白疫苗的猪显示出有效的中和抗体和免疫保护[10,16-17]。相关研究都证实E2蛋白作为亚单位疫苗靶点的有效性,本研究用杆状病毒表达系统表达经Ni-NAT亲和层析纯化获得CSFV E2蛋白,经硫酸铵沉淀和Ni-NTA亲和层析纯化获得纯度约95%的E2蛋白;通过选择铝佐剂、50V佐剂、201佐剂分别与重组E2蛋白组合制备亚单位疫苗,围绕不同佐剂对猪瘟亚单位疫苗免疫效果的影响开展研究,检测免疫后小鼠血清特异性抗体水平及免疫细胞相关因子水平,发现有佐剂组A、B、C、D组的特异性抗体水平明显优于无佐剂组(E组);50V佐剂(B组)和商品疫苗对照组(D组)亚单位疫苗的特异性抗体水平明显优于201佐剂(C组)和铝佐剂(A组);通常情况下IFN-γ、IL-2表示Th1免疫应答,以IL-4、IL-10表示Th2免疫应答,本研究通过比较细胞因子IL-2、IL-4、IL-10和IFN-γ的水平发现,除了IL-4外,IL-2、IL-10和IFN-γ在各组的含量差异并不显著;而50V佐剂、商品苗和201佐剂亚单位疫苗IL-4的分泌量明显优于其他各组,其中50V佐剂IL-4含量最高,几乎是PBS免疫组的2倍。IL-4是Th2细胞分泌的细胞因子,它也在调节体液免疫和适应性免疫中起关键作用,说明50V佐剂和201佐剂激发的免疫反应以Th2免疫应答为主。
本研究中油包水(W/O)的50V佐剂对E2蛋白的免疫效果提升明显优于其他佐剂,50V佐剂(W/O)组抗体效价是201佐剂(W/O/W)组的2倍,是氢氧化铝无机佐剂组的4倍,在免疫后第21天阻断率已达到60%,明显优于氢氧化铝无机佐剂组40%和ISA201 55%的阻断率,50V佐剂能明显提高CSFV E2蛋白亚单位疫苗的免疫效果。有研究用水佐剂(GEL01)、双相佐剂(ISA201)以及油包水佐剂(ISA植物油)3种佐剂对猪瘟E2基因工程亚单位疫苗免疫效果进行比较研究,发现油包水佐剂(ISA植物油)制备的疫苗与另外2种佐剂制备的疫苗相比,免疫空窗期短,免疫持续期长,中和抗体水平高,在临床上更具优势[18],这一结果与本研究结果基本一致。揭示了不同佐剂类型对CSFV E2蛋白亚单位疫苗免疫在小鼠体内诱发免疫反应的差异,为CSFV E2蛋白亚单位疫苗的研制提供理论依据和技术支撑。
猜你喜欢
心理学探新(2022年1期)2022-06-07 09:16:02
昆明医科大学学报(2021年3期)2021-07-22 07:39:08
幸福(2019年12期)2019-05-16 02:27:44
中成药(2018年10期)2018-10-26 03:40:48
猪业科学(2018年8期)2018-09-28 01:27:52
猪业科学(2018年5期)2018-07-17 05:55:18
湖南中医药大学学报(2016年1期)2016-12-01 04:08:14
兽医导刊(2016年6期)2016-05-17 03:50:36
现代畜牧兽医(2016年11期)2016-02-21 23:11:03
中国当代医药(2015年8期)2015-03-01 02:01:55
