
一株副溶血弧菌噬菌体生物学特性、全基因组特征及其在食品中的应用
2024-02-17 13:20张俊鹏刘文婷王华娟王宏勋
食品工业科技 2024年1期
张俊鹏,刘文婷,石 甜,王华娟,王宏勋,周 敏,
(1.武汉轻工大学食品科学与工程学院,湖北武汉 430000;2.武汉轻工大学生命科学与技术学院,湖北武汉 430000)
副溶血性弧菌(Vibrio parahaemolyticus,Vp)是一种常见于河口和海洋环境的革兰氏阴性嗜盐细菌。食用未加工成熟的海产品是人类感染副溶血性弧菌的主要原因[1]。近年来海鲜类食品在市场广泛流通,导致该菌由沿海向内陆传播蔓延,甚至超过沙门氏菌和志贺氏菌,成为腹泻的首要病原菌[2]。在过去的几十年里,由于抗生素的过度使用,副溶血性弧菌中出现了抗生素耐药性[3]。因此,迫切需要既安全又对消费者友好的新的控制方法。
噬菌体是细菌病毒,在自然界中广泛存在,目前噬菌体在副溶血弧菌控制方面已有多篇报道。袁琳等[4]研究了噬菌体SHOU24 对即食对虾中副溶血性弧菌的抑制效果,结果表明,经噬菌体处理的副溶血性弧菌的生长延滞期增长了4.04 h,噬菌体在常温条件下能使即食虾中副溶血性弧菌的浓度下降1 个数量级,噬菌体SHOU24 可显著降低即食对虾中的副溶血性弧菌数量。郑小双等[5]评价了副溶血性弧菌噬菌体VppMIX 的抑菌效果,结果显示在25 ℃恒温保藏12 h 后,不同剂量噬菌体VppMIX 处理的黄鱼样品中副溶血性弧菌数量比对照组降低了1.41~4.98 lgCFU/g。可见噬菌体十分适合用于海产品等易腐败食品中主要病原菌的控制。
目前,已报道的副溶血性弧菌裂解性噬菌体数量有限,为了丰富该致病菌的噬菌体库,本研究以10 株实验室保存的副溶血弧菌为宿主菌,从湖北省武汉不同地点采集的样品中分离烈性噬菌体,分析其形态、裂解谱、一步生长曲线、pH 及温度稳定性等生物学特征,进行基因组比较,并以人工污染的基围虾作为模型,研究噬菌体在贮藏温度下对模拟污染虾肉的抑菌效果,为开发噬菌体抑制剂提供理论支持。
1 材料与方法
1.1 材料与仪器
23 株副溶血弧菌分离株 武汉轻工大学微生物实验室,其中ATCC17802 为食品源标准菌株,F6、F7、F22、F23 来源于食品,Vp27、Vp33、Vp41、Vp44、460、461、469、470、474、475、485、486、O56、O514 来源于病人,H128、H256、H512 为抗生素诱导株。选取其中10 株细菌用于分离副溶血弧菌噬菌体;虾肉样品 武汉常青花园武商量贩、常青花园菜市场、盒马鲜生(武汉太和里店);TCBS 琼脂 青岛海博生物技术有限公司;Agar 琼脂粉 飞扬生物有限公司;酵母提取素、胰蛋白胨 北京瑞达恒辉公司;氯化钠、无水乙醇 上海国药集团化学试剂有限公司;96 孔微量培养板 美国康宁公司;0.22 μL 针头过滤器 天津津腾公司;一次性注射器 金塔有限公司;DNaseⅠ、RNase A、蛋白酶K Takara 公司。
DRP-9062 型电热恒温培养箱 上海森信实验仪器有限公司;SW-CJ-2FD 型双人单面净化工作台苏州净化设备有限公司;DHG-9053A 电热鼓风干燥箱 上海一恒科学仪器有限公司;PHS-25 pH 计、CP214 型电子天平 奥豪斯仪器公司;S4OPL201 超纯水机 上海乐枫科技有限公司;Mini 15K 离心机杭州奥盛仪器有限公司;DB150L 高压灭菌锅 北京亚欧德鹏科技有限公司;CMBR 全自动生长曲线分析系统 芬兰Bio screen。
1.2 实验方法
1.2.1 噬菌体的分离与纯化 取虾肉样品25 g 加入225 mL 生理盐水,均质2 min,混合液以5000 r/min离心10 min,取5 mL 上清液加入5 mL LB 液体培养基中,同时分别加入100 μL 处于对数生长期的10 株副溶血弧菌,37 ℃振荡培养8 h 后5000 r/min离心10 min,取上清液过0.22 μm 滤膜,滤液经10 倍梯度稀释后与宿主菌混合,使用双层平板法[6],37 ℃培养6~8 h 检测噬菌斑。挑取单个空斑至1 mL SM 缓冲液中,4 ℃过夜,用SM 缓冲液稀释噬菌体液,取适宜梯度加入宿主菌制成双层平板,37 ℃培养6~8 h,重复培养5 次,得到纯化噬菌体。
1.2.2 噬菌体效价测定 纯化的噬菌体液进行10 倍稀释,将10-5、10-6及10-7的噬菌体液100 μL 分别与100 μL 对数期的宿主菌混合,利用双层平板法[6]观察噬菌斑的个数。
噬菌体效价(PFU/mL)=噬菌斑数量×稀释倍数×10
1.2.3 噬菌体透射电镜观察 参考Ramírez-Orozco等[7]方法,采用磷钨酸负染法进行透射电镜观察噬菌体形态,并利用软件Digital Micrograph Demo 3.9.1测量噬菌体头部直径和尾长。
1.2.4 噬菌体宿主谱测定 使用点斑法测定噬菌体的宿主谱[8],取3.7 mL LB 半固体培养基加入0.1 mL宿主菌(109CFU/mL),颠倒混匀后倾倒在固体培养基上,待凝固后,滴加噬菌体悬液(109PFU/mL)10 μL,置37 ℃培养箱中培养8~12 h,观察裂解圈的形成。
1.2.5 噬菌体的最佳感染复数(multiplicity of infection,MOI)按照不同的MOI 值(0.001、0.01、0.1、1、10、100、1000)将噬菌体与宿主菌混合,对不同MOI 值的噬菌体效价进行测定[6],效价最高的MOI值则为噬菌体的最佳感染复数。
1.2.6 噬菌体的一步生长曲线 将噬菌体原液与宿主菌以最佳感染复数混合,在37 ℃水浴锅中温浴10 min,使噬菌体尽可能多地吸附到宿主菌上。将混合液4 ℃、8000 r/min 离心2 min,弃上清液使用等体积LB 培养基(3% NaCl)重悬,重悬2 次,取1 mL重悬液加入到9 mL LB 培养基(3% NaCl)。从0 min开始,每隔10 min 取500 μL 于4 ℃、8500 r/min 离心2 min,测定噬菌体效价[6]。
1.2.7 噬菌体的热稳定性 提前准备不同温度(30、40、50、60、70 ℃)的水浴锅,将噬菌体原液稀释到107PFU/mL,取1 mL 噬菌体原液于1.5 mL 离心管中,将离心管放入不同温度的水浴锅中温浴。从0 min开始,每隔30 min 测定离心管中噬菌体效价[6]。
1.2.8 噬菌体的pH 稳定性 提前准备不同pH 的PBS 缓冲液,将100 µL 噬菌体效价为107PFU/mL的噬菌体原液加入到900 µL 预先准备好的不同pH的PBS 缓冲液中,使得混合后体系的最终pH 分别为(2、3、4、5、6、7、8、9、10、11、12、13),于37 ℃水浴锅中温浴2 h。使用双层平板法测定噬菌体效价[6]。
1.2.9 噬菌体全基因组提取和功能基因分析 噬菌体全基因组提取采用苯酚-氯仿法,具体步骤参考文献[9]。委托上海泽塔公司对提取的噬菌体474x1基因组进行全基因组测序,并对噬菌体基因组进行拼接,得到完整的基因组序列。使用DNA Statistics(http://www.geneinfinity.org/sms/sms_dnastats.html)对噬菌体基因组序列进行分析,包括四种碱基的组成比例、GC 平均含量。使用JSpeciesWS(https://jspecies.ribohost.com/jspeciesws/)对从NCBI 数据库上下载的10 株弧菌噬菌体全基因组完整序列进行平均核苷酸同一性(ANI)统计,得到矩阵。使用微生信在线工具(http://www.bioinformatics.com.cn/plot_basic_matrix_heatmap_064)绘制热图。使用RAST 在线注释工具(https://rast.nmpdr.org/)对全基因序列进行基因预测。使用tRNAscan-SE(https://www.swmath.org/software/8005)在线工具预测全基因组中有无tRNA 基因。使用NCBI 数据库的BLASTn 对预测的各个基因进行逐个检索分析,推测其基因功能。用BRIG(0.95)软件绘制基因组圈图。使用VFDB数据库(https://www.mgc.ac.cn/VFS/)预测噬菌体474x1 基因组中可能存在的毒力基因,使用CARD数据库(https://card.mcmaster.ca/)预测基因组中可能存在的抗生素抗性基因。使用MEGA 7.0 软件建立基于末端酶大亚基的进化树,分析噬菌体474x1 的亲缘性。
1.2.10 噬菌体对基围虾中副溶血性弧菌的抑制作用参考葛强等[10]的实验步骤稍作改进,将购买的新鲜虾去壳,用无菌刀将虾肉切为1 cm×1 cm 的样品,重约1 g,将肉样煮沸30 min 后置于无菌平皿中。在样品中接种100 μL 1.0×105lgCFU/mL 副溶血性弧菌474,于超净台中风干10 min 使其吸附后用封口膜封住,作为对照组。实验组应在副溶血性弧菌474 吸附后另外分别接种100 μL 1.0×108lgCFU/mL(MOI=1000)和100 μL 1.0×109lgCFU/mL(MOI=10000)的噬菌体悬液,常温条件下静置10 min 使其吸附后用封口膜封住。将样品置于4 ℃,在第0、3、6、9、12 h 分别取样,将样品放入10 mL 离心管中,加入5 mL 生理盐水,3000 r/min 离心10 min,使虾肉沉淀,取上清液进行适当梯度稀释,取1 mL 稀释液采用平板计数法检测虾肉中副溶血性弧菌的菌量,从而检测噬菌体的抑菌效果。
1.3 数据处理
采用Excel 2019 计算数据平均数和标准差、Origin 2022 进行折线图绘制和IBM SPSS Statistics 23 的独立样本T检验进行显著性分析。
2 结果与分析
2.1 副溶血弧菌噬菌体形态学特征
以副溶血弧菌474 菌株为宿主菌从海鲜市场基围虾样品中成功分离出1 株裂解性较强副溶血弧菌噬菌体,命名为474x1。培养后474x1 可在474 菌株的菌苔上形成清晰、透明的噬菌斑,直径1.5~2 mm(图1)。电镜下观察噬菌体的头部呈六边形,直径约70.8 nm,尾长约18.3 nm。具有典型的有尾噬菌体目短尾病毒科病毒形态特征(图2)。
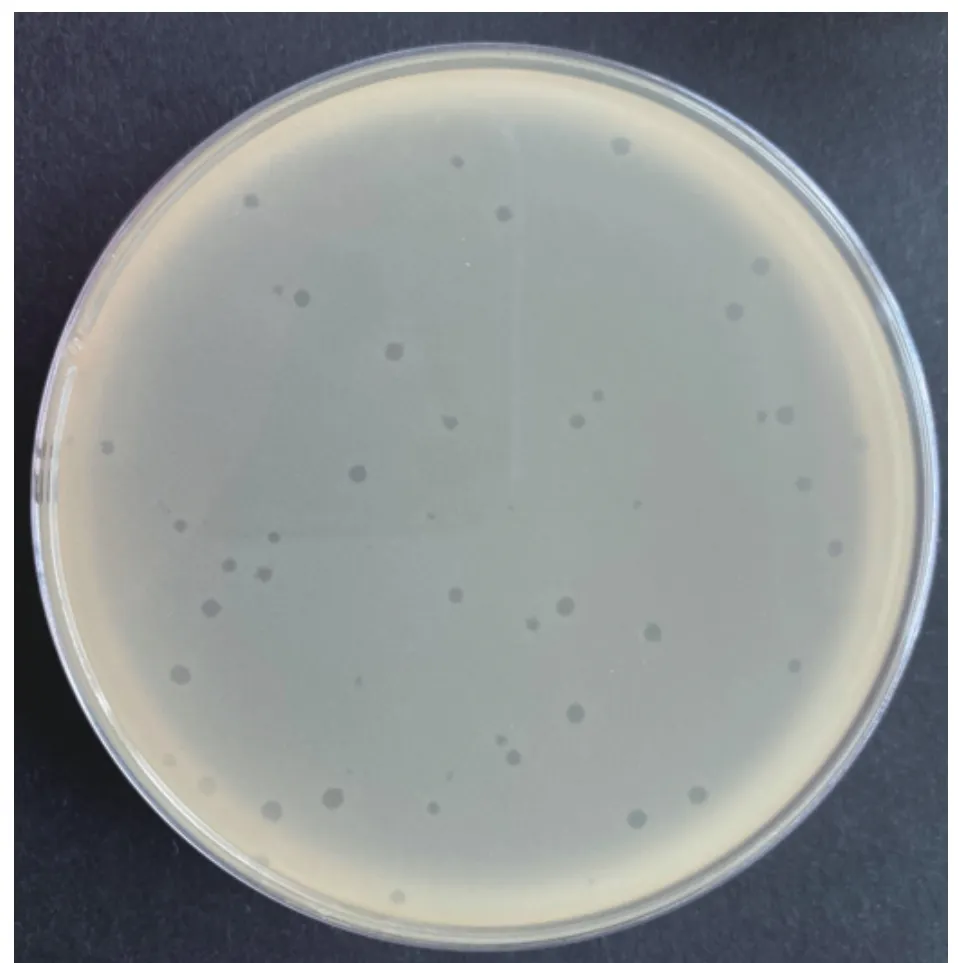
图1 副溶血弧菌噬菌体474x1 噬菌斑形态Fig.1 Plaque morphology of V.parahaemolyticus phage 474x1

图2 副溶血弧菌噬菌体474x1 透射电镜形态Fig.2 Transmission electron microscope morphology of V.parahaemolyticus phage 474x1
2.2 副溶血弧菌噬菌体474x1 宿主谱
副溶血弧菌噬菌体474x1 宿主谱如表1。噬菌体474x1 能够裂解23 株副溶血弧菌中的19 株(19/23=82.61%),表明噬菌体474x1 对耐药副溶血弧菌具有良好的裂解能力。
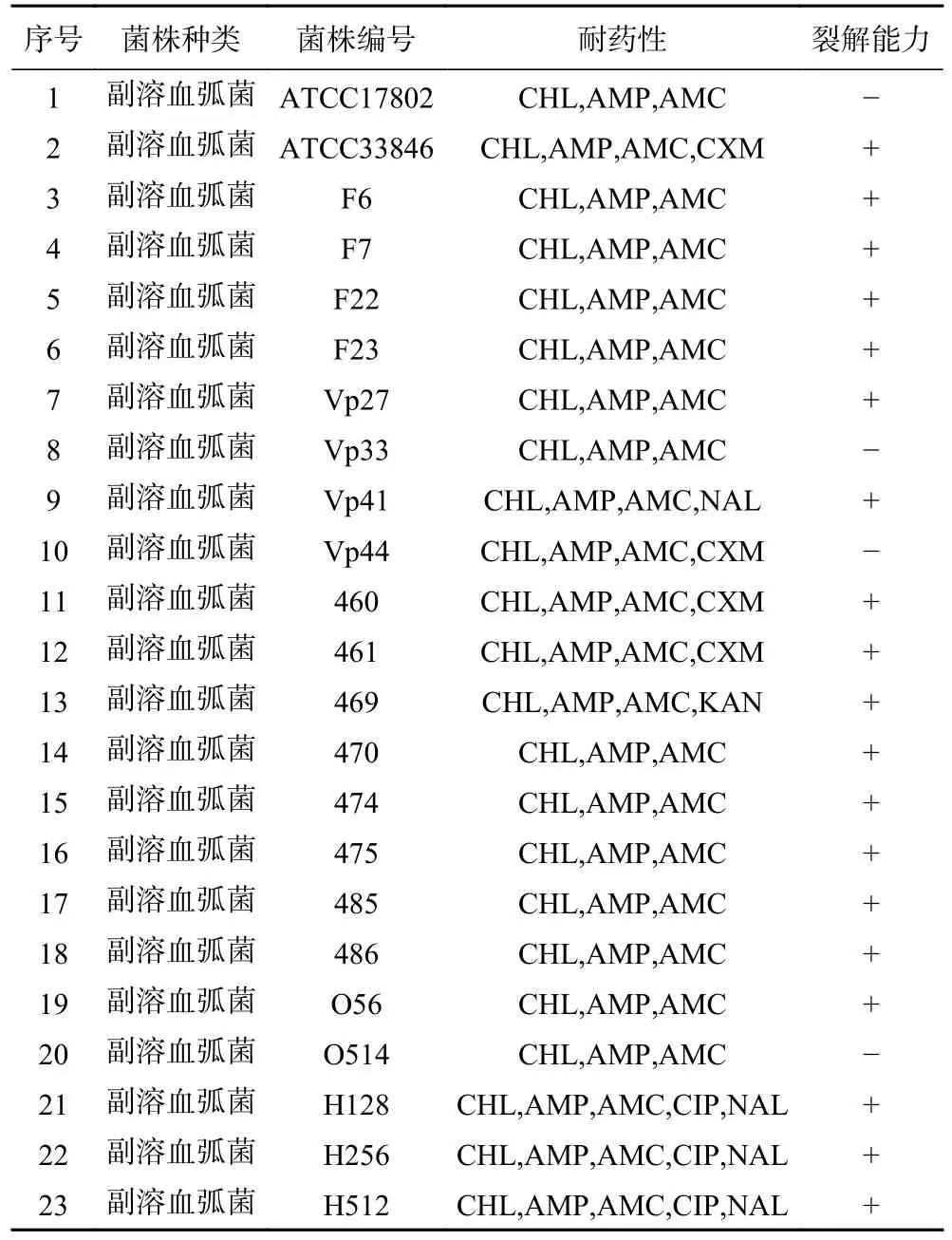
表1 副溶血弧菌噬菌体474x1 宿主谱Table 1 Host range of V.parahaemolyticus phage 474x1
2.3 副溶血弧菌噬菌体474x1 最佳感染复数
表2 为副溶血弧菌噬菌体474x1 效价测定结果。当MOI 为0.01 时,噬菌体效价达到最大为9.66 lgPFU/mL,说明噬菌体474x1 在感染副溶血弧菌474 时,噬菌体与宿主菌的数量的比值为0.01 时,可增殖最多的噬菌体。
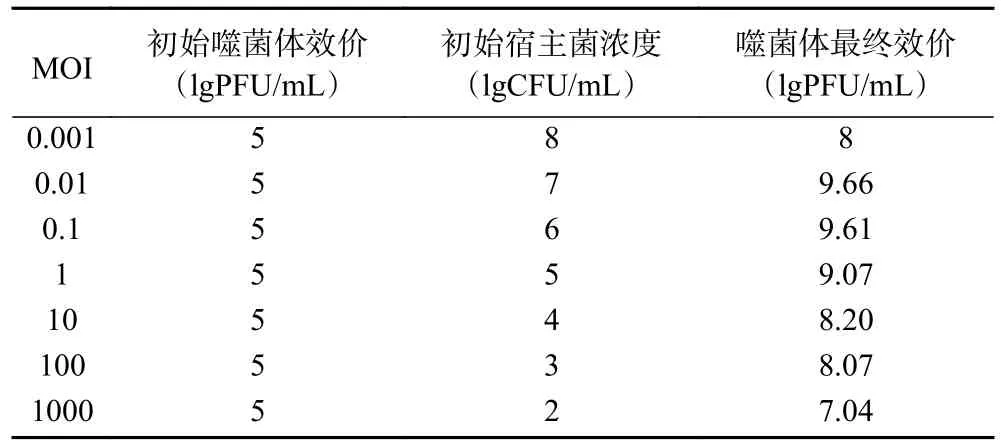
表2 副溶血弧菌噬菌体474x1 的最佳感染复数Table 2 Multiplicity of infection of V.parahaemolyticus phage 474x1
2.4 副溶血弧菌噬菌体474x1 一步生长曲线
以培养时间为横坐标,以反应体系中噬菌体效价的对数为纵坐标绘制噬菌体的一步生长曲线。如图3 所示,噬菌体474x1 的潜伏期为10 min,裂解期为10~90 min,裂解末期噬菌体效价达到8.81 lgPFU/mL,裂解量为115 PFU/cell。
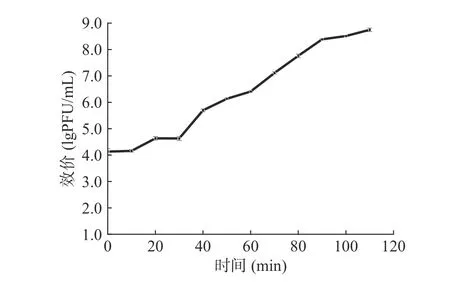
图3 副溶血弧菌噬菌体474x1 一步生长曲线Fig.3 One-step growth curve of V.parahaemolyticus phage 474x1
2.5 副溶血弧菌噬菌体474x1 热稳定性和pH 稳定性
噬菌体对温度的敏感性如图4 所示,474x1 在30~50 ℃的范围内活性稳定,70 ℃时其活性迅速下降,30 min 被完全灭活;60 ℃孵育1 h 后效价可减少1 个数量级。474x1 的效价在pH4~11 之间均较稳定,但pH>11 和pH<4 条件下孵育2 h 后活性丧失(图5)。
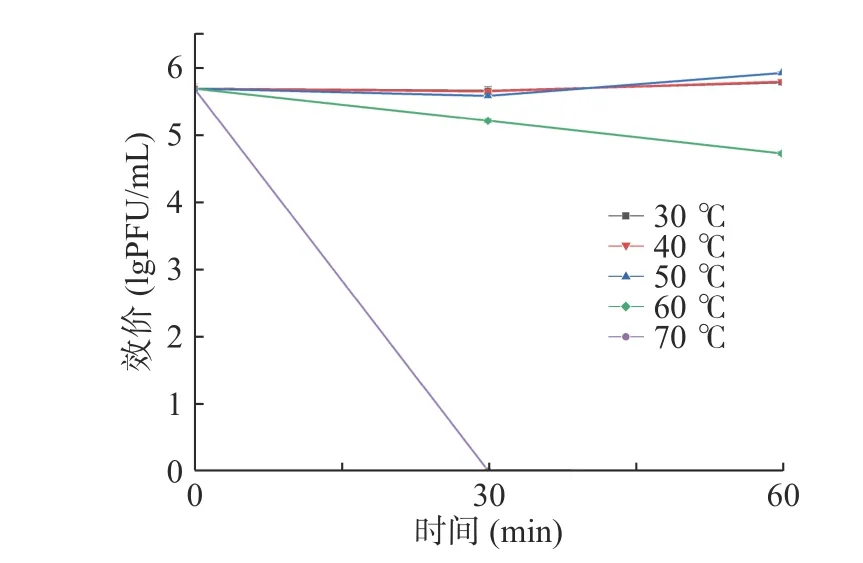
图4 副溶血弧菌噬菌体474x1 热稳定性Fig.4 Thermal stability of V.parahaemolyticus phage 474x1
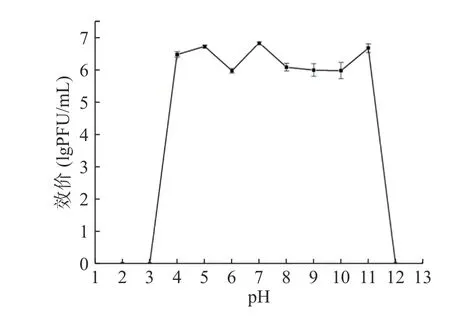
图5 副溶血弧菌噬菌体474x1 pH 稳定性Fig.5 pH stability of V.parahaemolyticus phage 474x1
2.6 副溶血弧菌噬菌体474x1 全基因组DNA 的提取
使用苯酚-氯仿法提取了噬菌体474x1 全基因组DNA,用超微量分光光度计测定提取的噬菌体474x1 DNA 终浓度为220 ng/mL 且OD260/280为1.88,说明提取出的DNA 浓度适宜且含有的杂质较少,满足测序要求。
噬菌体474x1DNA 进行琼脂糖凝胶电泳结果如图6 所示,可以发现DNA 电泳条带单一无其它条带,且DNA 分子量较大,说明完整地提取出了噬菌体474x1 的基因组DNA。
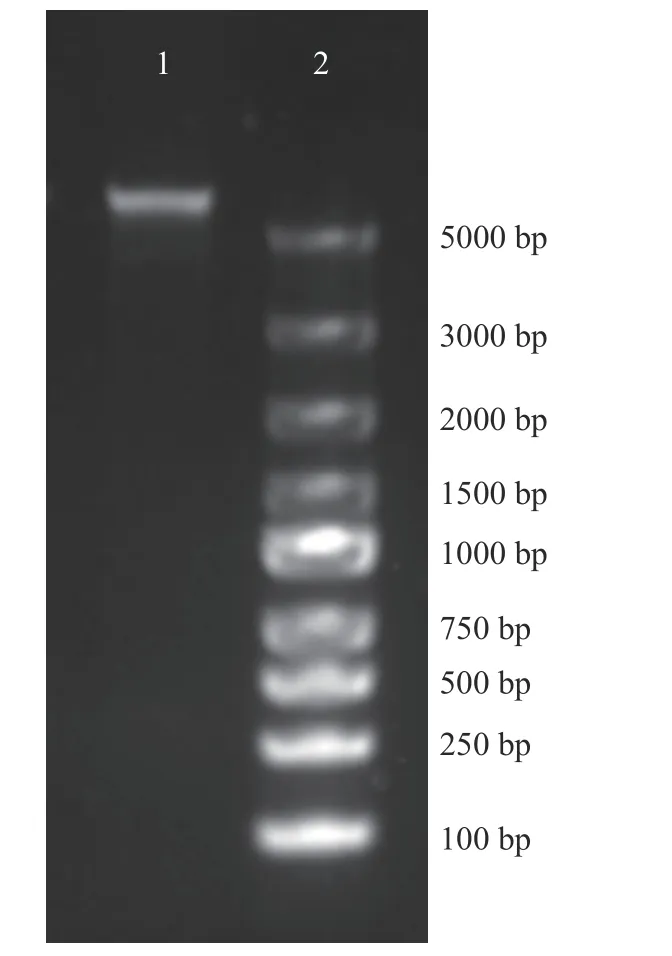
图6 噬菌体474x1 的全基因组提取Fig.6 Extraction of whole genome DNA of V.parahaemolyticus phage 474x1
2.7 副溶血弧菌噬菌体474x1 全基因组分析
噬菌体474x1 的基因组长度为47830 bp,碱基分布情况为A(30.42%)、T(29.16%)、C(20.45%)、G(19.96%),GC 平均含量为40.41%。噬菌体474x1全基因组序列BLASTn 比较结果显示,噬菌体474x1 与副溶血弧菌噬菌体Vp41s3(索引号:MZ0 73369.1)有较高的同源性,同源性99.06%,覆盖率为77%。但与其他弧菌噬菌体同源性均小于80%,如与霍乱弧菌噬菌体ICP2(索引号:HQ641345.1)同源性为78.55%,与创伤弧菌噬菌体Saratov-15(索引号:MT767883.1)同源性为78.45%。ANI 热图显示474x1 与副溶血弧菌噬菌体Vp41s3 同源性最高(同一性×覆盖率=73%),但与其他噬菌体基因组同源性均小于50%(图7)。
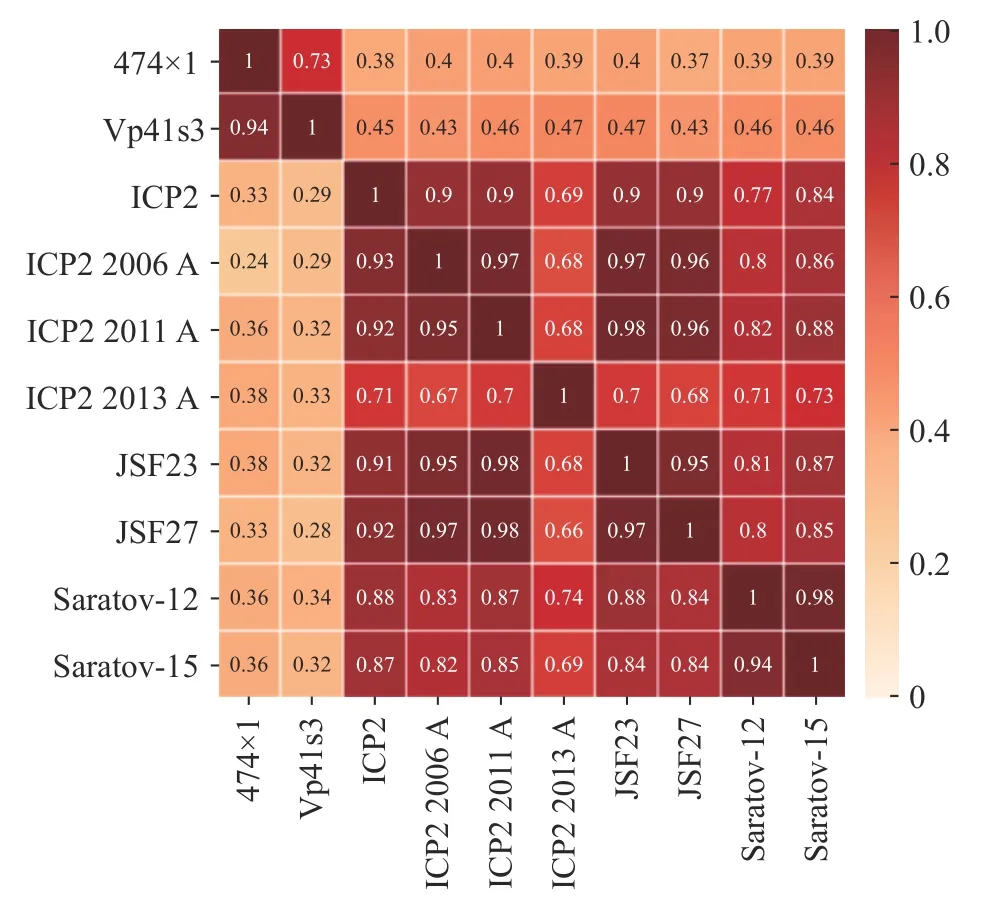
图7 平均核苷酸同源性热图Fig.7 Average nucleotide identity heatmap
2.8 副溶血弧菌噬菌体474x1 基因组功能基因注释
使用RAST 在线注释工具进行功能基因注释。结果显示,噬菌体474x1 基因组共有69 个开放阅读框(open reading frame,ORFs),其中有24 个ORFs位于正链上,45 个ORFs 位于负链上。基因组长度为47830 bp,最长的ORFs 为4245 bp,最短的仅126 bp,平均长度643 bp。ORFs 总长为44376 bp,占全长的92.78%。经BLAST 比对,推测出的69个ORFs 中有14 个ORFs 已确定功能。通过这些特征基因功能将其分为4 个模块,包括DNA 复制与调控模块(ORF29、ORF35、ORF36、ORF39、ORF45、RF47、ORF49、ORF51、ORF54、ORF55)、DNA包装模块(ORF4、ORF5)、结构模块(ORF1)、附加功能模块(ORF32)。除了这14 个具有特定功能的基因外,其他的ORFs 都为假定蛋白,约占79.71%,其基因组图谱如图8 所示。采用tRNAscan-SE 筛查全基因组中无tRNA 基因,表明474x1 依赖宿主翻译。通过VFDB 数据库和CARD 数据库预测噬菌体474x1 基因组不含毒力基因与抗生素抗性基因。因此,该结果表明噬菌体474x1 应用于食源性致病菌防控的安全性。
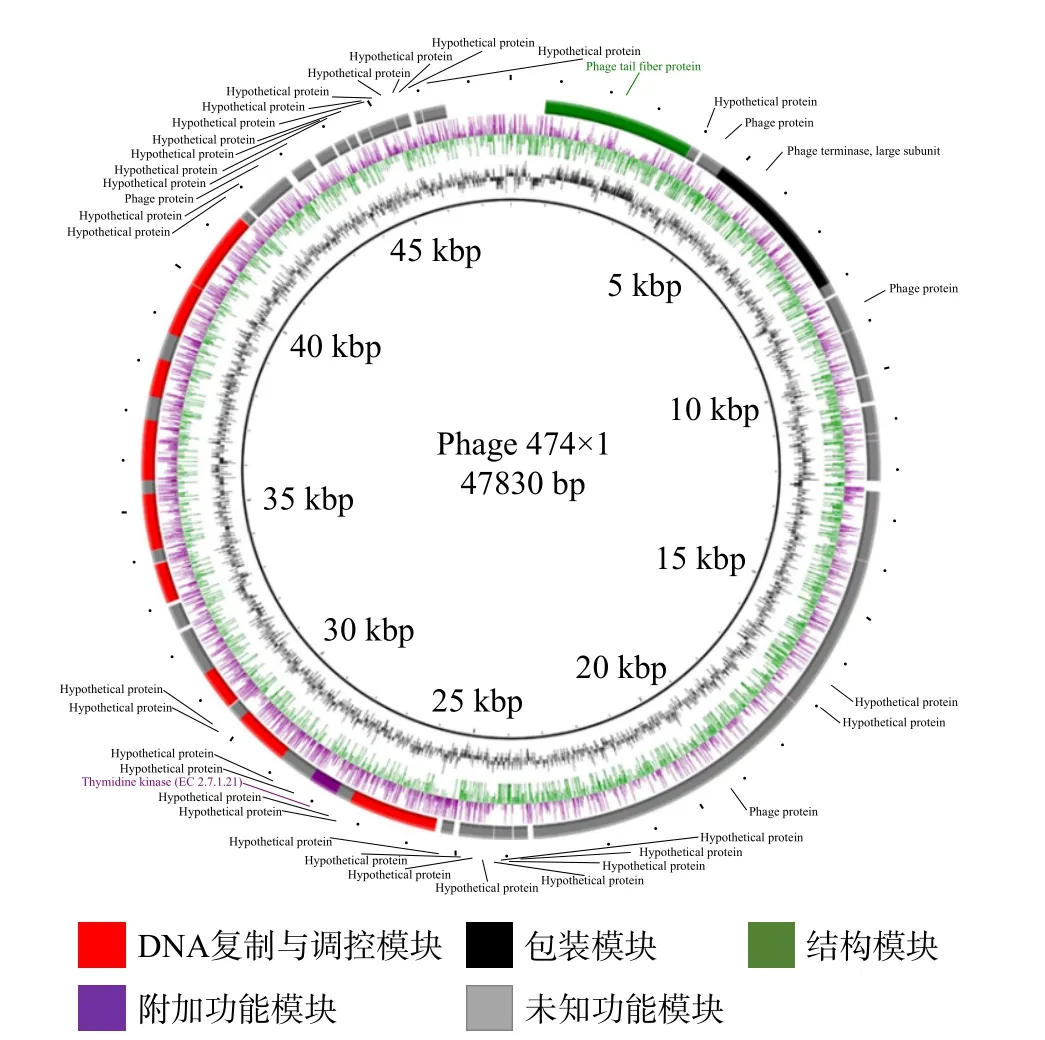
图8 噬菌体474x1 的基因组图谱Fig.8 Genomic map of phage 474x1
2.9 副溶血弧菌噬菌体474x1 进化树分析
基于噬菌体474x1ORF4 末端酶大亚基构建的系统发育进化树如图9 所示,副溶血弧菌噬菌体474x1 与副溶血弧菌噬菌体Vp41s3(短尾噬菌体)在同一个进化分支上,另外与霍乱弧菌噬菌体ICP2,创伤弧菌噬菌体Saratov-12、Saratov-15 形成一簇,有着很近的亲缘关系。

图9 基于末端酶大亚基构建的系统进化树Fig.9 Phylogenetic tree constructed based on terminal enzyme large subunit
2.10 噬菌体对基围虾中副溶血性弧菌的抑制作用
虾肉在4 ℃放置12 h,副溶血弧菌浓度变化如图10A 所示。在放置期间,接种5 lgCFU/mL 细菌的对照组菌量没有明显增长,从第0 h 的3.73 lgCFU/mL 下降到第12 h 的3.55 lgCFU/mL。实验组的菌量相比于对照组在12 h 内均有明显下降,具有统计学差异(P<0.05),MOI=1000 实验组在第3 h 相较对照组菌量下降了0.39 lgCFU/mL,MOI=10000 实验组在第12 h 相较对照组菌量下降了0.92 lgCFU/mL,具有极显著性差异(P<0.01)。
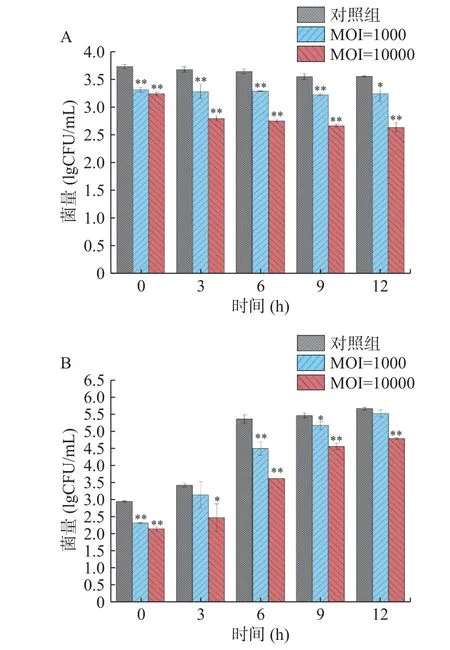
图10 副溶血弧菌噬菌体474x1 对虾肉中副溶血弧菌抑菌效果实验Fig.10 Bacteriostatic effect of phage 474x1 on V.parahaemolyticus in prawn meat
虾肉在25 ℃放置12 h,副溶血弧菌浓度变化如图10B 所示。在放置期间,接种5 lgCFU/mL 细菌的对照组菌量呈稳步增长的趋势,在第12 h 增长到5.66 lgCFU/mL。实验组的菌量相比于对照组在12 h 内均有明显下降,MOI=1000 实验组在第6 h 相较对照组菌量下降了1.04 lgCFU/mL,MOI=10000在第6 h 相较对照组菌量下降了1.82 lgCFU/mL,具有极显著性差异(P<0.01)。
3 讨论与结论
目前,国内已有学者相继分离出副溶血弧菌烈性噬菌体,丁云娟等[11]从水产品市场的污水中分离出来一株烈性噬菌体qdvp001,彭勇等[12]从青岛岸边海水中分离出两株副溶血弧菌噬菌体VPp2、VPp3。本实验室周敏等[13]从海鲜内脏、污水中分离出一株烈性噬菌体F23s1。本实验也从基围虾样品中分离出宽谱噬菌体474x1,对研发生物抑菌剂有重要价值。
本研究中,宿主谱的结果表明474x1 能够裂解23 株耐药副溶血弧菌中的19 株,表现出较宽的裂解谱范围,宿主谱的差异可能是由于细菌细胞表面存在的受体的差异影响了噬菌体的吸附[14]。此外,噬菌体吸附具有高度特异性,这也意味着噬菌体对其他细菌是无害的,特别是那些有益的菌群。这也表明,474x1可以在未来用于防治特定的细菌性疾病,如弧菌病。
MOI 指的是噬菌体与宿主菌的比率。不同的噬菌体感染和杀灭细菌的方式不同,因此具有不同的最佳MOI。例如两株副溶血性弧菌裂解噬菌体VB_VPS_BA3 和VB_VPS_CA8,最佳MOI 为0.1[15]。Ibrahim 等[16]对一株能够裂解溶藻弧菌、哈维氏弧菌和副溶血弧菌的烈性噬菌体PhVh6 测定的最佳MOI为1。在本研究中,噬菌体增殖的最佳MOI 为0.01,表明以较低数量噬菌体感染宿主菌时,最终能获得高效价的噬菌体,在生产中可用较低的成本得到大量噬菌体。
研究噬菌体的一步生长曲线可以发现噬菌体的生长规律[17]。挑选出潜伏期短裂解量大的噬菌体,对提高工业生产有巨大意义。474x1 的潜伏期与噬菌体VppYZU92[5]相似,后者的潜伏期较短为10 min,裂解量为35 PFU/cell。另一株噬菌体VppYZU64的潜伏期较长为35 min,但裂解量为150 PFU/cell[5]。在本研究中,474x1 的潜伏期较短且裂解量偏大,为10 min 和115 PFU/cell,十分适合规模化制备。
在各种胁迫条件下对噬菌体效价稳定性的测定结果有助于为噬菌体在细菌控制中的应用提供参考[18-19]。在本研究中,测定了噬菌体对温度和pH 的抵抗力,以确定这些噬菌体对副溶血性弧菌感染的生防效果。温度是影响噬菌体存活的一个重要因素[20]。它在噬菌体的附着、穿透和增殖过程中起着重要作用[21]。在本研究中,474x1 在50 ℃下稳定,在60 ℃孵育1 h 后效价下降,在70 ℃以上30 min 后完全失活,当应用于食品粗加工环境中时,它仍然可以存活。此外,环境的酸碱度会影响噬菌体的稳定性[19]。据报道,低pH 影响噬菌体聚集,降低其在细菌细胞上的吸附能力[22]。474x1 在较宽的pH 范围保持高效价,与vap04 相似[23]。
本实验选择了2 种贮藏温度,其中,4 ℃代表保藏温度,25 ℃代表室温。4 ℃下副溶血弧菌菌量均低于25 ℃下同一时间点菌量,这可能是因为低温会抑制副溶血弧菌的生长。随着贮藏时间的延长,25 ℃下MOI=1000 和MOI=10000 实验组中的菌量均明显下降,在第12 h 时实验组中副溶血性弧菌数量分别比对照组降低0.18、0.87 lgCFU/mL,说明噬菌体474x1 能够显著(P<0.01)抑制宿主菌的生长,但不能完全杀灭副溶血弧菌。高璐[23]在噬菌体对鱼汁的抑菌实验中,三株噬菌体Vmp03、Vpp07、Vap04 在鱼汁分别对其宿主菌均有一定的抑制能力。在25 ℃条件下,经过Vmp03 处理后的实验组相较对照组的菌量在第8 h 下降约2 lgCFU/mL;经过Vpp07、Vap04 处理后的实验组相较对照组的菌量在第6 h 下降约1 lgCFU/mL。本实验在25 ℃下MOI=1000,MOI=10000 的实验组相较对照组在第6 h 时菌量分别下降1.04、1.82 lgCFU/mL,抑菌效果和高璐的研究相似。
综上所述,474x1 是一株新的副溶血性弧菌噬菌体。该噬菌体裂解谱宽、裂解量大、潜伏期短。它在高温下失活,但在60 ℃以下表现出较好的稳定性,能耐受宽范围的pH,在食品中也有良好的抑菌效果,由于缺乏抗生素抗性基因和毒力基因,474x1 在不久后将会用来防控水产品副溶血性弧菌。
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
昆明医科大学学报(2021年5期)2021-07-22
中国动物传染病学报(2021年3期)2021-07-21
当代水产(2019年5期)2019-07-25
广东海洋大学学报(2015年3期)2015-12-22
药学与临床研究(2015年4期)2015-06-05
现代检验医学杂志(2015年4期)2015-02-06
沈阳医学院学报(2014年4期)2014-12-27
