
益生菌缓解金黄色葡萄球菌感染牛乳腺炎的研究与应用进展
2024-02-21 11:17朱妍刘慧燕闫丹丽赵春光滕政蓉张英刘晓燕方海田武俊瑞
中国乳品工业 2024年1期
朱妍,刘慧燕,闫丹丽,赵春光,滕政蓉,张英,刘晓燕,方海田,武俊瑞*
(1.沈阳农业大学食品学院辽宁省食品发酵技术工程研究中心沈阳市微生物发酵技术创新重点实验室,沈阳 110866;2.宁夏食品微生物应用技术与安全控制重点实验室宁夏大学食品与葡萄酒学院,银川 750021;3.宁夏伊品生物科技股份有限公司,银川 750001)
0 引 言
奶牛乳腺炎根据乳房以及乳汁的临床表现,可分为临床乳房炎(Clinical mastitis,CM)和隐性乳腺炎(Subclinical mastitis,SCM)。SCM 又称亚临床型乳腺炎,其特征是肉眼观察奶牛乳房外表和乳汁颜色等均不见病理变化和临床病变,通过平常的临床检查方法较难发现[1]。据统计在我国SCM 发病率占奶牛乳腺炎总发病率的77%~79%[2]。导致生产成本和治疗费用增加,牛奶品质及牛场收入下降。其中金黄色葡萄球菌(Staphylococcus aureus,S. aureus)是感染的主要病原体[3],可侵入上皮细胞,从而逃避免疫防御并持续感染。使用抗生素治疗是首选方法,但随着多药耐药病原体的出现以及牛奶和奶制品中抗生素残留的问题开始显现[4]。抗生素不再被认为是治疗由S. aureus引起的牛乳腺炎的最佳选择,而益生菌是存在于体内的天然微生物,性质稳定且对机体起有益作用[5],有研究使用鼠李糖乳杆菌(Lactobacillus rhamnosus)、乳酸菌(L.casei)及其分泌物干预乳腺上皮细胞炎症模型,具有良好抗炎效果[6],因此用益生菌替代抗生素治疗是新的治疗方法[7]。本文概述有关S. aureus感染牛乳腺炎的发病机制,从基因和蛋白方面论述S. aureus侵染牛乳腺的过程的3 个阶段,总结目前益生菌代替抗生素治疗牛乳腺炎的研究进展,阐明益生菌预防或治疗牛乳腺炎的过程。
1 SA 感染牛乳腺炎的发病机制
S. aureus感染牛乳腺分为3 个阶段,首先S. aureus黏附乳头皮肤和皮下基质,其表面存在各种结合蛋白和聚集因子,等使其定值于乳腺上皮细胞(bovine Mammary Epithelial Cells,bMEC)并在整合素下完成入侵,释放细胞表面黏附因子、毒素、凝固酶等毒力因子,其表达受群体感应系统Agr,SarA 蛋白家族以及SaeRs二次元信号系统等局限性调控因子的协同调控,干扰机体免疫系统[8]。第二阶段侵入、破坏乳腺组织和细胞,S. aureus入侵时bMEC 和巨噬细胞通过TLR-2 受体识别到并做出免疫应答[9]。但S. aureus会在多个细胞层中积累,形成尽可能多的细菌聚集物形成生物膜逃避免疫系统并大量增殖,如图1 所示。因此导致牛奶产量与品质急速下降的原因是S. aureus损害bMEC 和腺泡的功能。
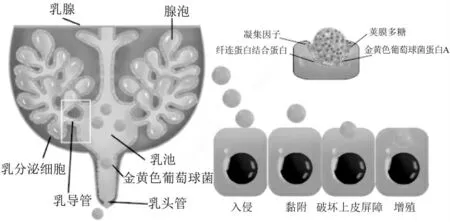
图1 金黄色葡萄球菌入侵牛乳腺上皮细胞侵染过程
1.1 免疫逃避机制
S. aureus黏附聚集到一定程度形成生物膜,保护细菌逃避免疫系统的攻击和抗菌药物的伤害[10]。生物膜的成熟和迁移阶段受Agr基因的调控[11],是重要的毒力调控因子控制hla及psm的表达,该基因由RNAIII(一种调节性mRNA 分子)激活肽的靶点构成,其增加会诱导生物膜生长。psm基因调控酚溶性调控蛋白(Phenolsoluble modulin,PSMs)的表达[12],有利于生物膜解离、播散及内部微孔通道形成,让深层细菌吸取营养。细菌之间的聚集受PIA 蛋白(多糖胞间黏附素由线性β-1,6-糖苷键连接氨基葡萄糖聚糖组成)的调节[13],其合成由细胞间黏附(icaA、icaD、icaBC)位点介导。一旦生物膜完全成熟,它就会分散并释放固着细胞,这些细胞准备重新填充相同的位点(初级位点)或一个新的位点。据统计,在S. aureus致奶牛乳腺炎过程中至少有25 种毒素,15 种黏附蛋白,20 种免疫逃避分子发挥重要作用[14]。通过生成细胞膜、分泌超抗原样蛋白-3 以及分泌一系列的毒力因子如溶血素,黏附素等来减弱固有免疫系统的应答,引发持续性感染。
1.2 破坏免疫屏障
S. aureus菌株黏附上皮形成荚膜多糖(Capsular polysaccharide,CP)阻挡中性粒子识别,阻拦巨噬细胞吞噬,其识别病原体主要依赖模式识别受体( patternrecognition receptors,PRR),而Toll 样受体 (Toll like receptor,TLR) 和NOD 样受体 (nucleotide-binding oligomerization domain ( NOD ) like receptors,NLR)最为庞大。研究发现S. aureus 通过NOD1/NOD2 识别激活NF-κB(nuclear factor Kappa-B)信号通路,负责促炎细胞因子(尤其是IL-6 和TNF-α)的转录,通过TLR2干扰细菌识别或掩蔽TLR2-active 脂蛋白促进CP 的表达[15]。以小鼠建立的乳腺炎模型证明脂多糖(lipopolysaccharide,LPS)能够通过调控乳腺细胞紧密连接蛋白的表达量来破坏血液与乳汁之间的屏障[16]。通过下调奶牛乳腺上皮细胞E 钙黏蛋白(E-cadherin)的表达量导致细胞黏附连接的破坏,从而破坏血液与乳汁之间的屏障[17]。S.aureus通过抑制MTCRCI 和JAK/STAT 通路的活性进而降低氨基酸转运蛋白基因的表达,入侵MCF-10A 的能力并在细胞内增殖,抑制MCF-10A 对外源谷氨酸(Glu)、天冬氨酸(Asp)和亮氨酸(Leu)的吸收,抑制乳蛋白合成和分泌[18]。在S. aureus感染早期的致病因子中黏附素最为重要,只有当S. aureus黏附到宿主细胞后才能释放毒素和荚膜[19]。参与金黄色葡萄球菌侵染机体的相关基因与蛋白如表1 所示。

表1 参与金黄色葡萄球菌侵染的相关基因与蛋白
除上述相关蛋白外,还有一部分蛋白通过相关基因调控生物膜中PIA 合成机制,介导生物膜形成的初始定植和黏附例如:金黄色葡萄球菌蛋白A(Staphylococcalprotein A,SPA)、生物膜相关蛋白(biofilm-associated protein,BAP)、金黄色葡萄球菌表面蛋白G(Staphylococcus aureussurface protein G ,SASG)[20]。
1.3 感染后侵袭过程
在乳腺炎发病过程中,S. aureus首先激活bMEC的TLR2 信号通路,然后进一步激活包括Caspae 在内的多种促凋亡信号,可能是通过Fas-FADD-caspase8-caspase3 通路介导启动细胞凋亡程序[21],而线粒体细胞色素C-caspase9 与caspase1-IL1β 途径可能不参与此调亡诱导过程。S. aureus可上调乳腺组织中趋化因子和补体因子基因,以及白细胞介素受体基因的mRNA 转录水平,同时下调自噬调节因子基因的mRNA 转录水平,从而应对奶牛乳腺炎症过程[22]。还有研究发现患乳腺炎的奶牛相关基因表达与DNA 甲基化水平有关[23],奶牛乳腺炎源性S.aureus可以诱导乳腺纤维化[24],促进乳腺组织中TGF-β1 的表达。TGF-β1是一种多功能的细胞因子可调节机体免疫与细胞的易感性方面,通过促进bMECs 表面Fn,β1 整合素(Integrinβ1)和细胞凋亡的膜表面分子(Fas)大量表达从而促进S. aureus 黏附与侵入bMECs[25]。在TGF-β1诱导的奶牛乳腺纤维细胞(BMFBs)成纤维化中,S. aureus通过抗α 平滑肌动蛋白(α-SMA)和I 型胶原抗体(ICollagen)增强BMFBs 的黏附与侵袭受ERK 信号调控。此外乳腺纤维化还与丝裂原激活蛋白激酶(Mitogen-activated protein kinase,MAPK) 信号通路有关,激活后的核苷酸结合寡聚化结构域蛋白(NOD2)能使其激活从而导致宿主防御及炎性细胞因子的产生。热灭活的S. aureus促进BMFBs ERK1/2、P38 及JNK 的磷酸化水平,MAPKs 特异性抑制剂显著抑制S. aureus诱导的MMP-2/TIMP-1、-2 及uPA/PAI-1的表达,进而初步说明S. aureus感染奶牛乳腺组织导致乳腺纤维化的发病过程中的相关机理。
2 益生菌对牛乳腺炎的调节机制
2.1 益生菌治疗机制研究进展
益生菌对牛乳腺炎作用机制主要通过以下3 种方式:(1)与病原菌直接相互作用或分泌抑菌物质抑制病原菌在肠道内的定植;(2)益生菌黏附在肠上皮细胞上能够促进黏蛋白、防御素的表达分泌,增强肠上皮细胞的紧密连接功能,从而提高肠上皮细胞的屏障功能;(3)益生菌到达胃肠道内能够调节机体的免疫反应,释放细胞因子,增强宿主的抵抗力,其中白细胞介素(IL)、干扰素(INFs)、肿瘤坏死因子(TNF)、转化生长因子(TGF)和免疫因子均为细胞因子。
2.1.1 益生菌与病原体直接相互作用
益生菌可抑制S. aureus产生生物膜,进而阻止其入侵。有研究发现乳酸菌可以改善皮肤健康或抵抗病原菌的入侵。MITSUWAN W 等[26]从棕榈糖中提取出的乳酸杆菌株,其益生菌上清液可抑制S. aureus生物膜形成。WALENCKA E 等[27]使用3 种嗜酸乳杆菌菌株获得的表面活性剂抑制S. aureus的沉积速率以及生物膜的发育和成熟,且不影响细胞的生长。也有发现其产生脂肪酶而抑制S. aureus生物膜的形成。毕赤酵母中含有高度特异性溶菌蛋白酶通过对其进行基因改造,获得活细胞培养物更有效破坏S. aureus生物膜[28]。
益生菌与S. aureus存在生存竞争关系,在肠道内定植后干扰其群体感应信号,消除包括S. aureus在内的其他病原体,并分泌脂磷壁酸和脂肽等具有抗菌活性的物质从而抑制生物膜的形成。研究发现植物乳杆菌K8 通过其细胞壁中所含的脂磷壁酸抑制角质形成细胞中的肿瘤坏死因子-α(Tumor necrosis factor-alpha,TNF-α)或干扰素-γ(Interferon-gamma,IFN-γ)表达。2 种嗜酸乳杆菌与S. aureus存在直接竞争关系,产生拮抗物质,抑制其生物膜形成及黏附。PELLEGINOM S 等[29]测试乳酸杆菌亚种CRL-1655及CRL-1724 其黏附较高,并产生有机酸和乳酸链球菌素等抑菌物质,与S. aureus存在竞争关系可有效预防奶牛乳腺炎[30]。罗伊氏乳杆菌通过竞争性排斥或阻断α5β1 路径抑制S. aureus对角质形成细胞的黏附[31]。双歧杆菌与过氧化氢的形成热稳定蛋白对S. aureus有抑制作用[32]。干酪乳杆菌LCR719、ATCC 334 副干酪乳杆菌亚种A20都对S. aureus表现出抗黏附和抗菌活性[33-34]。从多种来源分离出的许多乳酸杆菌和双歧杆菌菌株在体外抑制了S. aureus临床分离株的生长[35]。除上述益生菌可以通过直接细胞竞争性排斥之外,鼠李糖乳杆菌GG、弗氏丙酸杆菌、痤疮丙酸杆菌、物乳杆菌、保加利亚乳杆菌、枯草芽胞杆菌、发酵乳杆菌也可通过竞争性排斥以及酸或细菌素样抑制剂的产生来介导[36]。张岩等[37]将枯草芽胞杆菌、地衣芽胞杆菌和酿酒酵母发酵液混合制成复合益生菌制剂,因其产生细菌素对S. aureus有显著抑制作用可以用来治疗患有SCM的奶牛。综上所述,益生菌与S. aureus存在竞争关系,可分泌抑菌物质阻止其形成生物膜,为寻找更多益生菌对S. aureus感染牛乳腺炎起到调节作用,其分子机制还需要进一步深化探讨。
2.1.2 增强肠屏障功能
益生菌黏附在肠上皮细胞促进黏蛋白、防御素的表达分泌,增强肠上皮细胞功能,抑制S. aureus黏附和侵染的数量达到预防乳腺炎的目的。L.caseiBL23 在S. aureus刺激下表现出良好抗炎效果[38],其表面具有黏附因子,如纤连蛋白结合蛋白(Fibronectin-binding protein,FBPA)[39],增加益生菌对bMEC 的黏附能力,还降低bMEC 中IL-6、IL-8、IL-1α、IL-1β 及TNF-α几种促炎细胞因子的表达。通过使用乳酸菌抑制S.aureus黏附 bMEC 产生的黏附素 ClfA、EfB 及FNBPA,发现其可作为一个可持续的替代治疗或预防乳房炎的手段。研究表明选取的4 株乳杆菌皆可抑制S. aureus的生长及其黏附HeLa 细胞的能力,同时发现降低S. aureus附着于HeLa 细胞最好的方法是排除作用。王亮亮[40]从牛奶样品中分离筛选得到3 株益生特性较好的乳酸菌菌株,通过试验也印证其抗炎机理是降低细胞促炎因子TNF-α 和IL-6 的升高利用FbpA抑制S. aureus形成生物膜并减少对bMEC 的黏附和侵染。加氏乳杆菌LA806 可诱导S. aureus黏附和内化降低5 倍并减少促炎因子的表达,并且灭活的LA806 也具有屏障和免疫调节特性[41]。
益生菌通过分泌防御素增强上皮细胞,鼠李糖乳杆菌裂解物可改善重建的人表皮模型中的皮肤屏障功能,并且用作局部治疗的多菌株益生菌制剂可改善感染的慢性缺血性伤口的愈合。研究发现罗伊氏乳杆菌ATCC-55730 通过降低其IL-8 和HBD-2 的转录水平,对感染的角质形成细胞发挥抗炎作用;干酪乳杆菌不削弱防御素的诱导,如舌抗菌肽和防御素β1,甚至在S. aureus感染期间轻微增加气管抗菌肽的诱导,最后没有改变模式识别受体NOD2 的表达,维持肠道完整性。BOUCHARD S 等[41]研究表明:L. caseiCIRM-BIA 667 可以有效地降低60%~80%金黄色葡萄球菌RF122 和Newbould 305 对bMEC 的侵染率,而且不会影响bMEC 的正常形态和存活率。L.casei对bMEC 的侵染率很低,但是比金黄色葡萄球菌RF122更容易在bMEC 中存活。乳酸乳球菌CRL 1655 和唾液乳杆菌CRL 1724 通过theta 通路黏附bMEC[42]。有发现用鼠李糖乳杆菌GR-1 (LGR-1) 处理MAC-T细胞,可保护紧密连接的完整性以及被BC 破坏的小带occludens-1 (ZO-1) 和occludin 的表达[7]。此外,LGR-1 预处理降低了NOD 样受体家族成员pyrin结构域蛋白3(NLRP3),半胱天冬酶募集和激活结构域(ASC)、Caspase-1 p20、gasdermin D (GSDMD) p30、IL-18 的表达及NLRP3 炎症小体活性,并增加了脂多糖 (LPS) ATP 刺激诱导的ZO-1 和occludin 的表达。Lactobacillus rhamnosusGG 裂解物通过增加诱导角质形成细胞迁移的CXCL2 趋化因子及其受体CXCR2 的表达来加速角质形成细胞单层的再上皮化[43]。因此表明,乳酸乳球菌、干酪乳杆菌和鼠李糖乳杆菌均可用于预防奶牛乳房炎。
2.1.3 提高机体免疫力
益生菌可通过诱导不同细胞因子释放来调节黏膜免疫反应,改善肠道微环境,增强动物对乳腺炎的免疫力[44]。有研究人员制备益生菌植物提取物复合生物制剂可显著降低牛乳腺炎的乳体细胞数[45],促进有益微生物的生长,为益生菌提供营养,还可以作为辅助因子刺激免疫反应,从而显著提高了动物的机体免疫能力。乳酸菌可帮助机体有益菌群的生长并在厌氧条件下产生乳酸,形成特殊抗生素有效抑制S. aureus的生长,其中TNF-α 和INF-γ 可以通过JNK 信号通路、STAT-1 和HIF-1 转录因子促进巨噬细胞的炎症反应、B 淋巴细胞和NK 细胞的增殖,增加炎性细胞因子IL-1、IL-2、IL-5、IL-6 和TNF-α 的分泌,通过NF-κB 或组胺依赖性途径调节上皮相关T 细胞。口服乳酸菌可促进T 细胞分化,主要反映在Th1 和Th17 细胞的稳态能力中的细胞因子IFN-γ 和IL-17分泌水平上调,这增强了动物抵抗外来致病菌入侵的免疫力。将干酪乳杆菌和鼠李糖乳杆菌混合可以刺激巨噬细胞产生细胞因子(TNF-α)并调节了宿主的免疫系统。在奶牛饲料中添加酵母菌和乳酸球菌混合制成的复合微生态制剂也印证可通过服用益生菌提高奶牛机体免疫力。植物乳杆菌17-5 可通过抑制奶牛bMEC 中NF-κB 和MAPK 信号通路的激活减弱病原菌诱导的炎症反应。L.casei在繁殖过程中能分泌多种物质,包括胞外多糖(Extracellular polymeric substances,EPS)、乳酸、短链脂肪酸、一氧化氮(NO)和细菌素。徐智敏等[46]发现使用L.caseiEPS 处理巨噬细胞能降低NO 和TNF-α 的分泌。表2 列举了某些益生菌对于患牛乳腺炎机体免疫调控的具体路径。
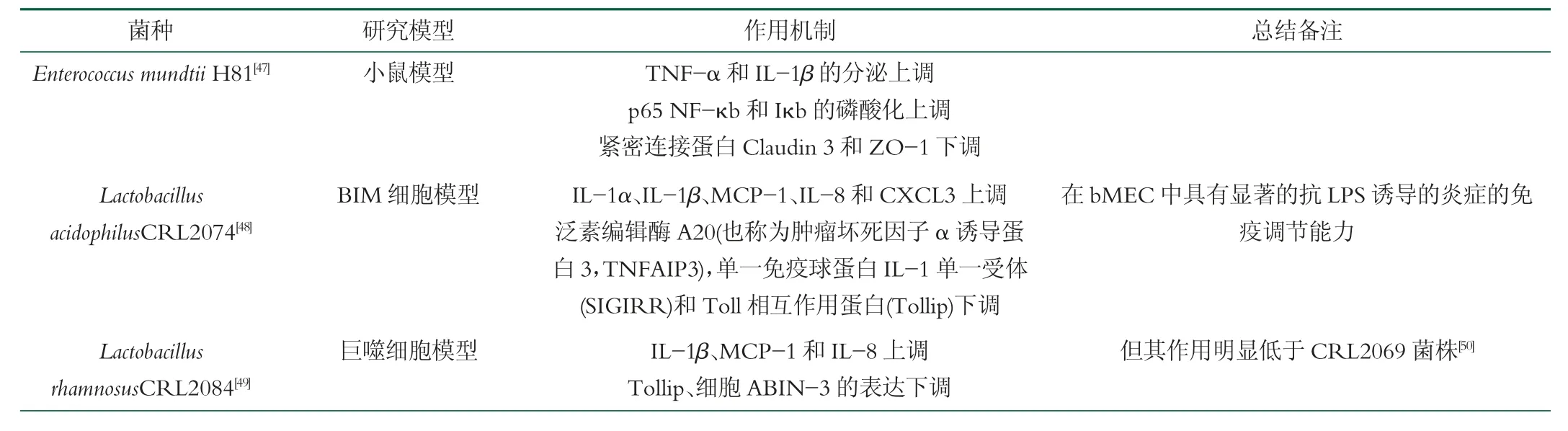
表2 益生菌调控机体免疫具体路径
2.2 益生菌预防机制研究进展
在机体感染前内服或外用益生菌可以起到预防牛乳腺炎的发生,PRINCE T 等[31]研究发现在感染S. aureus之前添加鼠李糖乳杆菌AC413 和唾液乳杆菌ucc118 可起到预防作用。有报道一种基于乳酸杆菌的新型益生菌消毒剂可用于乳房消毒,还有发现植物乳杆菌80065 和植物乳杆菌10155 混合制成的混合物可替代市面上的消毒剂[57],减少乳腺炎相关细菌,改善奶牛的微环境,提高机体免疫力。JONES M 等[58]开发了一种产生气态一氧化氮(gNO)的固定发酵L.fermentumNCIMB 7230 的贴剂装置,结果表明,将产生gNO 的益生菌贴片应用于S. aureus的培养物中,可在4~8 h 内导致细菌细胞完全死亡。奶牛口服枯草芽孢杆菌c-3102 比普通饲料效果更优患有牛乳腺炎的概率降低,因其消耗并破坏S. aureus在体内的定居。用乳酸杆菌和酿酒酵母喂养荷斯坦奶牛可以减轻牛乳腺炎的发病率和炎症的发生,还有研究发现从乳头顶端分离的细菌菌株主要是棒状杆菌属和芽孢杆菌属,可以抑制乳腺炎病原体的生长,对S. aureus起到抑制作用[59]。
3 预防与展望
奶牛乳腺炎对奶牛产生不利影响,减少产奶量,损害繁殖能力,甚至导致奶牛死亡,这些因素都对奶牛养殖养的发展产生显著的负面影响。除了外部致病菌感染之外,奶牛乳腺炎还可通过肠-乳腺途径引起[60]。研究显示益生菌具有潜在可缓解牛乳腺炎发生的作用,其副作用小,牛奶中有害物质残留量小,长期使用不会造成耐药性,是一种可以近似替代抗生素的手段。牛通过自身血液屏障保护机体免受有害菌的侵害,当细菌突破免疫系统时可通过服用益生菌增加机体T 细胞、B 淋巴细胞及NK 巨噬细胞的免疫反应,降低IL-1β、IL-6、IFN-γ、TNF 等促炎因子的表达,利用益生菌抑制S. aureus对bMEC 的黏附和侵染,降低S. aureus生物膜的形成,以减少S. aureus在乳腺内的定植,从而起到减轻乳腺感染效果。但益生菌的治疗效果有局限性,为了增加益生菌对牛机体的免疫效果有很多学者把目光转向合生元及微生态制剂方面。研究发现向4~12 周龄的犊牛饲喂添加菊粉和屎肠球菌组成的合生元饲料,犊牛腹泻比例减少,每天活体重数增加[61]。FLEIGE S 等[43]在饲喂屎肠球菌犊牛中添加乳果糖,可调节淋巴细胞含量与T 细胞亚群组成,减少致病细菌并稳定肠道环境。MIDDELBOS I S 等[62]在肠道中注入甘露聚糖,其注入部分感染负荷降低。还有研究采用喷雾干燥( spray-drying,SD )和喷雾冷冻干燥(spray-freeze-drying,SFD )技术对植物乳杆菌( NCIM 2083 )进行包埋。以低聚果糖( FOS )、乳清蛋白( WP )和麦芽糊精( MD )为包合剂,在不同配方下进行包合提高了免疫球蛋白水平,对吞噬作用有调节作用。
胡蕾等[63]在奶牛日粮中加入枯草芽胞杆菌、乳酸菌和丁酸梭菌形成复合益生菌制剂,通过观察奶牛乳品质发现,随着服用复合益生菌制剂的时间增加,乳品质有明显提高,具体表现在乳糖率、乳蛋白率、总固形物含量的显著提升。夏元军等[64]在奶牛日粮中添加3×108CFU/(头·d)嗜酸乳杆菌和枯草芽胞杆菌制成的复合益生菌制剂,结果显示,复合益生菌制剂可使奶牛日平均产奶量提高20%。这些研究均表明益生菌对奶牛机体的有益作用,对预防及缓解牛乳腺炎有一定疗效。但也有一小部分研究阐述了益生菌缓解牛乳腺炎的不足之处,如耐药基因或有害影响、疗效不如抗生素显著等问题。为了使用益生菌代替抗生素并最大限度地发挥预防和控制的优势,还需要进一步的研究。
猜你喜欢
现代临床医学(2021年5期)2021-11-02
昆明医科大学学报(2021年4期)2021-07-23
中国生殖健康(2020年8期)2021-01-18
中国生殖健康(2018年3期)2018-11-06
猪业科学(2018年5期)2018-07-17
幸福(2017年18期)2018-01-03
饲料与畜牧(规模养猪)(2017年9期)2017-10-16
中国乳品工业(2017年1期)2017-02-20
兽医导刊(2016年12期)2016-05-17
中华皮肤科杂志(2014年4期)2014-12-19
