
湖泊深水暂养过程中鳙鱼品质变化研究
2024-03-11 03:43凌俊陈小雷江河汪长祥童开满潘庭双胡玉婷段国庆周华兴汪焕
安徽农业科学 2024年3期
凌俊 陈小雷 江河 汪长祥 童开满 潘庭双 胡玉婷 段国庆 周华兴 汪焕

摘要 為了研究天然水体长成鳙鱼在不同月份暂养过程中鳙鱼体形、肌肉营养、滋味、免疫及抗氧化物质的变化,采集花亭湖5月、8月、11月和次年3月暂养鳙鱼进行系统研究。结果显示,从体形来看,暂养处理对鳙鱼有瘦身效果;不同月份鳙鱼肌肉基本营养成分含量有差别,暂养处理后,鳙鱼肌肉蛋白质和脂肪含量降低,水分和灰分含量升高;脂肪酸组成发生变化,主要表现在饱和脂肪酸(SFA)含量降低,多不饱和脂肪酸(PUFA)含量升高;肌肉免疫和抗氧化成分含量随着暂养时间的延长显著降低;游离氨基酸总量随着暂养时间的延长不断提高,呈现甜味氨基酸占比不断提高;呈味核苷酸总量提高。
关键词 鳙鱼;暂养;营养;免疫;抗氧化;滋味
中图分类号 S965.114 文献标识码 A 文章编号 0517-6611(2024)03-0080-08
doi:10.3969/j.issn.0517-6611.2024.03.019
Study on the Quality Changes of Bighead Carp During Temporary Rearing in Deep Lake
Abstract In order to study the changes in body shape,muscle nutrition,taste,immunity,and antioxidant substances of adult bighead carp in natural water bodies during temporary rearing in different months,a systematic study was conducted on the temporary cultivation of bighead carp in Huating Lake in May,August, November and March of the following year.The results showed that from the perspective of body shape,temporary feeding treatment had a slimming effect on bighead carp;The basic nutritional content of bighead carp muscle varies in different months.After temporary cultivation,the protein and fat content of bighead carp muscle decreases,while the water and ash content increases;The composition of fatty acids changes,mainly manifested in a decrease in saturated fatty acid (SFA) content and an increase in polyunsaturated fatty acid (PUFA) content;The content of muscle immunity and antioxidant components significantly decreases with the prolongation of temporary maintenance time;The total amount of free amino acids continues to increase with the extension of temporary incubation time,and the proportion of sweet amino acids continues to increase;The total amount of flavor nucleotides increased.
Key words Bighead carp;Temporary rearing;Nutrition;Immunization;Antioxidant;Taste
湖泊、水库等大水面渔业,是中国淡水渔业的重要组成部分,在建设水域生态文明、保障优质水产品供给、推动产业融合发展、促进农民增收等方面发挥重要作用,其产量约占全国淡水渔业产量的20%。按照《关于加快推进水产养殖业绿色发展的若干意见》和《关于推进大水面生态渔业发展的指导意见》要求,大水面的主要利用方式是发展生态渔业。鳙鱼作为典型的滤食性鱼类,不仅可以通过滤食浮游生物带走水体过量氮磷、有效控制水体中浮游生物过度繁殖,还能够将水体中的水生生物资源转化为优质水产品,在生态渔业中发挥着巨大作用。然而,鳙鱼在生长过程中,由于受水体中放线菌、真菌和藻类等物质沉积影响,肌肉存有一定土腥味,为消费者所不喜[1]。近年来,随着市场对优质蛋白的需求增加及消费市场的升级,急需提升鳙鱼品质以适应市场需求。
原位暂养可以降低鳙鱼体脂肪含量和土腥味,提高鱼肉品质和风味,如在循环水养殖系统下,通过饥饿暂养可以提升其肌肉品质[2]。对于大水面捕捞的鳙鱼而言,在特定深水区域、固定设施中暂养一段时间是普遍被采用的一种大水面鱼上市前的预处理手段,该暂养处理模式兼具经济价值和生态效益,具有广阔的发展前景。但是深水暂养处理后,鳙鱼鱼肉品质随着暂养时间推进如何改变却缺乏数据支撑;鳙鱼一年四季均可以上市,不同季节大水面捕捞的鳙鱼暂养后鱼肉品质是否具有差别,目前还未有相关的报道,急需更多的理论基础和数据依据为深水原位暂养鳙鱼生产实践提供支撑。
该研究通过分析花亭湖暂养过程中鳙鱼形态、免疫、基本营养成分、游离氨基酸、脂肪酸组成、呈味核苷酸等变化,为鳙鱼深水暂养提供科学依据,指导精准施策,降低企业暂养风险,提高暂养技术水平和效益。
1 材料與方法
1.1 材料与仪器
有机鳙鱼,采自太湖县花亭湖;5′-肌苷酸(IMP)标准品、5′-鸟苷酸(GMP)标准品、5′-腺苷酸(AMP)标准品,购自上海安谱实验科技股份有限公司;丙二醛测试盒、超氧化物气化酶测试盒、过氧化氢酶测试盒、总蛋白测试盒,购自南京建成生物工程研究所。
GC-2010 Plus气相色谱仪,日本岛津公司;L-8900全自动氨基酸分析仪,日本日立公司;TU-1901紫外-可见分光光度计,北京普析通用仪器公司;FA25型高速分散机,弗鲁克(上海)公司;CR22G型高速冷冻离心机,日本日立公司。
1.2 试验条件和方法
1.2.1 暂养设施设置及日常管理。
于花亭湖水深大于12 m的水层处构建“田”字框架结构,使用锚泊装置固定框架主体。框架结构内置放暂养箱体(由双层网衣构成,内层:长8~10 m、宽5~6 m、深9~10 m;外层:长宽同内层,深度较内层网衣长1 m),使用浮力装置保持整体框架高出水平面30~40 cm。在微流水条件下每平方箱体暂养鳙鱼80~100 kg。放养前进行鱼体消毒。在暂养过程中,不投饵,不施肥,早晚巡查,日常监控鱼体活动情况。
1.2.2 样品采集与处理。
于2019年5月12、18和25日抽取本批次花亭湖深水暂养初期(5-1)、暂养中期(5-2)和暂养末期(5-3)的鲜活鳙鱼;同年8月11、18和25日抽取本批次花亭湖深水暂养初期(8-1)、暂养中期(8-2)和暂养末期(8-3)的鲜活鳙鱼;同年11月15、22和29日抽取本批次花亭湖深水暂养初期(11-1)、暂养中期(11-2)和暂养末期(11-3)的鲜活鳙鱼;次年3月11、18和25日抽取本批次花亭湖深水暂养初期(3-1)、暂养中期(3-2)和暂养末期(3-3)的鲜活鳙鱼。每次取样设10个平行,鳙鱼体重(3.00±0.50) kg。样品活体充氧运输至安徽省农业科学院水产研究所实验室,断鳃致死后取鳙鱼背部和腹部肌肉,分别混匀后,按照不同检测项目要求制作样品,-80 ℃冷藏备检测用。
1.2.3 测定方法。
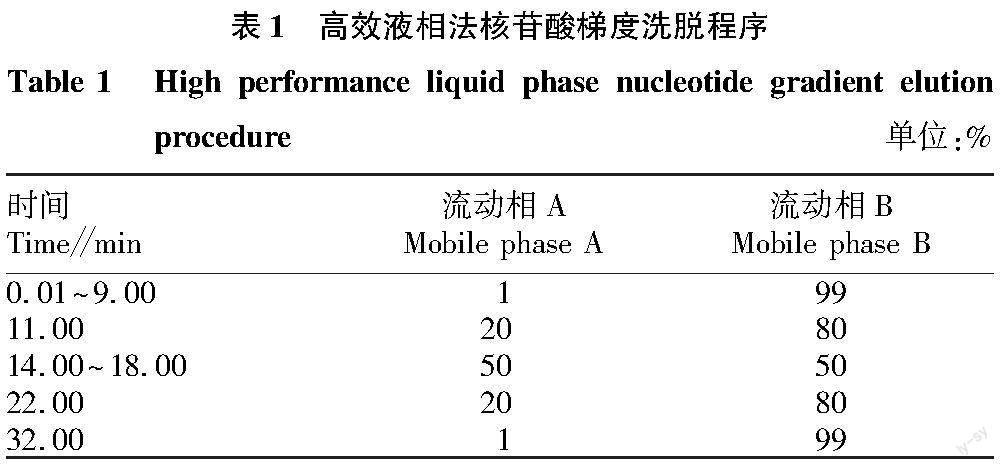
①形态测定。参照李武辉等[3]的方法,稍作修改。采用游标卡尺、直尺测量,精确到0.01 cm。测量参数分为2类,一类是传统形态学可量性状,即全长(L)、体长(L1)、头长(L2)、头宽(W1),体质量(T);另一类为框架数据,测量参数为:胸鳍起点—吻端(D1),胸鳍起点—腹鳍起点(D2),胸鳍起点—头背部末端(D3),吻端—头背部末端(D4),腹鳍起点—头背部末端(D5),腹鳍起点—臀鳍起点(D6),腹鳍起点—背鳍起点(D7),腹鳍起点—背鳍末端(D8),头背部末端—背鳍起点(D9),臀鳍起点—臀鳍末端(D10),臀鳍起点—背鳍末端(D11),臀鳍起点—尾鳍背部起点(D12),背鳍起点—背鳍末端(D13),臀鳍末端—尾鳍腹部起点(D14),臀鳍末端—尾鳍背部起点(D15),背鳍末端—尾鳍背部起点(D16),尾鳍腹部起点—尾鳍背部起点(D17)。框架系统所取定位点如图1所示。②基本营养成分测定。粗蛋白质含量测定依据GB 5009.5—2016,粗脂肪含量的测定依据GB 5009.6—2016,水分含量的测定依据GB 5009.3—2016,灰分含量的测定依据GB 5009.4—2016。③肌肉脂肪酸含量测定。依据GB 5009.168—2016。④肌肉丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和总蛋白(TP)含量测定。分别称取定量新鲜鱼肉样品,加入5倍体积去离子水,10 000 r/min匀浆1 min后分别参照对应试剂盒说明书测定。⑤肌肉游离氨基酸和呈味核苷酸含量测定。参照陈小雷等[4]的方法。流动相洗脱程序见表1。
1.3 数据处理
试验数据采用平均值±标准差表示,利用SPSS 17.0统计软件进行方差分析、多重比较和相关性分析,差异显著水平为0.05。
2 结果与分析
2.1 暂养对花亭湖鳙鱼形态的影响
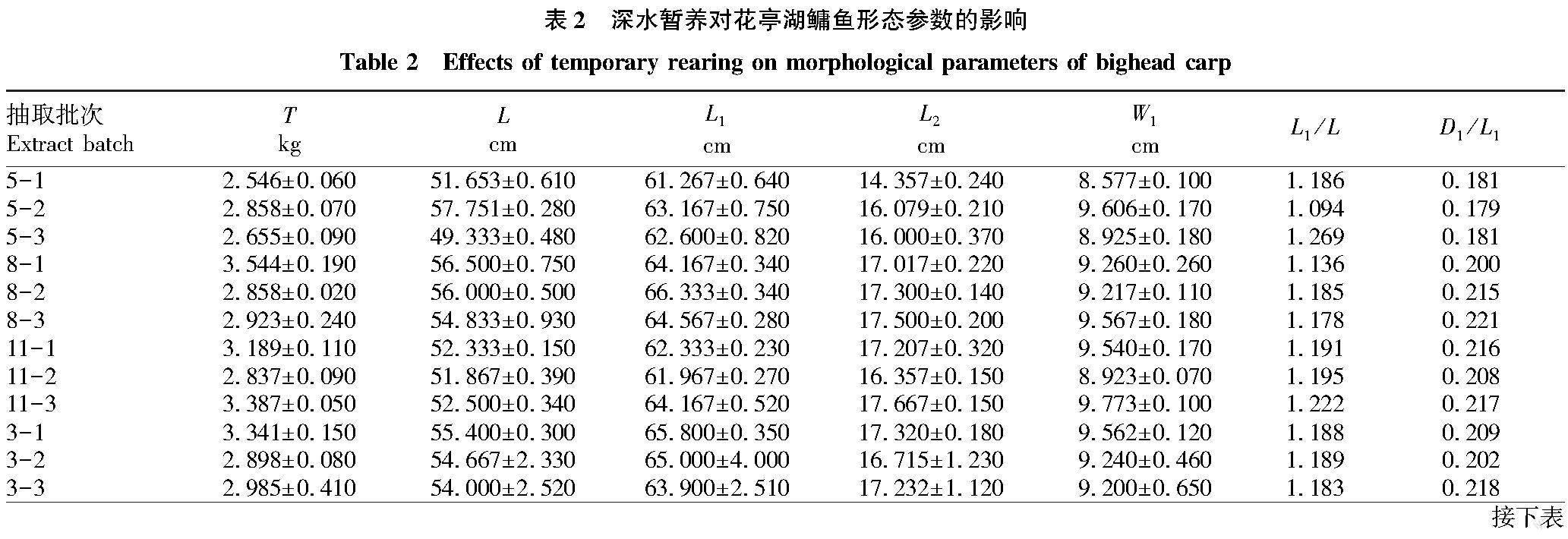
借助可数性状及可量性状进行单一比较的传统形态学分析方法,对物种种水平的判别有效,对形态相似度高的群体却难以准确判别。Bookstein[5]提出的框架系统分析方法比传统形态学分析方法在反应形态差异方面更有效。为了消除鳙鱼规格对形态的影响,将各参数原始值除以体长值予以校正。从表2可以看出,鳙鱼体形在暂养过程中发生了一些变化,尤其以D5/L1、D7/L1、D8/L1、D11/L1、D12/L1和D15/L1变化最为明显,不同月份暂养过程中,D5/L1、D7/L1、D8/L1、D11/L1、D12/L1和D15/L1均呈现减小趋势。
2.2 暂养对鳙鱼肌肉免疫和氧化指标的影响
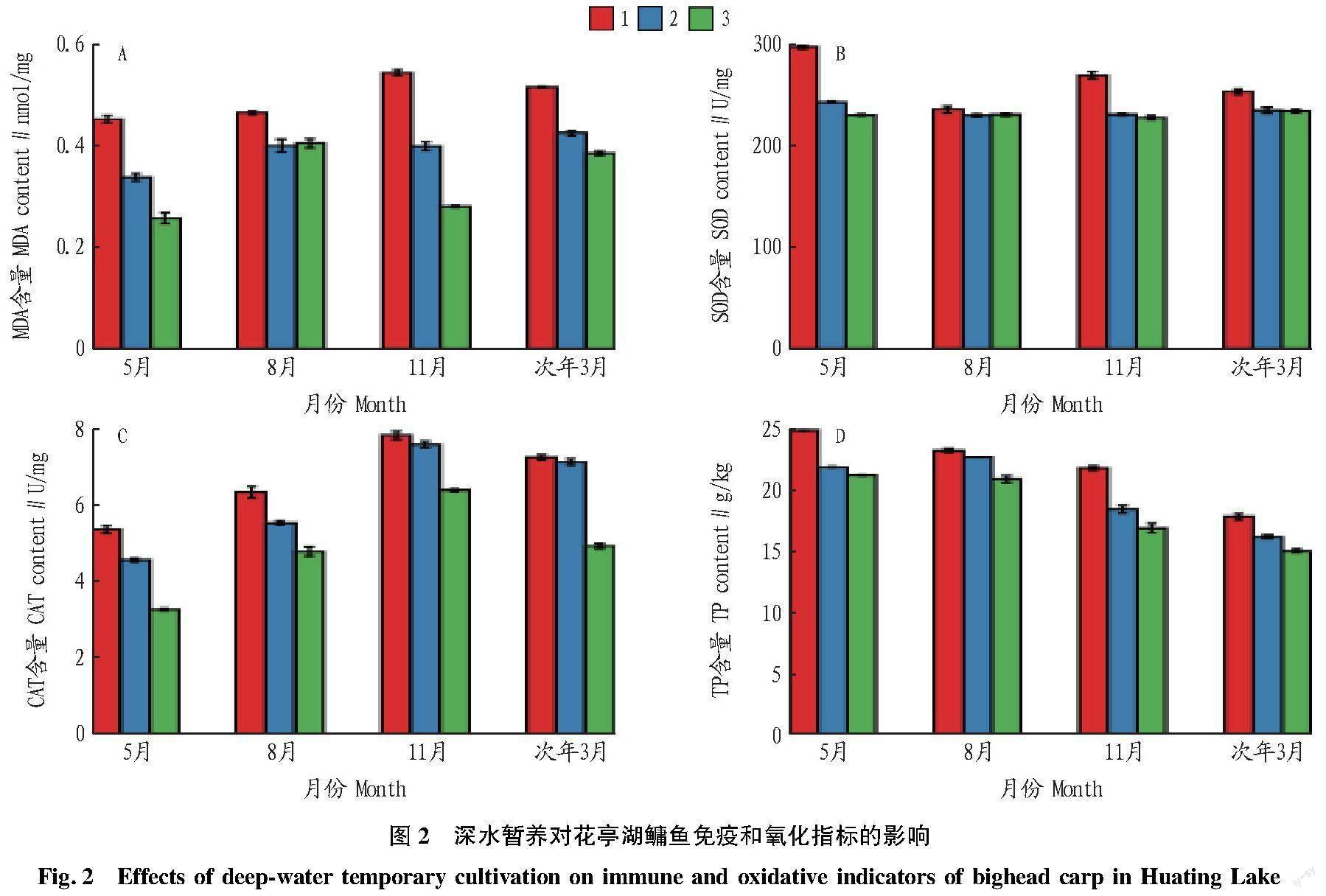
暂养过程中鳙鱼肌肉MDA、SOD、CAT和TP含量变化如图2所示。不同月份暂养的鳙鱼肌肉中MDA、SOD、CAT和TP含量差异显著。暂养过程中,鳙鱼肌肉MDA、SOD、CAT和TP含量随着暂养时间的延长均呈现降低的趋势。
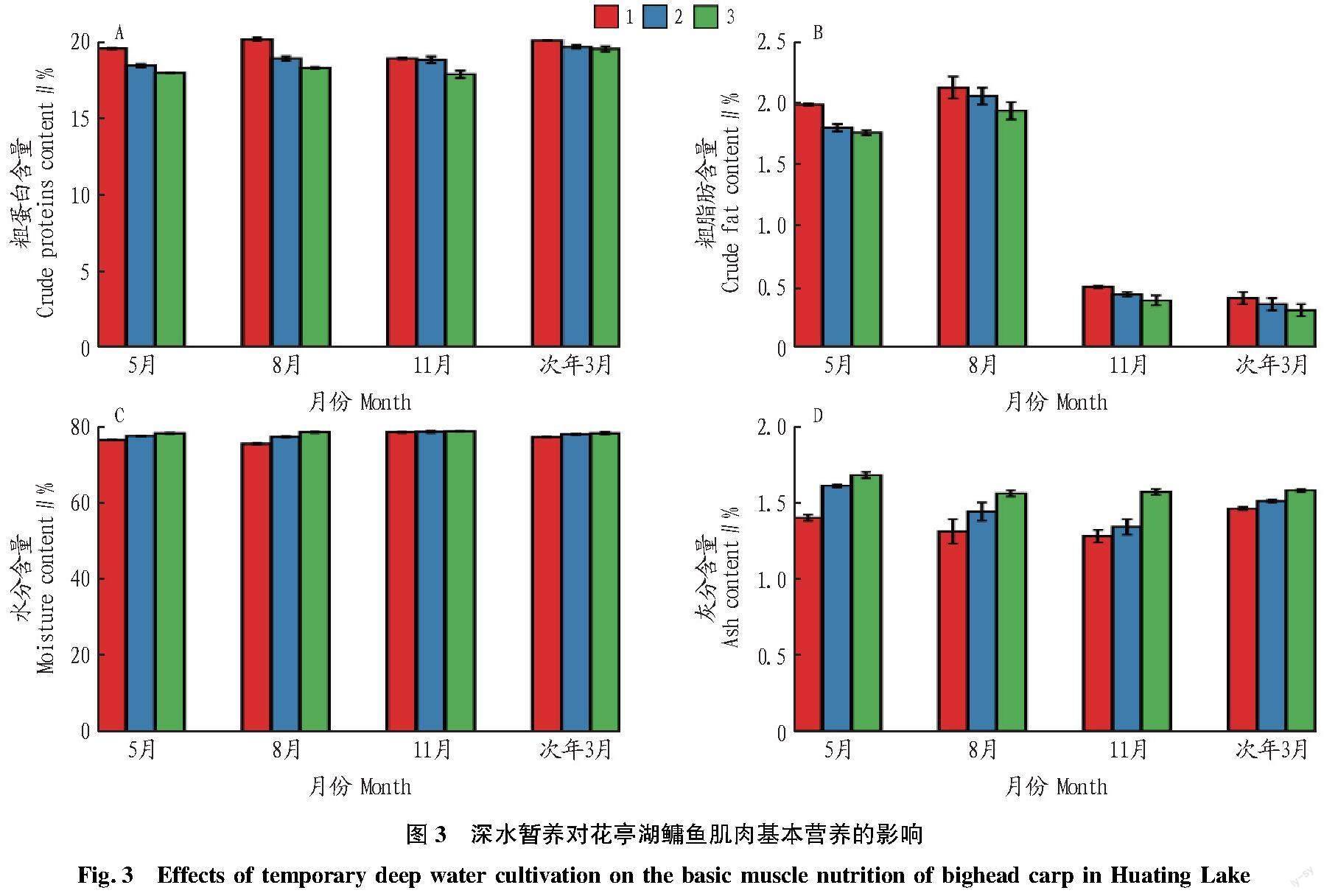
2.3 暂养对花亭湖鳙鱼肌肉基本营养的影响
从图3可以看出,不同月份(5、8、11月和次年3月)之间鳙鱼肌肉粗蛋白、粗脂肪、水分和灰分含量差异显著。其中,鳙鱼肌肉粗蛋白含量次年3月最高,粗脂肪含量8月最高,水分含量11月最高,灰分含量5月最高。随着暂养时间的延长,鳙鱼肌肉粗蛋白和脂肪含量降低,其中,8月鳙鱼蛋白质降低幅度最大,为9.39%;次年3月鳙鱼脂肪降低幅度最大,为25.00%。暂养过程中鳙鱼肌肉水分和灰分含量显著升高。
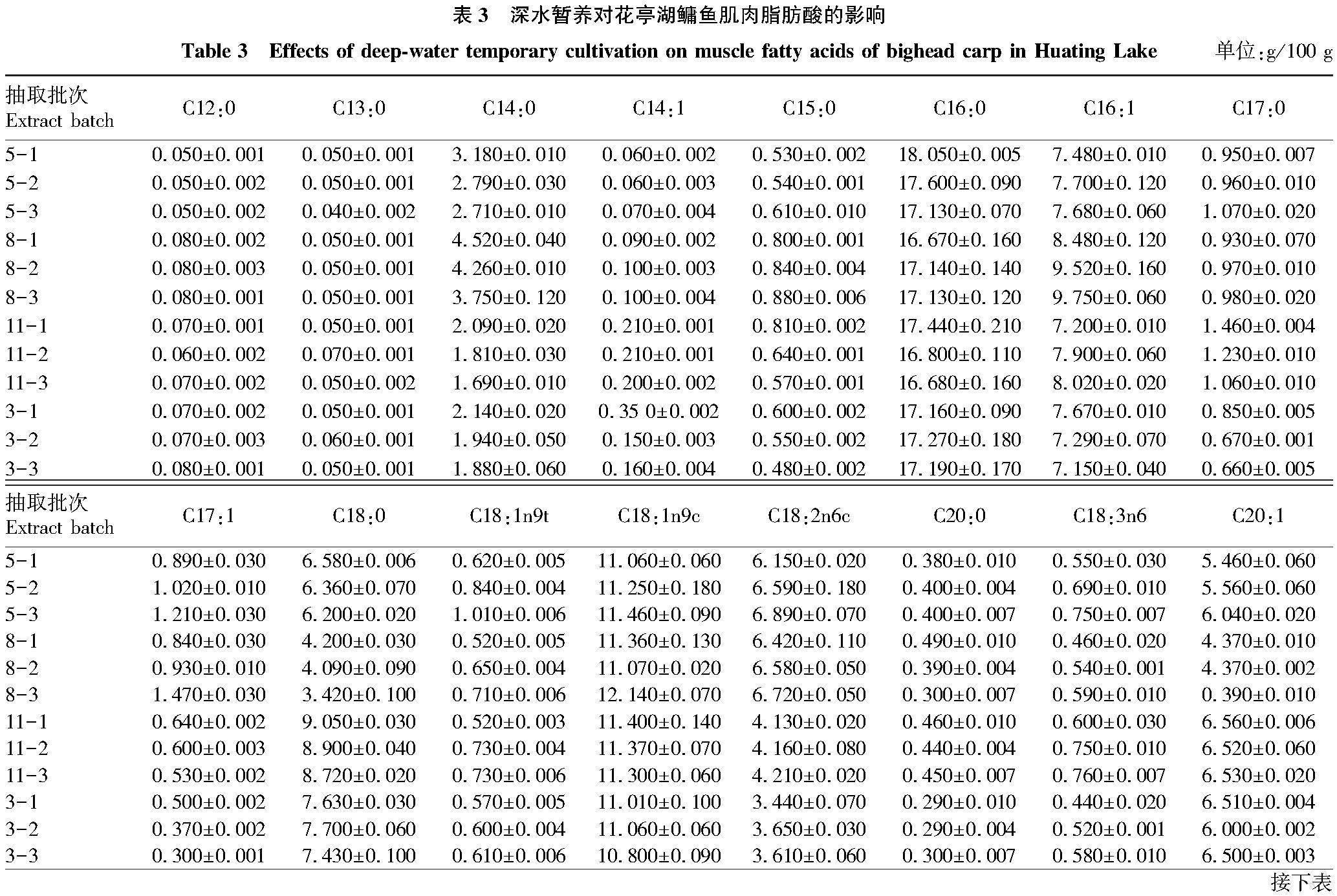
2.4 暂养对鳙鱼肌肉脂肪酸的影响
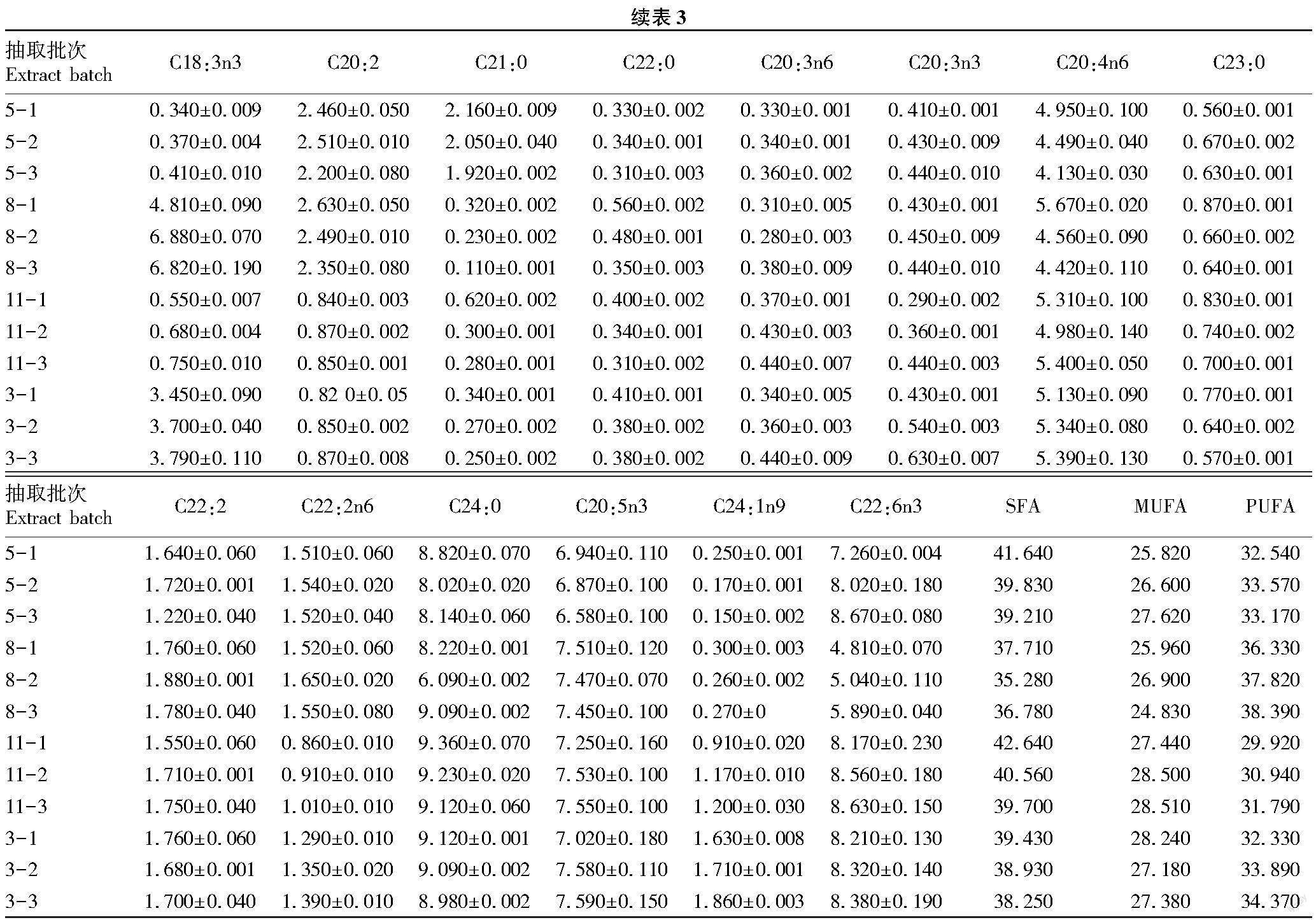
不同季节鳙鱼暂养过程脂肪酸含量变化如表3所示。鳙鱼肌肉中共检测出30种脂肪酸,碳链长度在12碳至24碳之间。其中有12种饱和脂肪酸(SFA)、7种单不饱和脂肪酸(MUFA)、11种多饱和脂肪酸(PUFA)。鳙鱼肌肉主要脂肪酸为C16:0、C16:1、C18:0、C18:1n9、C18:2n6、C20:1、C20:4n6、C24:0、C20:5n3和 C22:6n3。随着暂养时间的延长,5月鳙鱼肌肉SFA总量由41.640 g/100 g降低至39.210 g/100 g,MUFA总量由25.820 g/100 g升高至27.620 g/100 g,PUFA总量由32.540 g/100 g升高至33.170 g/100 g;8月SFA总量由37.710 g/100 g降低至36.780 g/100 g,MUFA总量由25.960 g/100 g降低至24.830 g/100 g,PUFA总量由36.330 g/100 g升高至38.390 g/100 g;11月SFA总量由42.640 g/100 g降低至39.700 g/100 g,MUFA总量由27.440 g/100 g升高至28.510 g/100 g,PUFA总量由29.920 g/100 g升高至31.790 g/100 g;次年3月SFA总量由39.430 g/100 g降低至38.250 g/100 g,MUFA总量由28.240 g/100 g降低至27.380 g/100 g,PUFA总量由32.330 g/100 g升高至34.370 g/100 g。
2.5 暂养对鳙鱼肌肉游离氨基酸的影响
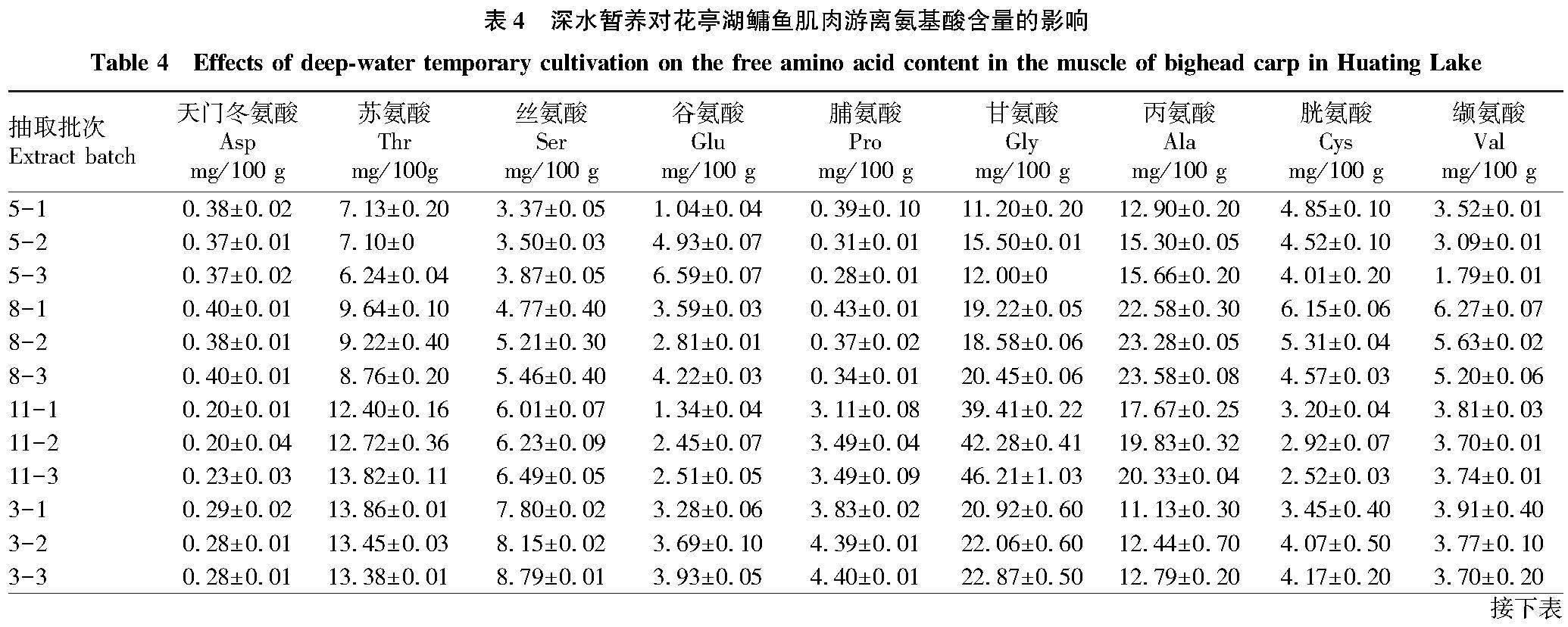
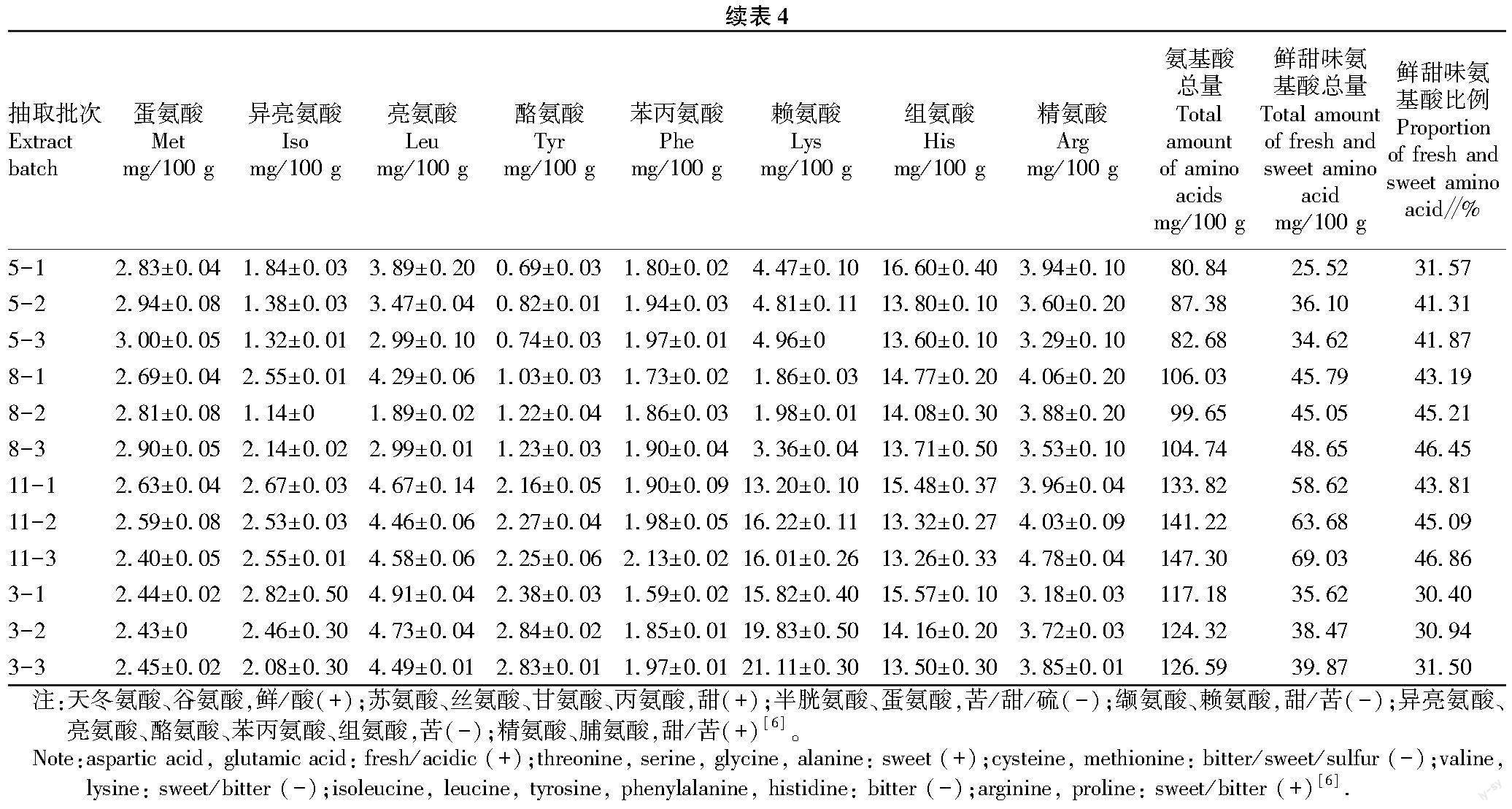
暂养过程中鳙鱼肌肉游离氨基酸含量变化如表4所示。鳙鱼肌肉中主要游离氨基酸为苏氨酸、丝氨酸、甘氨酸、丙氨酸、胱氨酸、缬氨酸、亮氨酸、赖氨酸、组氨酸和精氨酸,占游离氨基酸总量88%以上。随着暂养时间的延长,游离氨基酸总量变化显著,5月鳙鱼肌肉游离氨基酸总量从80.84 mg/100 g升高至87.38 mg/100 g后降至82.68 mg/100 g,8月从106.03 mg/100 g波动降至104.74 mg/100 g,11月从133.82 mg/100 g升高至149.00 mg/100 g,次年3月从117.18 mg/100 g升高至126.59 mg/100 g。
呈鲜甜味游离氨基酸的含量变化趋势与总氨基酸含量变化趋势基本相同,但呈鲜甜味游离氨基酸占比在暂养过程中不断升高,5月鳙鱼肌肉呈鲜甜味游离氨基酸从31.57%升高至41.87%,8月份从43.19%升高至46.45%,11月份从43.81%升高至46.50%,3月份从30.40%升高至31.50%。
暂养过程中不同氨基酸变化不同,其中,缬氨酸、异亮氨酸、亮氨酸和组氨酸含量降低,丝氨酸、谷氨酸、丙氨酸、酪氨酸、赖氨酸和苯丙氨酸含量升高,苏氨酸、脯氨酸、甘氨酸、胱氨酸、蛋氨酸和精氨酸在不同月份变化趋势不同。
2.6 暂养对鳙鱼肌肉呈味核苷酸含量的影响
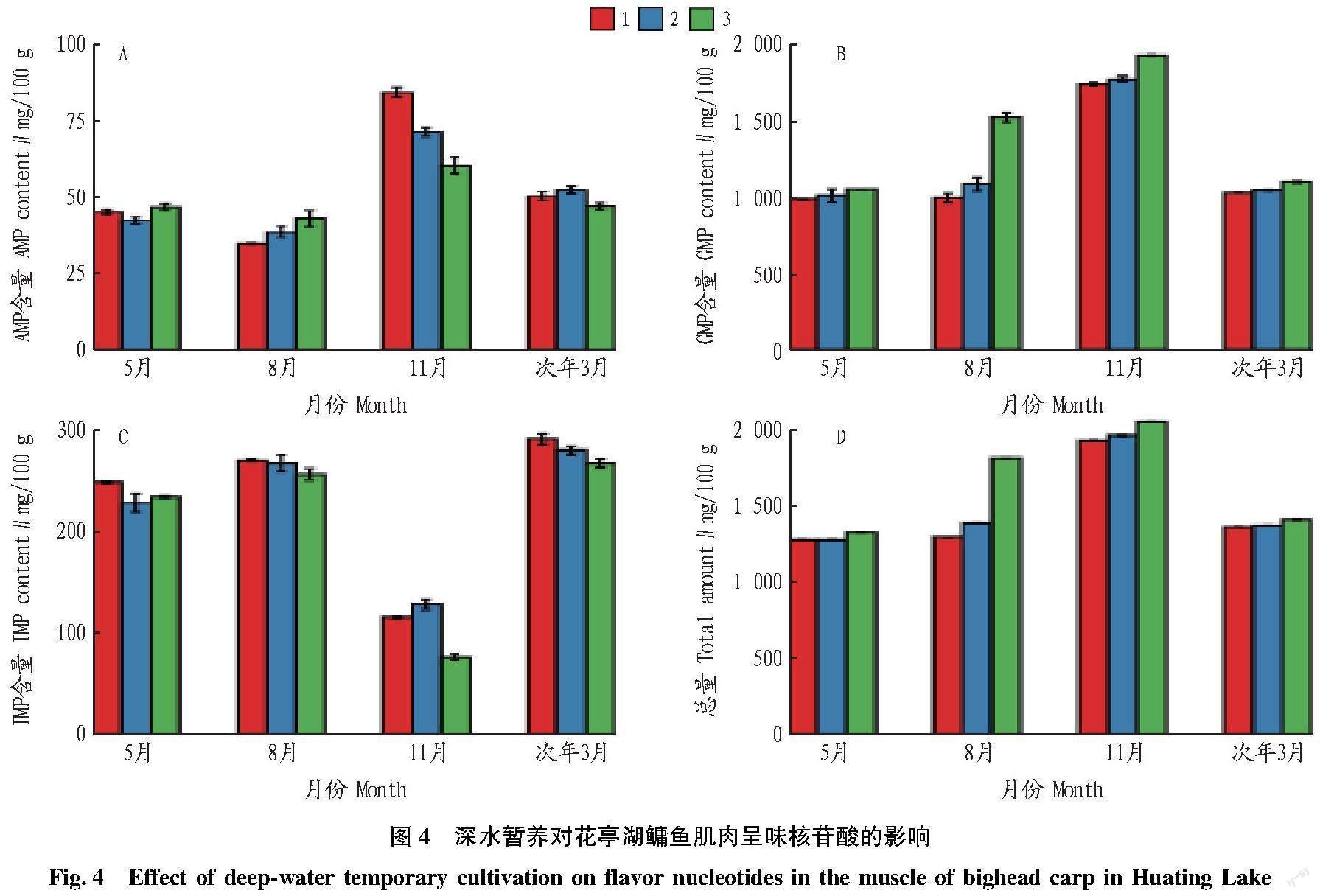
不同月份暂养对鳙鱼肌肉呈味核苷酸的影响如图4所示。不同月份鳙鱼肌肉IMP、GMP和AMP含量差异显著,导致呈味核苷酸总量差异显著。暂养期间,鳙鱼肌肉IMP、GMP和AMP含量会有波动,但是呈味核苷酸总量随着暂养时间的延长而升高,其中5月份呈味核苷酸总量从1 278.2 mg/100 g升高至1 330.8 mg/100 g,8月份从1 297.8升高至1 820.0 mg/100 g,11月份从1 934.2 mg/100 g升高至2059.4 mg/100 g,次年3月份从1 368.4 mg/100 g升高至1 413.1 mg/100 g。
3 讨论
3.1 暂养对鳙鱼形态的影响
形态学指标是可以反映鱼類在食物匮乏或停食情况下其自身营养储备消耗情况的直观指标[7-8]。该研究将传统形态学分析与框架分析相结合对鳙鱼暂养过程中形态变化进行研究。基于对鳙鱼形态学的观测,发现与鳙鱼腹部密切相关的框架参数如D5/L1、D7/L1、D8/L1、D11/L1、D12/L1和D15/L1随暂养时间的延长而减小,说明暂养对鳙鱼有“减肥”作用。以往关于鱼类暂养的形态学研究主要基于肥满度、脏体比等指标进行观测[2,7-8],需解剖鱼类才能获得相关指标,耗时耗力。该研究使用非解剖方法的形态学指标可以科学衡量鳙鱼“瘦身”效果,操作更简单,成本更低廉,可推广于企业,为企业暂养提供可操作性更强的技术支撑。
3.2 暂养对鳙鱼免疫和抗氧化能力的影响
天然水体中长成的鳙鱼,在大水面捕捞、运输至暂养设施的过程中,会产生应激反应,机体的一些免疫和应激反应指标会随之发生变化。MDA是反映鱼体的氧化损伤程度指标之一。该研究中,不同月份鳙鱼暂养初期肌肉MDA含量最高,可能是鳙鱼暂养初期,捕捞时高密度暂养、食物缺乏和运动受限导致鳙鱼应激反应加重,但随着暂养时间的延长,应激反应得到缓解[9]。该研究中,暂养初期,SOD和CAT含量最高,与宋志明等[10]的研究结果一致。在正常生理条件下,鱼体内SOD和CAT等多种抗氧化酶,可清除鱼体内代谢产生的活性氧[11]。在清除活性氧过程中,SOD产生的H2O2在CAT的催化作用下,可生成H2O和O2,从而对机体起保护作用[12]。TP在免疫和供能等方面发挥重要作用[13],可用于衡量鱼类肝损伤程度[14]。该试验中,暂养初期TP含量最高,可能是由于在暂养初期鳙鱼的应激反应使其肝损伤比较严重,从而导致TP浓度含量升高,黑鲷(Sparus inacrocephalus)[15]和半滑舌鳎(Cynoglossus semilaevis)[14]在胁迫后,肌肉内TP含量也呈现着升高的趋势。暂养后期TP含量降低,与赛娜等[16]的研究结果一致,说明随着暂养时间的延长,鳙鱼机体处于免疫应激状态,血液中TP浓度会逐渐下降。
总之,暂养时间的适当延长对鳙鱼的抗氧化能力及免疫机能具有改善作用。捕捞胁迫会使鳙鱼产生严重的应激反应,若立刻上市销售会造成鳙鱼的大量死亡,而暂养可以消除捕捞时产生的环境胁迫,提高鳙鱼的免疫机能,暂养后的鳙鱼耐运输,适宜专卖店、酒店等消费前的二次暂养。
3.3 暂养对鳙鱼基本营养成分和肌肉口感的影响
面对饥饿胁迫时,鱼类会动用机体营养物质来维持生命活动,在饥饿过程中,糖类、脂肪和蛋白质将会不同程度地被消耗[17]。该研究中,随着暂养时间的延长鳙鱼蛋白质和脂肪含量的降低,水分和灰分含量上升,与吴朝朝等[18]和钟金香等[2]的研究结果一致。暂养后,鳙鱼肌肉水分含量升高,一方面是随着鱼体营养物质的不断消耗,水分含量相对上升;另一方面,鱼类少食或停食,使得它们代谢的一些内源性物质被水取代[19]。以往的研究表明,鳙鱼在0~20 d的暂养期内蛋白质分解代谢程度高[20],在此过程中蛋白质分解会产生亲水力强的多肽和肽段,导致鱼肉持水性增强,肉质嫩度提升[21]。该研究亦发现随着暂养的加长,鳙鱼鱼肉蛋白质含量呈现降低的趋势,由此推测适当的暂养可以提升鳙鱼的肉质品质。
3.4 暂养对鳙鱼脂肪酸的影响
随着深水暂养时间的延长,鳙鱼会消耗体内的脂肪提供机体能量,其肌肉中不同种类脂肪酸在暂养过程中损失速率不同。在暂养过程中,鳙鱼肌肉不饱和脂肪酸(UFA)具有升高的趋势,草鱼暂养过程中也出现UFA比例增加现象[22]。鳙鱼暂养处理后,肌肉SFA比例降低,MUFA和PUFA比例升高。以往的研究已经发现,SFA过高则会导致机体肥胖,引起心血管疾病等[23-24]。PUFA可降低脂质在各器官的蓄积[9],促进机体生长发育及抵抗疾病。MUFA具有抗炎、降低心血管疾病等功效[25]。花亭湖鳙鱼暂养15 d左右后其肉质具有更高比例的MUFA和PUFA,更低比例的SFA,表明深水暂养15 d左右对于鳙鱼暂养是适合的,以往有研究认为循环水系统暂养20~25 d是较为理想的处理周期[2],推测鳙鱼在不同养殖环境对暂养的耐受不同,后续需要为鳙鱼暂养提供更为多样化的数据支撑。总体而言,深水暂养处理后的鳙鱼具有更高的营养价值。
3.5 暂养对鳙鱼风味的影响
湖泊深水暂养过程中鳙鱼肌肉中组氨酸含量降低,循环水养殖车间的鳙鱼在饥饿处理后,其肌肉中组氨酸含量也降低[2]。其他鱼类,如鮸鱼(Miichthys miiuy)[26],遮目鱼(Chanos chanos)[27],异育银鲫(Carassiusauratus gibelio)[28]和草鱼(Ctenopharyngodon idella)[29],在饥饿胁迫后体内组氨酸含量也呈现降低的趋势。这可能是由于组氨酸是生糖氨基酸,在饥饿期间可以转化为糖原为鱼类提供能量[30],是首先被鱼类机体消耗的氨基酸种类之一[28]。故随着饥饿时间的延长,会导致肌肉组氨酸含量的持续下降。值得注意的是,组氨酸在组氨酸脱氢酶的催化下可以形成组胺,组胺中毒是水产品存在的安全问题[31],上市前的暂养处理使鳙鱼肌肉组氨酸含量显著降低,有助于降低组胺产生的风险。在该研究中,深水暂养使鳙鱼肌肉中丝氨酸、丙氨酸和苯丙氨酸含量升高,其他鱼类在饥饿过程中,亦会有部分氨基酸含量升高,如异育银鲫在饥饿过程中,肌肉中异亮氨酸、谷氨酸和苏氨酸等会升高[28],推测饥饿状态下不同氨基酸会发挥特定的生理功能,所以具有不同的變化趋势[30],如经过饥饿胁迫后,鱼体肌肉分解加强会导致大部分的氨基酸转化为丙氨酸和谷氨酰胺,故引起鳙鱼肌肉中谷氨酸含量的升高。谷氨酸是多种氨基酸进入能量代谢循环的中间产物,所以其含量升高显著。暂养后呈鲜甜味氨基酸总量及比例皆有升高,则主要得益于甜味氨基酸中谷氨酸、甘氨酸和丙氨酸含量的升高,以往关于草鱼的瘦身养殖研究亦有类似的结论[32]。呈味核苷酸是水产品中重要的呈味成分和助味成分,如AMP能加强鱼肉的滋味[6],ATP的分解产物与鱼肉的风味密切相关[9]。该试验中,随着暂养时间的延长,呈味核苷酸总量增加可能是因为能量消耗牵扯到ATP的降解[33],使其中间产物总量含量升高。
4 结论
深水湖泊暂养后的鳙鱼“瘦身”效果明显,暂养对鳙鱼的抗氧化能力及免疫机能具有改善作用。暂养使鳙鱼肌肉持水性更强,增加肉类的嫩度;甜味氨基酸比例和呈味氨基酸总量均增加。总之,深水暂养鳙鱼为消费者提供了更为优质健康的食材,并且15 d的暂养对鳙鱼品质提升而言是较为适宜的。该研究可为深水湖泊暂养鳙鱼推广提供数据支持。
参考文献
[1]薛勇,王超,于刚,等.鳙鱼肉中土腥味物质的测定方法[J].中国水产科学,2010,17(5):1094-1100.
[2] 钟金香,李俊伟,颉晓勇,等.饥饿处理对鳙鱼形态和肌肉营养成分的影响分析[J].基因组学与应用生物学,2019,38(8):3515-3520.
[3] 李武辉,胡婕,孙成飞,等.大口黑鲈 (♀)× 蓝鳃太阳鱼 (♂) 杂交F1的形态及遗传特征[J].水产学报,2020,44(8):1225-1236.
[4] 陈小雷,胡王,马朝彬,等.丁香油对蟹黄鲜虾酱贮藏品质的影响[J].渔业科学进展,2020,41(6):181-191.
[5] BOOKSTEIN F L.Foundations of morphometrics[J].Annu Rev Ecol Syst,1982,13:451-470.
[6] 陈剑岚.草鱼的生理差异及保藏温度对肌肉呈味成分的影响[D].上海:上海海洋大学,2017:11-12.
[7] 朱站英,华雪铭,于宁,等.草鱼蛋白质和脂肪代谢对饥饿胁迫的响应[J].水产学报,2012,36(5):756-763.
[8] 钟金香,李俊伟,颉晓勇,等.短期饥饿对斑点叉尾鮰形态、肌肉品质构成及营养组成的影响[J].南方水产科学,2018,14(2):90-95.
[9] BARRENTO S,POWELL A.The effect of transportation and re-watering strategies on the survival,physiology and batch weight of the blue mussel,Mytilus edulis[J].Aquaculture,2016,450:194-198.
[10] 宋志明,刘鉴毅,庄平,等.低温胁迫对点篮子鱼幼鱼肝脏抗氧化酶活性及丙二醛含量的影响[J].海洋渔业,2015,37(2):142-150.
[11] WINSTON G W.Oxidants and antioxidants in aquatic animals[J].Comp Biochem Physiol C,1991,100(1/2):173-176.
[12] 胡静,吴开畅,叶乐,等.急性盐度胁迫对克氏双锯鱼幼鱼过氧化氢酶的影响[J].南方水产科学,2015,11(6):73-78.
[13] 周玉,郭文场,杨振国,等.鱼类血液学指标研究的进展[J].上海水产大学学报,2001,10(2):163-165.
[14] 孙学亮,邢克智,陈成勋,等.急性温度胁迫对半滑舌鳎血液指标的影响[J].水产科学,2010,29(7):387-392.
[15] 李丹丹,陈丕茂,朱爱意,等.塑料袋密封充氧运输密度对增殖放流黑鲷成活率的影响[J].南方水产科学,2018,14(5):36-44.
[16] 赛娜,宋凯.运输胁迫对牙鲆血液生化指标的影响[J].黑龙江畜牧兽医,2011(13):151-152.
[17] GAYLORD T G,GATLIN D M Ⅲ.Dietary protein and energy modifications to maximize compensatory growth of channel catfish (Ictalurus punctatus)[J].Aquaculture,2001,194(3/4):337-348.
[18] 吴朝朝,陈丽丽,袁美兰,等.短期暂养对鲫鱼肉品质影响的研究[J].食品工业科技,2015,36(15):334-337.
[19] 胡伟华,吕昊,樊启学,等.净化时间对微流水系统中鳙品质影响的研究[J].水生生物学报,2019,43(5):1056-1061.
[20] GASPAR A L C,DE GES-FAVONI S P.Action of microbial transglutaminase (MTGase) in the modification of food proteins:A review[J].Food Chem,2015,171:315-322.
[21] PALMERI G,TURCHINI G M,MARRIOTT P J,et al.Biometric,nutritional and sensory characteristic modifications in farmed Murray cod (Maccullochella peelii peelii) during the purging process[J].Aquaculture,2009,287(3/4):354-360.
[22] ABI-AYAD S M E A,BOUTIBA Z,MLARD C,et al.Dynamics of total body fatty acids during early ontogeny of pikeperch (Sander lucioperca) larvae[J].Fish Physiol Biochem,2004,30(2):129-136.
[23] 張宁宁,王真真,陈乃宏.膳食脂肪酸与抑郁症的相关性及机制研究进展[J].药学学报,2021,56(4):906-912.
[24] 陈镜宇,范建高.代谢相关脂肪性肝病的危害和饮食治疗对策[J].肝脏,2021,26(11):1287-1289.
[25] 付国玲.单不饱和脂肪酸在铁死亡中的作用以及机制的研究[D].无锡:江南大学,2018:5-6.
[26] 柳敏海,罗海忠,傅荣兵,等.短期饥饿胁迫对鲬免鱼生化组成、脂肪酸和氨基酸组成的影响[J].水生生物学报,2009,33(2):230-235.
[27] SHIAU C Y,PONG Y P,CHIOU T K,et al.Effect of starvation on free histidine and amino acids in white muscle of milkfish Chanos chanos[J].Comp Biochem Physiol B Biochem Mol Biol,2001,128(3):501-506.
[28] HE W P,LI P P,YAN H G,et al.Long-term fasting leads to preferential catabolism of His,Arg,and branched-chain amino acids in the dorsal muscle of gibel carp (Carassius auratus gibelio):Potential preferential use of amino acids as energy substrates[J].Aquaculture,2022,552:1-11.
[29] 游文章,文华,马琳.饥饿对草鱼血清游离氨基酸的影响[J].淡水渔业,2007,37(3):26-29.
[30] HALVER J E,HARDY R W.Fish nutrition[M].Third Edition.San Diego:Academic Press,2003:309-365.
[31] 谢超,王阳光,邓尚贵.水产品中组胺产生机制及影响因素研究概述[J].肉类研究,2009,23(4):74-78.
[32] 唐洪玉,魏士卜,刘长江,等.瘦身养殖对草鱼形态和肌肉营养成分的影响[J].重庆水产,2021(3):32-40,50.
[33] TRBOVIC′ D,MILOJKOVIC′-OPSENICA D,PETRONIJEVIC′ R,et al.Influence of diet on proximate composition and fatty acid profile in common carp (Cyprinus carpio)[J].J Food Compos Anal,2013,31(1):75-81.
猜你喜欢
家教世界(2021年4期)2021-03-09
学生天地(2019年31期)2019-08-25
湖北农业科学(2019年24期)2019-02-06
数位时尚(幼儿教育)(2018年3期)2018-04-12
云南中医中药杂志(2016年9期)2016-11-29
中外医学研究(2016年28期)2016-11-28
人间(2016年24期)2016-11-23
垂钓(2016年9期)2016-10-31
爱尚生活(2016年9期)2016-10-21
垂钓(2016年8期)2016-09-09
