
大豆肽不溶性聚集体形成机制及应用研究进展*
2024-03-06 10:43江连洲苏睿涵张雪晴冯玉田
大豆科技 2024年1期
江连洲,苏睿涵,张雪晴,冯玉田,孙 娜
(东北农业大学食品学院,哈尔滨 150030)
蛋白质在酶解过程中会出现凝胶状物质或沉淀[1-2],通过对这种物质的成分及形成机制研究发现疏水作用是不溶性聚集体形成的主要机制,但也不排除少量其他作用(如转肽作用、离子键等)的存在。这种形成不溶性聚集体的反应可引起肽结构改变,改善肽的生物活性[3]。产生的不溶性聚集体被认为是酶水解过程中的副产物,可降低蛋白质水解效率。这种不溶性聚集体中氨基酸含量丰富,是一种优质的蛋白质资源。ZHANG 等[4]将大米蛋白水解产生的不溶性聚集体进行超声处理,制备出具有抗氧化性的Pickering 乳液。DU 等[5]用酶解羊奶酪蛋白所产生的不溶性聚集体制备纳米颗粒用于运载姜黄素。
大豆肽是大豆蛋白水解产生的多种肽的混合物,氨基酸组成与大豆蛋白几乎相同,但大豆肽的致敏性更低,且较大豆蛋白更易被消化吸收。同时大豆肽还具有抗氧化、抗衰老、平衡肠道菌群、降低胆固醇、改善新陈代谢和提升免疫力等生理活性[6],对人体健康具有重要作用。近年来,国内外学者对大豆肽不溶性聚集体的功能性质及其在食品中的应用展开了深入研究。本文综述了大豆肽不溶性聚集体的形成机制、影响因素、功能特性及在食品领域中的应用,为肽不溶性聚集体的深入研究及在食品领域中的应用拓展提供了理论基础。
1 不溶性聚集体形成机制
在蛋白酶催化蛋白质水解成小分子肽或游离氨基酸的过程中会产生溶解性较差的聚集体(一种类蛋白凝胶状物质)。1974 年,ASO 等[7]提出疏水作用是影响不溶性聚集体形成的重要因素。1976年,HOFSTEN 等[8]提出plastein 反应是一种重排过程,小分子肽通过非共价键连接,转肽和水解所起到的作用较缩合更重要。GONCA 等[9]借助实验证实plastein 反应只涉及疏水作用和离子键机制,未检测到肽键的形成和转肽作用。因此,plastein反应过程中可能存在疏水、转肽、水解和离子键等多种作用,其具体形成机制仍有待深入研究。目前疏水作用被认为是不溶性聚集体形成的主要机制。
不溶性聚集体的形成过程可分为3 个步骤,①蛋白酶水解蛋白质形成不同分子质量的短肽和游离氨基酸[10];②通过蒸发、冷冻干燥或喷雾干燥浓缩水解产物;③在高底物浓度(30%~50%)下采用相同或不同的酶孵育水解产物[2](见图1)。LIU等[11]通过对乳清蛋白水解进行研究提出不溶性聚集体的形成分为2 个步骤,①在0.5 h 内形成具有疏水核心的蛋白结构;②这些结构聚集在一起,较小的结构也参与聚合。

图1 不溶性聚集体形成机制
研究发现,不溶性聚集体的产生与疏水相互作用及离子键有关。
2 影响不溶性聚集体形成的因素
2.1 物理及化学因素
蛋白质水解过程中,pH、温度及其他物质的添加等理化因素通过影响酶活性或多肽结构对不溶性聚集体的形成产生影响。由于蛋白酶种类不同,能使聚集体达到最高产率的pH 也不同(3.0-11.0)[9]。碱性环境下,大豆肽不溶性聚集体的产率降低[12],酸性环境是否影响其产率尚待研究。此外,温度也是重要影响因素。ZHAO 等[13]研究发现反应温度越高,不溶性聚集体生成速率越快,但产率会随之降低。研究表明,不溶性聚集体的形成除受酶活性影响外,还与其自身表面疏水性及其他物质(如多肽、多糖、盐、脂类等)的添加等因素有关[14]。
2.2 酶的种类
蛋白质水解程度随酶的种类不同各异,对不溶性聚集体的产生影响较大。研究表明含有疏水基团的蛋白酶更易诱导聚集体形成。LI 等[15]研究发现Alcalase 碱性蛋白酶较木瓜蛋白酶更易促进不溶性聚集体的形成。赵明等[16]研究发现,在相同处理条件下,风味蛋白酶较碱性蛋白酶和中性蛋白酶产生更多的不溶性聚集体。也有学者指出胰蛋白酶不能使大豆肽产生聚集体[17]。
2.3 底物
蛋白质水解过程中,底物种类和浓度不同可导致聚集体产率存在差异。赵明等[16]采用相同酶分别水解β-伴大豆球蛋白(7S)和大豆球蛋白(11S)时,相应聚集体产率存在差异。SUKAN 等[18]采用胃蛋白酶水解真菌蛋白和酪蛋白,其中水解酪蛋白得到的不溶性聚集体产率较真菌蛋白高14%。SHUN 等[19]研究发现,随着底物浓度增加,不溶性聚集体产率呈指数增长。因此,可以推测不同底物种类和浓度是影响不溶性聚集体产率的主要因素,且这种影响可能与底物中疏水性氨基酸含量有关。
3 大豆肽不溶性聚集体营养价值及功能特性
大豆蛋白是一种营养价值丰富且具有良好功能特性的蛋白质,其被不完全水解后产生的大豆肽作为功能性肽被广泛应用于食品领域。大豆蛋白在水解过程中由于疏水作用发生聚集,产生大豆肽不溶性聚集体。大豆肽不溶性聚集体的扫描电子显微镜图像[16]如图2 所示。蛋白形成聚集体后,其组成及结构产生变化,导致其营养价值及功能特性发生改变。

图2 大豆肽不溶性聚集体扫描电子显微镜图像
3.1 营养价值
与大豆分离蛋白(SPI)相较,大豆肽不溶性聚集体中的Thr、Val、Ala、Leu、Ile、Tyr、Phe、Pro、Met 和Cys 比例显著升高(p <0.05),Glu 和Asp 比例显著降低,其他氨基酸如Ser、His、Lys和Arg 的含量无显著差异(p >0.05)[20]。Val、Ala、Leu、Ile、Phe、Tyr 和Pro 均为疏水性氨基酸,因此大豆肽不溶性聚集体中的疏水性氨基酸含量较高,其占比超过40%。此外,大豆肽不溶性聚集体中的必需氨基酸含量也高于SPI,其占比达38%。进一步表明不溶性聚集体的形成是由于蛋白质被酶水解时氨基酸间发生疏水相互作用,疏水性氨基酸聚集形成凝胶或沉淀。所形成的大豆肽不溶性聚集体中必需氨基酸含量丰富,具有较高营养价值。
3.2 功能特性
UDENIGWE 等[21]研究发现,与SPI 相较,大豆肽不溶性聚集体表现出较好的抗胃消化特性和良好的抗氧化活性,但在肠道中易被消化,可作为姜黄素等生物活性物质的递送载体[22]。此外,大豆肽不溶性聚集体还具有良好的盐离子稳定性[23]。研究发现,物理化学改性会对大豆肽不溶性聚集体的结构及功能特性产生影响,进而拓宽其在食品领域中的应用。赵明[16]采用均质和pH 偏移方法处理不溶性聚集体,使其溶解度提升,改性后的不溶性聚集体的乳化性优于SPI,且经pH 偏移处理的不溶性聚集体所制备的乳液具有良好的贮藏稳定性[24]。经超声处理后的大豆肽不溶性聚集体的抗氧化能力良好,与表面活性剂HPMC/Tween 80 混合后提升了乳化能力,可作为一种双功能乳化剂[25]。
4 大豆肽不溶性聚集体在食品中的应用
4.1 营养应用
大豆肽不溶性聚集体中含有大量必需氨基酸,营养价值丰富。此外,还可通过掺入所需氨基酸的方式赋予其特定营养,提高大豆肽不溶性聚集体营养价值(见图3(a))。如在大豆肽不溶性聚集体中添加蛋氨酸用作虾饲料可有效促进幼虾生长[26]。也可通过添加半胱氨酸提升大豆肽不溶性聚集体营养价值,合成理想蛋白质[27]。与游离氨基酸相较,将所需氨基酸掺入肽聚集体中更易被人体吸收利用。氨基酸掺入的方法还可提升蛋白水解物的抗炎活性[28]。但由于该种方法成本较高,且产物的营养价值和功能特性具有可替代性,因此未被广泛应用。

图3 大豆肽不溶性聚集体特定氨基酸掺入反应机制及脱苦机制
4.2 脱苦应用
大豆蛋白的水解物通常含有苦味,限制了其在食品领域中的应用。产生的苦味与其氨基酸序列有关。目前采用物理、化学及生物方法在蛋白质水解时可抑制苦味肽生成,减少蛋白质水解产物苦味。生物脱苦可在保证脱苦效果的同时降低脱苦成本。在酶水解过程中,蛋白质结构展开,内部疏水基团暴露。LIAO 等[29]研究表明肽中疏水基团种类和数量可影响苦味阈值,具有高疏水性侧链基团的氨基酸包括Trp、Tyr、Phe、Leu、Ile、Val、Pro。ISHIBASHI 等[30]研究证实,增加Phe 或Tyr 比例可引起苦味强度提升,苦味与肽疏水性及疏水性氨基酸含量呈正相关[31]。大豆肽不溶性聚集体中疏水性氨基酸占比为40%~45%。如图3(b)所示[32],在蛋白酶水解过程中氨基酸间由于疏水作用发生聚集,形成将疏水性氨基酸包裹在内的聚集体,减小味觉对苦味的感知,这就是不溶性聚集体的形成可用来脱苦的原因[33]。
4.3 活性物质递送及其他应用
不溶性聚集体是大豆肽生产过程中的副产物。ZHANG 等[22]研究发现这种不溶性聚集体含有较高比例的必需氨基酸,且在消化过程中可释放小肽和氨基酸,表现出良好的抗氧化活性。为了避免资源浪费,如何有效利用不溶性聚集体成为新的研究方向。通过理化手段将不溶性聚集体进行改性可显著提升其溶解度、乳化性、贮藏稳定性及抗氧化能力等[9,23-24]。
由图4 可知,改性后的大豆肽不溶性聚集体可用于活性物质的递送。ZHANG 等[22]研究发现经pH 偏移处理的不溶性聚集体在递送姜黄素时表现出良好的模拟胃肠消化稳定性、抗氧化活性和体外生物利用度。大豆肽不溶性聚集体也可与其他物质结合。DONG 等[34]将不溶性聚集体与EGCG 结合,制备的乳液具有更好的稳定性。JING 等[35]研究发现经碱性处理后的大豆肽不溶性聚集体可用于制作蛋黄酱,表现出良好的感官特性和稳定性,可作为绿色食品代替传统蛋黄酱。改性后的大豆肽不溶性聚集体的稳定性和乳化性提升,并可作为活性物质的优质载体,在食品领域中具有良好的应用价值。
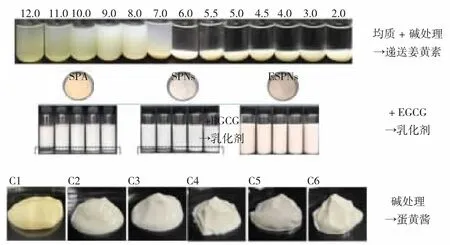
图4 大豆肽不溶性聚集体的改性及应用
5 总结与展望
大豆肽不溶性聚集体是酶解过程中的副产物,其形成机制仍有待深入研究。目前已知其主要聚集原因为疏水作用,也包含转肽、离子键等其他因素作用。大豆肽不溶性聚集体含有丰富的必需氨基酸,具有良好的抗氧化活性和抗炎活性,具有一定的营养价值和功能特性,在食品领域中应用前景广阔。目前大豆肽不溶性聚集体主要用于解决生物活性肽的苦味问题及提升营养价值。此外,由其制备的水凝胶和纳米颗粒还可用于生物活性物质运输。大豆肽不溶性聚集体的有效利用减少了资源浪费,为绿色生产及食品工业中副产物的有效利用奠定基础。
猜你喜欢
合成化学(2024年3期)2024-03-23
合成化学(2023年12期)2024-01-02
职工法律天地·上半月(2022年3期)2022-04-15
河南工业大学学报(自然科学版)(2021年6期)2022-01-26
老年博览·上半月(2019年8期)2019-10-21
猪业科学(2018年8期)2018-09-28
中成药(2018年8期)2018-08-29
中成药(2018年7期)2018-08-04
儿童故事画报·智力大王(2017年2期)2017-06-05
中学生数理化·中考版(2015年12期)2015-09-10
