
PAK1在幽门螺杆菌联合MNNG诱导人食管上皮细胞分泌IL-8和GRO-α中的作用
2024-03-16 10:01郭雨松李思瑶陈艳
浙江临床医学 2024年2期
郭雨松 李思瑶 陈艳*
食管癌(esophageal cancer,EC)是全球最常见的消化系统恶性肿瘤之一,严重威胁人类的健康与生命。根据全球癌症统计(global cancer statistics,GLOBOCAN)数据显示,2020 年我国食管癌新发病例数为324,422例,死亡病例数为301,135 例,分别占全球食管癌新发病例和死亡病例的53.70%和55.35%[1]。亚硝胺类化合物具有极强的致癌作用,N-甲基-N’-硝基-N-亚硝基胍(MNNG)是一种具有较强诱变及致癌能力的人工合成化合物,常被用于化学致癌物诱变机理的研究[2]。幽门螺杆菌(Helicobacter pylori,HP)已被证实与众多消化系统疾病的发生密切相关,但其与食管癌的相关性研究甚少。近年来,炎症与肿瘤之间的关系愈发成为研究热点。本课题组前期通过Agilent Human LncRNA芯片检测DNMT1 高表达食管上皮细胞和正常细胞株差异表达的LncRNA 和mRNA,结合生物信息学技术和共表达网络分析发现LncRNA TUG1 的异常表达与幽门螺杆菌感染的上皮信号通路有关[3-4],而PAK1(p21-activated protein kinase 1)为该信号通路的关键因子。CXCR2 是与PAK1 相关的趋化因子受体之一,可参与肿瘤细胞增殖、引起细胞运动及促血管生成[5],其中最知名的配体是IL-8(CXCL8)和GRO-α(CXCL1)。本研究通过制备RNAi和过表达慢病毒并感染人食管上皮细胞构建PAK1 低、高表达HEEC 稳定细胞株,探讨PAK1在HP 联合MNNG 诱导人食管上皮细胞分泌趋化因子IL-8、GRO-α 中的作用,以期为食管癌的发病机制提供理论依据。
1 材料与方法
1.1 材料 人食管上皮细胞系(HEEC)购自宝信生物公司(中国武汉),内参基因、目的基因引物由上海吉凯公司设计与合成,工具载体GV112、GV341 购自上海吉凯公司。幽门螺杆菌标准菌株(ATCC26695)来源于广东省微生物菌种保藏中心,MNNG 分析纯购自上海罗恩公司,PMI1640 培养基购自美国HyClone 公司,磷酸缓冲盐溶液(phosphate buffer saline,PBS)、胎牛血清(fetal calf serum,FBS)、0.25%胰蛋白酶、青链霉素混合液均购自美国Gibco 公司,二甲基亚砜(dimethyl sulfoxide,DMSO)购自北京索莱宝公司,布氏肉汤购自杭州微生物试剂有限公司,甘油三酯购自广东西陇科学试剂公司,RNAiso Plus、PrimeScript™ RT Master Mix(Perfect Real Time)和TB Green® Premix Ex Taq™(Tli RNaseH Plus)均购自日本Takara 公司。GRO-α 及IL-8 ELISA 试剂盒购自欣博盛公司。
1.2 仪器 CO2恒温孵育箱、生物安全柜和超低温冰箱购自美国Thermo Scientific 公司,生物超净台购自上海天呈公司,低速自动平衡离心机购自北京京立公司,低温离心机购自德国Eppendorf 公司,生物厌氧培养箱购自浙江义乌冷冻机总厂,紫外分光光度计购自北京普析通用仪器有限公司,PCR 仪(Mastercycler)和Realtime PCR 仪(RealpleX2)均购自德国Eppendorf 公司。
1.3 方法 (1)PAK1 低表达HEEC 稳定细胞株的构建:通过制备RNAi 慢病毒并感染HEEC 构建PAK1 低表达稳定细胞株(PAK1-KD)。引物由上海吉凯公司设计与合成、工具载体GV112 购自上海吉凯公司。具体步骤为设计目的基因干扰靶点→干扰靶点克隆构建至病毒载体上→测序确保靶点序列一致→质粒扩增、质检→质粒转染细胞包装病毒→病毒收获、浓缩、纯化→病毒质检、分装→慢病毒预感染→病毒感染→目的基因的干扰效率检测。引物序列见表1、2。(2)PAK1 高表达HEEC 稳定细胞株的构建:通过制备过表达慢病毒载体并感染HEEC 构建PAK1 高表达HEEC 稳定细胞株(PAK1-OE)。目的基因引物由上海吉凯公司设计与合成、工具载体GV341 购自上海吉凯公司。具体步骤为设计目的基因获取→重组质粒构建至病毒载体上→测序确保序列一致→质粒扩增、质检→质粒转染细胞包装病毒→病毒收获、浓缩、纯化→病毒质检、分装→慢病毒预感染→病毒感染→目的基因过表达效率检测。目的基因引物序列见表3。(3)细胞染毒:PAK1-KD 和PAK1-OE 细胞分别分为HP+MNNG 联合染毒组、HP 染毒组和MNNG 染毒组并设立空白对照组,共8 组。将PAK1-KD/ PAK1-OE 细胞接种于6 孔板中,当细胞铺满70%~80%瓶底面积时,按感染复数(MOI)=100 ∶1,即细菌∶细胞=100 ∶1 加入至细胞中进行共培养,加入MNNG 溶液至终浓度为IC50[6]进行染毒。(4)IL-8、GRO-α 含量的测定:各组细胞染毒3 h、6 h 和9 h 后弃去上清液,用PBS 清洗3 次,用含庆大霉素的无血清培养基培养48 h 后做支原体检测,避免支原体污染,收集上清液,严格按ELISA 试剂盒说明书方法进行测定,每组设3 个复孔。

表1 PAK1 RNAi靶点

表2 PAK1 RNAi合成引物信息
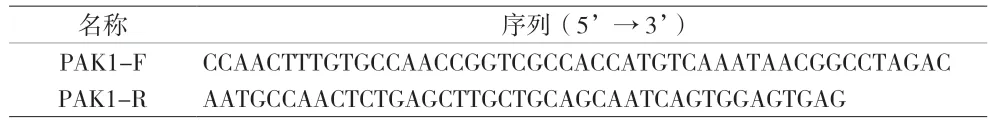
表3 PAK1合成引物信息
1.4 统计学分析 采用SPSS 26.0 统计学软件。计量资料进行正态性检验,均为正态分布,采用(±s)进行统计描述。两独立样本间比较采用独立样本t检验,多组间比较采用单因素方差分析,两两比较采用Bonferroni 法。用Graphpad Prism 8 绘图软件绘图。检验水准α=0.05。
2 结果
2.1 HEEC PAK1 低、高表达稳定株的构建 经实时荧光定量PCR 验证,与PAK1-NC 组相比,PAK1-KD 基因敲减效率达84.1%(t=14.627,P=0.004),PAK1-OE组PAK1 基因的表达丰度是PAK1-NC 组的1,101.646 倍(t=16.104,P=0.004),见表4,图1。表明成功构建了PAK1 低、高表达人食管上皮细胞稳定株。2.2 PAK1 在HP 联合MNNG 染毒促进细胞IL-8 分泌中的作用 PAK1-KD 与PAK1-OE 细胞染毒3 h、6 h和9 h 后,各染毒组细胞IL-8 含量随染毒时间增加,经趋势性检验线性趋势显著(PAK1-KD 细胞:F对照组=2.682,P=0.153;FMNNG=20.425,P=0.004;FHP=40.862,P=0.001;FHP+MNNG=25.181,P=0.002。PAK1-OE 细胞:F对照组=2.190,P=0.189;FMNNG=39.286,P=0.001;FHP=36.478,P=0.001;FHP+MNNG=193.692,P<0.001)。相同染毒时间,各组细胞IL-8 含量差异有统计学意义(PAK1-KD 细胞:F3h=126.292,P<0.001;F6h=281.997,P<0.001;F9h=232.455,P<0.001。PAK1-OE 细胞:F3h=486.373,P<0.001;F6h=588.701,P<0.001;F9h=378.793,P<0.001),且均出现联合染毒组>HP 染毒组>MNNG 染毒组>对照组,差异有统计学意义(均P<0.05)。相同染毒时间,与PAK1-KD 相比,PAK1-OE 显著促进联合染毒组、HP 染毒组和MNNG 染毒组细胞分泌IL-8(联合组:t3h=3.377,P=0.028;t6h=3.017,P=0.039;t9h=3.394,P=0.027;HP 染毒组:t3h=2.863,P=0.046;t6h=2.909,P=0.044;t9h=3.629,P=0.022;MNNG 染毒组:t3h=2.964,P=0.043;t6h=3.014,P=0.040;t9h=2.837,P=0.047)。对照组间无显著差异(均P>0.05)。见图2。说明PAK1 高表达可以促进HP 联合MNNG 诱导人食管上皮细胞分泌IL-8。
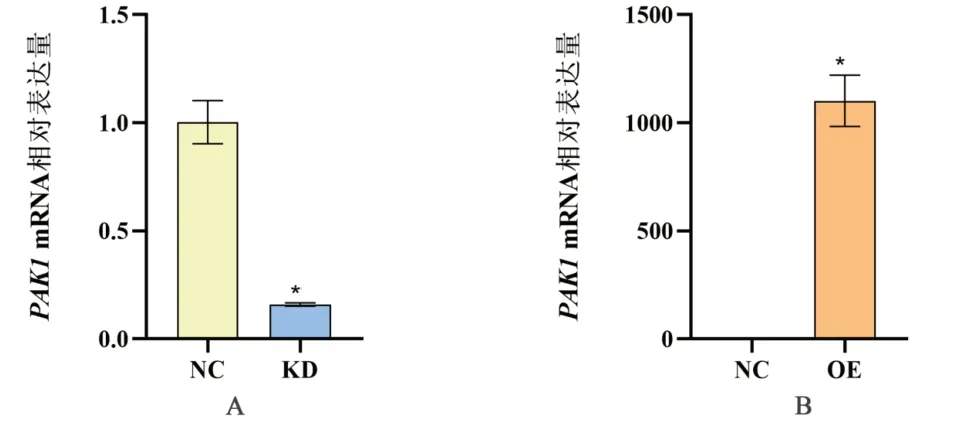
图1 PAK1低、高表达HEEC稳定细胞株构建效率。注:与NC组比较,*P<0.01
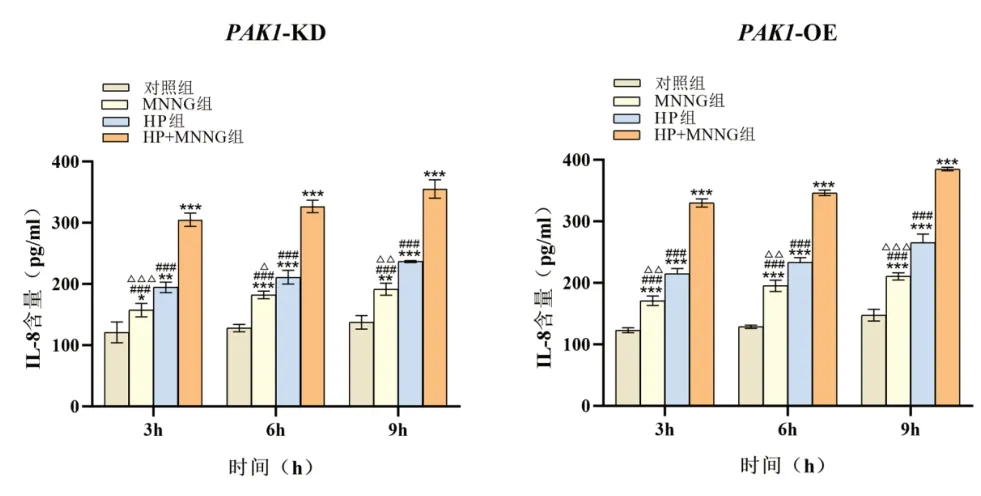
图2 PAK1 对各处理组不同染毒时间细胞分泌IL-8 的影响。注:与对照组比较,*P<0.05,** P<0.01,***P<0.001;与HP+MNNG 组比较,#P<0.05,##P<0.01,###P<0.001;与HP 组比较,△P<0.05,△△P<0.01,△△△P<0.001
表4 慢病毒感染后PAK1基因相对表达量[(±s),2-ΔΔCt]

表4 慢病毒感染后PAK1基因相对表达量[(±s),2-ΔΔCt]
注:细胞标记说明:NC-空载体对照组、KD-低表达组、OE-高表达组
?
2.3 PAK1 在HP 联合MNNG 染毒促进细胞分泌GRO-α 中的作用 PAK1-KD 与PAK1-OE 细胞染毒3 h、6 h 和9 h 后,各染毒组细胞GRO-α 含量均随染毒时间增加,经趋势性检验线性趋势显著(PAK1-KD细胞:F对照组=2.679,P=0.153;FMNNG=38.283,P=0.001;FHP=28.980,P=0.002 ;FHP+MNNG=121.938,P<0.001。PAK1-OE 细胞:F对照组=0.597,P=0.469;FMNNG=58.925,P<0.001 ;FHP=40.543,P=0.001 ;FHP+MNNG=246.745,P<0.001)。相同染毒时间,各组细胞分泌GRO-α 含量差异有统计学意义(PAK1-KD 细胞:F3h=226.755,P<0.001 ;F6h=835.844,P<0.001 ;F9h=1182.142,P<0.001。PAK1-OE 细胞:F3h=1523.939,P<0.001;F6h=675.016,P<0.001;F9h=1654.557,P<0.001),且均出现联合染毒组>HP 染毒组>MNNG 染毒组>对照组,差异有统计学意义(均P<0.01)。相同染毒时间,与PAK1 低表达相比,PAK1 高表达显著促进联合染毒组、HP 染毒组和MNNG 染毒组细胞分泌GRO-α(联合组:t3h=6.645,P=0.004;t6h=9.059,P=0.012;t9h=11.523,P<0.001;HP 染毒组:t3h=3.014,P=0.039;t6h=4.034,P=0.016;t9h=3.519,P=0.024;MNNG 染毒组:t3h=3.308,P=0.030;t6h=3.091,P=0.037;t9h=4.063,P=0.015)。对照组间无显著差异(均P>0.05)。见图3。说明PAK1 高表达可以促进HP 联合MNNG 诱导人食管上皮细胞分泌GRO-α。
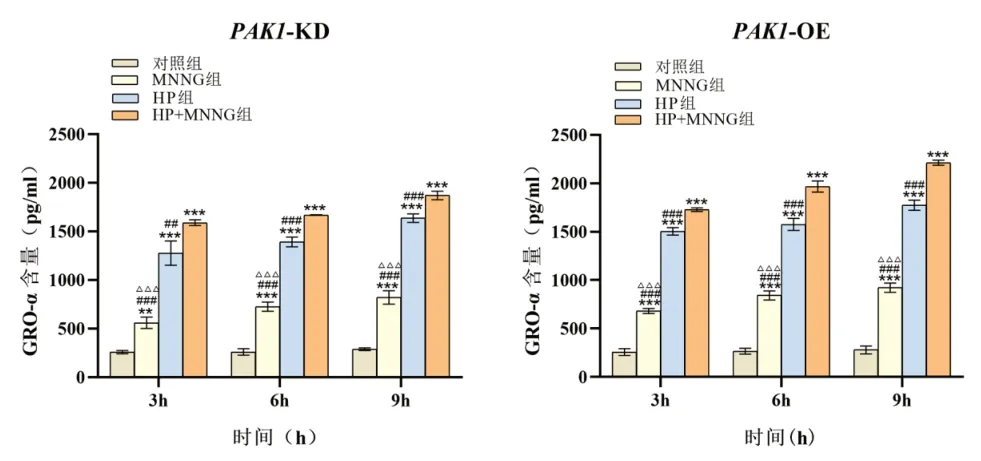
图3 PAK1 对各处理组不同染毒时间细胞分泌GRO-α 的影响。注:各组与对照组比较,*P<0.05,** P<0.01,***P<0.001;各组与HP+MNNG 组比较,#P<0.05,##P<0.01,###P<0.001;各组与HP 组比较,△P<0.05,△△P<0.01,△△△P<0.001
3 讨论
食管癌是一种与炎症相关的疾病,炎症在食管癌的发生、发展中发挥重要作用。有研究表明亚硝胺类化合物是食管癌的重要致病因素,NF-κB 通路通常在MNNG 诱导食管炎中起关键作用[7]。此外,某些细菌产物如HP 亦具有促肿瘤作用,当机体受到细菌感染时会引起促炎介质释放增加,进而改变趋化因子和细胞因子的表达,促进细胞生长与侵袭,导致肿瘤的发生[8]。已有研究表明HP 可导致慢性胃炎,并增加消化性溃疡、黏膜相关淋巴组织淋巴瘤(MALT)和胃癌的发病风险[9],然而关于HP 与食管癌的相关性研究甚少,且结论也不一致。吉胜利等[10]研究HP 感染与食管鳞癌相关性时,发现在食管鳞癌患者组中HP 感染阳性率最高,明显高于重度不典型增生患者组、轻-中度不典型增生患者组、慢性食管炎患者组和轻度不典型增生患者组,提示HP 感染可影响食管鳞癌的发生与发展。研究[11]表明HP 感染可提高食管癌细胞的增殖能力,导致细胞形态不规则及细胞间隙扩大,促进食管癌的发生、发展。PAK1 作为HP 感染上皮信号通路中的重要分子,其激活可引发下游IL-8、GRO-α 和其他中性粒细胞趋化因子的改变[12],引起促炎反应及趋化效应,促使肿瘤的发生。TRAN 等[13]研究表明HP 可通过EGFR、MAPK 和JAK/STAT 信号通路诱导趋化因子GRO-α 和IL-8 的表达。IL-8 及其受体在肿瘤的发展过程中发挥了关键作用,IL-8 可与其受体CXCR1 及CXCR2 相互作用参与肿瘤细胞增殖[14]、促进血管生成及诱导炎症反应等。范玉宏等[15]研究发现在食管鳞癌患者组织中GRO-α 及其受体CXCR2 的mRNA 和蛋白表达水平显著高于正常组织,说明食管黏膜在发生癌变过程中,GRO-α 和CXCR2 参与启动趋化过程来介导食管鳞癌的发生。作为癌症发展过程中的重要趋化因子,IL-8 和GRO-α 可通过与CXCR2 受体相结合的方式,参与激活TAK1/NF-κB 信号传导[16],促进癌细胞的增殖和转移。
PAK1 是一个潜在的癌基因,目前已在多种人类恶性肿瘤如乳腺癌、肺癌、淋巴瘤及黑素瘤等中观察到PAK1 基因和PAK1 蛋白的过度表达[17],其在调节细胞周期、细胞运动、存活和细胞生长信号转导、转化方面发挥关键作用[18]。本研究发现,PAK1 高表达显著促进了HP 联合MNNG 诱导人食管上皮细胞分泌趋化因子IL-8 和GRO-α,有理由认为HP 联合MNNG 可激活PAK1,引起下游炎症趋化因子IL-8 和GRO-α 分泌增加,引发食管癌相关不良反应。
由于食管癌发生的复杂性,本研究采用短期染毒方法,仅在分子水平观察到PAK1 对HP 联合MNNG 诱导的趋化因子IL-8 和GRO-α 的调控作用,在以后的研究中,需增加更长染毒时间点或慢性染毒人食管上皮细胞,以深入探讨PAK1 在HP 联合MNNG 致食管癌发病中的作用机制。
猜你喜欢
科学导报·学术(2020年19期)2020-07-09
实验动物科学(2019年6期)2019-12-02
环境与生活(2018年7期)2018-09-10
首都公共卫生(2017年3期)2017-11-29
中国医药生物技术(2015年4期)2015-12-26
医学研究杂志(2015年11期)2015-06-10
中国当代医药(2015年16期)2015-03-01
中国医药导报(2015年27期)2015-02-28
现代检验医学杂志(2015年4期)2015-02-06
现代检验医学杂志(2015年6期)2015-02-06
