
盐度和pH对平凡舟形藻生长和油脂积累的影响
2024-02-07 07:29:46信宏颖梁金燕王馨彤范亚文陆欣鑫隋丰阳
水生生物学报 2024年2期
信宏颖 梁金燕 王馨彤 刘 妍, 范亚文, 陆欣鑫, 隋丰阳,
(1.哈尔滨师范大学生命科学与技术学院, 哈尔滨 150025; 2.黑龙江省水生生物多样性研究重点实验室, 哈尔滨 150025)
利用可再生资源生产生物燃料是进行可持续发展的选择[1]。微藻在工业上的应用已经引起了人们的关注, 但由于微藻对环境变化敏感, 进一步影响微藻产油效率[2]。大多数微藻具有将大气中的二氧化碳通过光合作用转化为生物质、脂肪酸和脂类等物质储存在细胞内[3], 不仅使微藻在生产生物燃料方面具有开发潜力, 在生产高价值脂类化合物方面具有更高的应用价值[4]。微藻的总脂质含量约为干细胞重量的1%—70%, 油脂主要以甘油三酯和脂肪酸甲酯的形式存在, 适合生产生物柴油[5,6]。据报道, 许多微藻物种细胞中含有大量的脂质, 它们的脂质可以很容易地通过酯交换过程转化为生物柴油, 因此表明了它们作为未来生物能源替代来源的重要性[7]。微藻分布范围广, 可以在多种水生生境中正常生长, 包括湖泊、池塘、河流和海洋,甚至废水, 也可以在人为调控的不同条件下培养[8]。微藻具有广阔的应用前景, 但目前微藻规模化生产中仍存在产量低和成本高的问题。微藻在培养过程中还需要严格控制各种环境因素, 如碳源[9]、盐度[10]、氮[11]、温度[12]、CO2浓度[13]和光照强度[14]等。少量的微量营养素, 如铁[15]和锌[16]等也是影响藻类生长和代谢的重要因素, 微藻利用各种机制来适应环境胁迫, 例如改变其形态和物质组成[17]。因此, 生产中可以使用不同应激源来提高某些藻类中色素和脂类成分。其中盐度和pH是本研究的重点影响因素。
盐度是影响微藻生长、脂肪含量和生化组成的重要因素之一。由于藻种的生理差异, 微藻对盐度的适应性也不同[18]。在微藻生长过程中, 盐胁迫会影响类胡萝卜素合成和脂肪积累, 并影响脂肪酸组成[19]。有报道称,Botryococcus braunii可以有效地将盐排除在细胞外, 并在生长过程中积累甘油,作为对胞外盐浓度的直接反应[20]。通过增加盐度来胁迫藻类细胞, 可能会导致额外的脂肪积累[21]。在微藻生长过程中, 介质中的pH也起着重要的作用, 因为其影响二氧化碳浓度, 会导致物种的分布和碳的利用率不同。据报道, 莱茵衣藻的细胞器和其他内部过程需在胞液pH 7—7.1才能发挥作用[22],在酸性的外部条件下, 额外的ATP被消耗以维持胞液pH为7, 导致莱茵衣藻在酸性环境中的能效较低。当多余的ATP被耗尽时, 用于细胞生长和繁殖的能量就会减少, 在碱性pH中观察到相反的反应。此外, 叶绿素含量也受到pH的影响, 因为pH决定了光合作用过程[23]。因此, 研究盐和pH胁迫对藻株的生长和生化的影响尤为重要。
平凡舟形藻(Navicula trivialis)是本实验室分离的一株生长快的淡水底栖硅藻, 该藻隶属于硅藻门、硅藻纲、舟形藻目、舟形藻科、舟形藻属, 经实验室分离培养初步得到油脂含量较高的藻株, 有关该藻的生态习性及培养方法迄今为止鲜有报道。利用微藻生产生物柴油被认为是解决资源消耗、能源短缺和环境污染等问题的一种很有前途的策略[9]。因此, 藻类生物量和脂质生产力的优化是藻类生产生物燃料研究的主要重点。本研究将平凡舟形藻在不同NaCl浓度和pH条件下进行培养, 分析该藻株的最佳培养条件和油脂产量, 本研究的结果可能有助于了解不同的盐度和pH如何提高微藻生长和油脂产量, 为进一步提高其油脂的产量奠定基础。
1 材料与方法
1.1 藻种培养
本研究所用平凡舟形藻藻种由哈尔滨师范大学水生生物实验室采集并分离纯化, 采用水滴分离法, 用移液管和显微镜对微藻细胞进行多次分离,得到纯化的平凡舟形藻, 并将平凡舟形藻藻种转接至WC培养基中进行培养, WC培养基如表1所示。挑选生长良好的纯种藻种放入光照培养箱中进行逐级扩大培养, 培养温度为(25±1)℃, 光照条件为4000 lx, 光暗比时间为12h∶12h, 藻液每天定时摇动3—5次, 防止藻细胞附着在瓶壁上, 培养周期12d,达到对数生长期, 用于后续实验。
1.2 盐和pH胁迫实验
将平凡舟形藻培养至对数生长期进行实验,盐和pH的 初始接种藻种密度分别为3×105和1×105cell/mL。采用不同浓度的盐度和pH对平凡舟形藻分别进行单一盐和pH的胁迫处理。盐胁迫浓度分别为0、0.03、0.06、0.12、0.18和0.24 mol/L。用1.00 mol/L HCl和NaOH调节培养基pH至4.5、6.0、7.5、8.5、9.5和10.5。在相同的实验条件下, 每个处理重复3次, 每天摇瓶5次, 将平凡舟形藻放入WC培养基中, 置于(25±1)℃的恒温培养箱中培养, 光照强度4000 lx, 光暗比为12h∶12h, 分别在第0、第2、第4、第6、第8、第10和第12天时取样进行相关指标的测定。
1.3 测定指标及方法
细胞密度依照设定的盐浓度和pH胁迫后,采用分光光度法和血球计数法相结合的方法对平凡舟形藻细胞数量进行统计。得到细胞密度和透光率对应的标准曲线。取各处理组2 mL硅藻悬浮液, 在680 nm波长处测定透光率(以WC培养液为空白组)。根据标准曲线计算细胞数, 各处理组重复3次, 记录12天内平凡舟形藻细胞数量的变化。
叶绿素a含量采用丙酮萃取法测定藻细胞中的叶绿素a含量[24]。取不同盐浓度和pH胁迫后的藻液10 mL, 离心弃上清液, 使用5 mL 80%的丙酮溶液进行超声提取(40 kHz, 30min)。样品在4℃下提取24h, 在646和663 nm波长处测定吸光度(以80%的丙酮进行调零), 叶绿素a含量由以下公式计算:
式中,A663为663 nm处的吸光度;A646为646 nm处的吸光度。
叶绿素荧光参数取5 mL平凡舟形藻的藻液在暗处理下适应30min后, 使用雅欣叶绿素荧光仪(Yaxin-1161)测定平凡舟形藻相关叶绿素荧光参数[25]。
总脂含量使用Ryckebosch[26]的方法将培养12天的微藻离心, 弃上清(m总), 藻液与4 mL甲醇混合。加入氯仿2 mL和蒸馏水0.4 mL, 涡旋震荡混合2min, 再加入氯仿和蒸馏水各2 mL, 涡旋震荡混合。以2000 r/min的速度离心10min, 弃上层, 将下层转移到1个离心管中。剩余固体用4 mL氯仿-甲醇1∶1反复萃取3次。使用旋转蒸发仪进行干燥,称取脂肪收集瓶和脂肪总质量(m1),用旋转蒸发仪在40℃下旋转蒸发除去溶剂(m2), 然后用重量法测定脂肪含量。每个处理重复3次, 用以下公式进行计算:
式中,X为样品中脂肪含量;m1为脂肪收集瓶和脂肪总质量;m2为脂肪收集瓶质量;m总为样品的质量。
数据处理所有处理3次重复, 数据分析都在IBM SPSS 26.0软件中进行统计分析, 多重比较采用LSD法进行, 差异显著性水平为0.05 (P<0.05)。
2 结果
2.1 盐浓度和pH对平凡舟形藻生长的影响
盐浓度的改变对平凡舟形藻的生长有显著性差异。图1A显示了平凡舟形藻经不同盐浓度胁迫培养12d后的生长情况, 平凡舟形藻的生长在各处理组不同水平间表现出明显的差异。平凡舟形藻细胞密度在盐浓度0—0.12 mol/L时随着浓度的升高而增加, 在盐度为0.12 mol/L细胞密度增长最快,第6天细胞密度显著高于对照组(P<0.05), 后随着盐浓度的继续升高会显著降低。培养时间在第8天,盐度为0.12 mol/L时, 细胞密度达到最大值, 为5.5×105cell/mL。在盐浓度为0.03和0.06 mol/L时,生长呈缓慢上升的趋势, 细胞密度最高可达4.3×105和4.9×105cell/mL。在盐度为0.24 mol/L时,平凡舟形藻的细胞密度随着培养时间的增长一直呈现下降趋势, 说明盐度过高不利于平凡舟形藻的细胞密度的增加。
在本研究中, 平凡舟形藻能够在所测试的大多数pH(6.0、7.5和8.5)条件下生长(图1B), 其中pH为7.5在第10天时平凡舟形藻细胞密度增加到最大值为1.4×106cell/mL, 细胞密度相较于其他处理水平有显著增加(P<0.05), 其次细胞密度增长最快的是pH为8.5时。过酸或过碱的环境会导致平凡舟形藻的生长减慢, 如在pH为4.5、9.5和10.5细胞密度快速下降, 最高值分别为3.3×105、8.6×105和6.6×105cell/mL, 细胞密度比pH 7.5时低, 所以平凡舟形藻最适生长的pH为7.5。
2.2 盐浓度和pH对平凡舟形藻叶绿素a含量的影响
在不同的盐浓度下, 叶绿素a含量随盐度变化而变化。如图2A显示, 在6种盐浓度下的平凡舟形藻叶绿素a的含量呈现先升后降的趋势。在0.03和0.06 mol/L从第2天叶绿素a含量开始增加, 在第8天逐渐下降。当盐浓度为0.12 mol/L时, 叶绿素a含量增长最快, 与对照组相比有显著增加(P<0.05), 当盐浓度增长到0.18和0.24 mol/L时, 随着培养时间的增加, 叶绿素a含量积累缓慢, 处于较低水平。结果表明: 低浓度的盐可以促进叶绿素a含量的增加, 当盐浓度超过平凡舟形藻耐受的临界值, 会抑制叶绿素a含量增加。
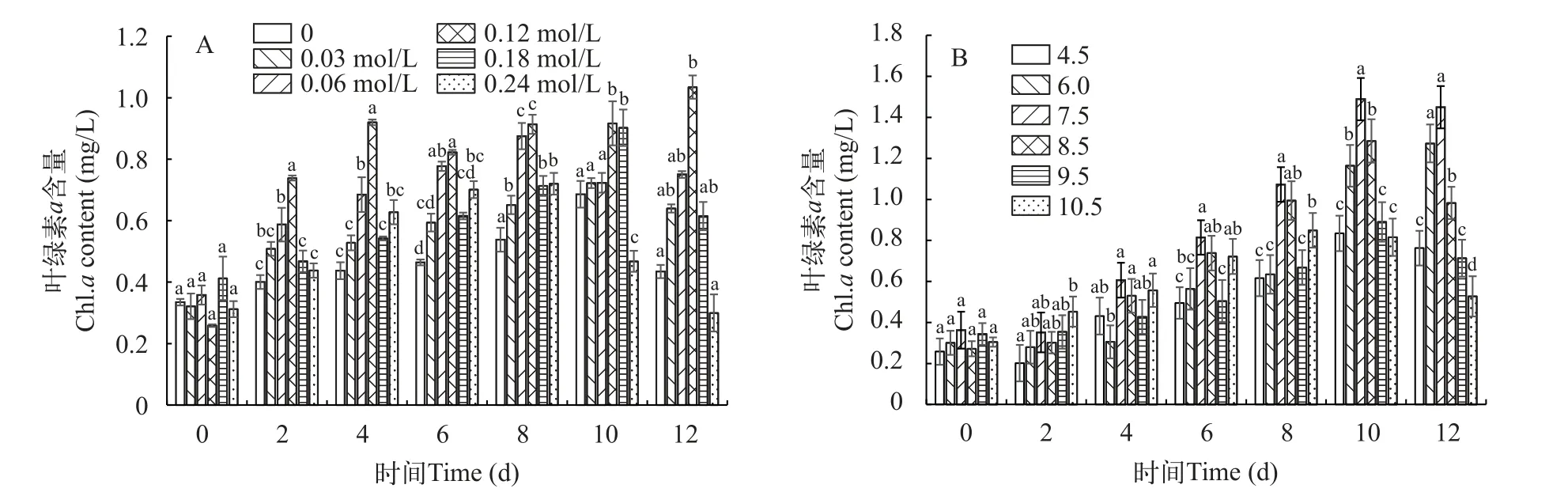
图2 盐度(A) 和pH (B)处理下平凡舟形藻叶绿素a含量Fig.2 Effect of salinity (A) and pH (B) on the chla content of Navicula trivialis
叶绿素a含量在不同pH处理条件下有显著性差异(图2B), 平凡舟形藻的叶绿素a含量最高值出现在pH为7.5时为1.48 mg/L。随着pH的增加或减少,叶绿素a的含量显著下降(P<0.05)。当pH为4.5、6.0、8.5、9.5和10.5时都抑制了叶绿素a的积累。当pH在碱性条件下, 如8.5、9.5和10.5, 叶绿素a含量相较于pH 7.5时显著下降(P<0.05), 分别为1.28、0.89和0.8 mg/L时, 相对于pH 7.5降低了1.15、1.66和1.83倍。当pH在酸性条件下, pH为4.5和6.0叶绿素a含量分别为0.83和1.27 mg/L, 呈下降趋势。所以pH为7.5更有利于平凡舟形藻叶绿素a含量的积累。
2.3 盐浓度和pH对平凡舟形藻叶绿素荧光参数的影响
叶绿素荧光参数可以很好的反映出微藻细胞光合系统的生理状态, 在研究不同环境条件下的光合效率具有高度敏感性和可靠性[27]。图3A和3B显示不同盐度处理水平下的叶绿素荧光参数PSⅡ的最大光能转化效率(Fv/Fm)和PSⅡ的潜在活性(Fv/Fo)的变化趋势, 二者趋势基本相同。在0—6d,各盐度处理组的叶绿素荧光参数Fv/Fm和Fv/Fo不同, 但整体呈现上升趋势。盐度为0—0.12 mol/L时, 平凡舟形藻的Fv/Fm和Fv/Fo显著高于盐浓度0.18和0.24 mol/L组(P<0.05), 且在盐度为0.12 mol/L时Fv/Fm和Fv/Fo的值最高。在培养8d后, 6组盐浓度处理下的叶绿素荧光参数开始呈下降趋势。结果表明, 高盐度和低盐度对平凡舟形藻的光合作用能力均有影响, 低浓度时平凡舟形藻的最大荧光比保持相对稳定。盐度为0.12 mol/L时, 是最适促进平凡舟形藻的光合效率的盐浓度。
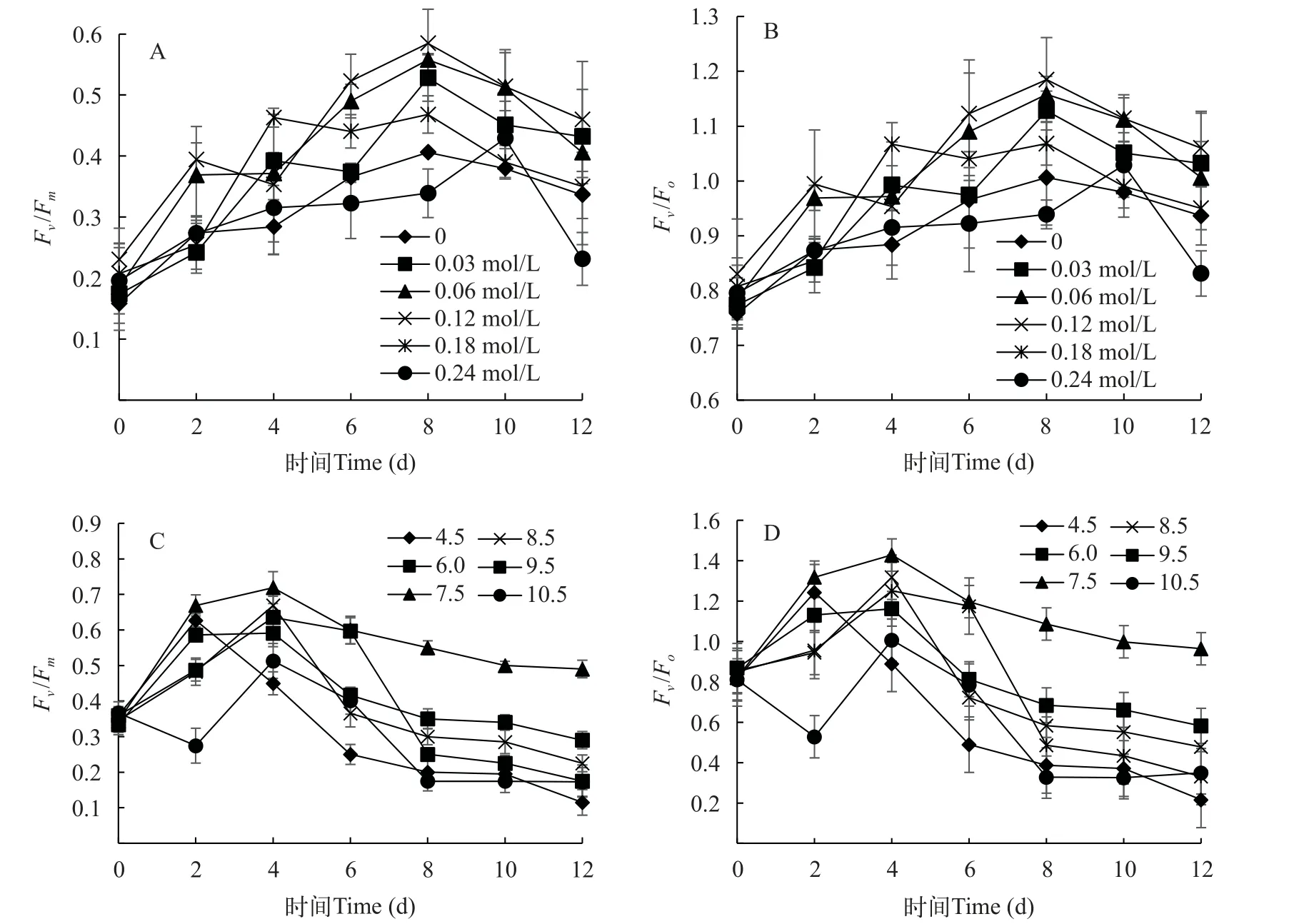
图3 不同盐浓度和pH下平凡舟形藻的叶绿素荧光参数Fv/Fm、Fv/FoFig.3 The effects of different salinities (A, B) and pH (C, D) on the Fv/Fm and Fv/Fo of chlorophyⅡ fluorescence parameters of Navicula trivialis
由图3C和3D可知, 平凡舟形藻叶绿素荧光参数Fv/Fm和Fv/Fo在pH为4.5—10.5均呈现相似的趋势, 都是先升高后降低, 在0—4d内,Fv/Fm和Fv/Fo叶绿素荧光参数数值迅速升高, 在pH为7.5时Fv/Fm和Fv/Fo叶绿素荧光参数值与其他处理水平有显著差异(P<0.05), 随着时间的增加, 在4d之后, 各处理组均呈现下降趋势, pH为7.5的Fv/Fm和Fv/Fo值高于其余pH。尤其是pH为4.5和10.5时,Fv/Fm和Fv/Fo参数值呈现明显的下降趋势, 这可能是由于光合作用对pH的变化敏感, 一旦超过藻类最适pH, 光合速率将会下降。
2.4 盐浓度和pH对平凡舟形藻总脂含量的影响
盐度胁迫在诱导藻类脂肪积累过程中起重要作用, 图4A显示了不同盐浓度胁迫下平凡舟形藻的总脂含量, 可以看出随着盐浓度的增加, 当氯化钠浓度从0 增加到0.24 mol/L时, 含油量总体呈上升趋势。对照组总脂含量为13.8%, 0.03和0.06 mol/L的总脂含量增加, 分别为17.2%和23.4%。盐浓度为0.12 mol/L的总脂含量与0.06 mol/L相比油脂含量增加, 当盐浓度增加到0.18和0.24 mol/L时, 总脂含量持续增加, 分别为30.3%和34.9%, 与其他盐处理组有显著性差异(P<0.05), 盐浓度0.24 mol/L的总脂含量比盐浓度0.18 mol/L高了4.7%。
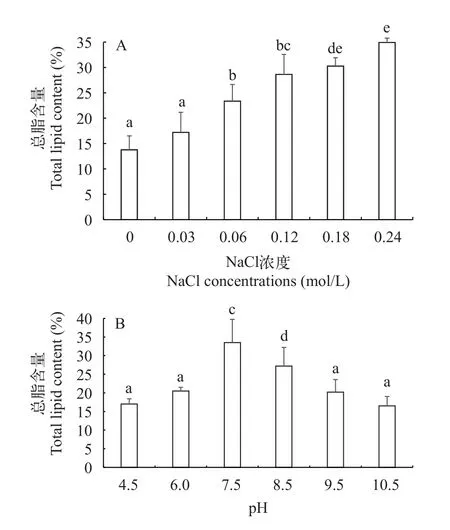
图4 不同盐浓度和pH下平凡舟形藻的总脂含量Fig.4 Effect of of salinity (A) and pH (B) total lipid content of Navicula trivialis
图4B显示了不同pH胁迫下平凡舟形藻的总脂含量, 可以看出不同的pH对总脂含量的影响不同。以pH为7.5的总脂含量最高, 为33.5%, 与其他pH处理组有显著性差异(P<0.05)。pH 8.5次之, 为27.2%。酸性和碱性pH并不利于总脂含量的积累, 如4.5、9.5和10.5, 总脂含量分别为17.0%、20.2%和16.5%,三者pH的总脂含量差异不显著(P>0.05)。pH为6.0时, 总脂含量达到20.5%, 由于在pH为7.5条件下的平凡舟形藻细胞密度和总脂含量最高, 因此确定了培养平凡舟形藻的最佳pH为7.5。
3 讨论
盐度和pH是影响微藻脂质合成的重要因素, 盐度的升高和pH的改变有利于诱导饱和脂肪酸的合成[28]。由于盐胁迫是大规模的, 如果能够发现一种在盐胁迫下可以积累脂肪的藻株, 这将为生物柴油的生产提供新的途径。
3.1 盐度和pH对平凡舟形藻生长的影响
盐浓度的不同会改变微藻的生长速度和组成[29]。当微藻细胞暴露在盐胁迫下时, 会出现特殊的适应机制, 例如细胞从活跃的分裂转变为储存能量, 积累渗透保护溶质以及保护生物化合物, 通过调节细胞膜的离子交换过程, 恢复生理生化过程[30,31]。pH决定了二氧化碳和必要营养物质的溶解度和可用性, 可以对藻类的新陈代谢产生重大影响[32]。较高的pH限制了二氧化碳中碳的可获得性, 而二氧化碳反过来又抑制了藻类的生长[32,33]。当pH较高时, 藻类的碳以碳酸盐的形式存在。较高的pH也会降低藻类对游离二氧化碳的亲和力[33,34]。与碱性pH类似, 酸性条件会改变营养物质的吸收[35]或引起金属毒性[36], 从而影响藻类的生长, 大多数藻类在中性pH为7.0—7.6附近能有最大限度的生长。改变盐度和pH相对来说操作简便, 有利于微藻生产生物柴油的规模化, 以增加更多的脂肪产量, 从而降低生物柴油的生产成本。
本研究表明: 在盐浓度0上升到0.24 mol/L这个区间时, 盐浓度为0—0.12 mol/L, 对平凡舟形藻细胞密度和叶绿素a含量起促进作用, 当盐浓度为0.12 mol/L时, 平凡舟形藻的细胞密度和叶绿素a含量达到最大值。Salama等[37]报道称, 在低盐浓度对微藻生长速度有促进作用。平凡舟形藻随着盐浓度的逐渐增加到0.24 mol/L时, 细胞密度和叶绿素a含量逐渐降低。据Markou等[38]报道随着盐浓度的增加, 小球藻YH703的干重和细胞密度显著降低。细胞密度和叶绿素a随着时间的推移而降低后, 可能是因为在营养耗竭后达到稳定阶段, 微藻细胞密度下降, 生长速度下降, 证明高盐度会抑制微藻的生长。这或许是由于低浓度钠离子提高了细胞离子交换能力, 加快了藻类新陈代谢, 从而促进了藻细胞的生长, 而在高盐度条件下, 藻细胞需要更多的能力来维持正常的渗透压和离子浓度, 用于生长的能量减少[39]。叶绿素荧光参数Fv/Fm和Fv/Fo在盐浓度为0—0.12 mol/L时, 平凡舟形藻的光合效率高,随着盐浓度的上升, 光合效率大幅度降低, Feng等[40]发现盐度过高或者过低都会影响梅尼小环藻的光合能力和PSⅡ活性。这表明在适宜的盐度下微藻的叶绿素荧光参数呈现增加趋势。藻细胞生长较慢, 因为盐度过低影响藻细胞渗透压, 影响藻细胞吸收营养盐的能力, 因此也影响微藻的生长。产生这种现象的原因可能是Na+在渗透调节和细胞膜电位调节中起重要作用, 然而, 过量的NaCl导致与光系统Ⅱ结合的多肽解离, 从而降低了光合机制的电子传递[41]。
3.2 盐度和pH对平凡舟形藻叶绿素a含量的影响
一般认为, 不同藻类的最适pH不同, 极端的pH条件影响藻类的光合作用、生长和养分同化[42]。在本研究中, 在酸性条件下(pH 4.5和6.0)和碱性条件下(pH 9.5和 10.5), 这4个pH使平凡舟形藻的细胞密度、叶绿素a含量显著减少, 对叶绿素荧光参数Fv/Fm和Fv/Fo也产生了抑制, 反映了pH过高或者过低对反应中心PSⅡ具有破坏作用, 使之受到限制[43]。在pH为7.5时, 细胞密度和叶绿素a含量达到最大值,叶绿素荧光参数Fv/Fm和Fv/Fo高, 其次是pH为8.5。王铭等[44]报道, 在pH为7.5时微藻的生理状态与光合作用表现为最佳。由此可见, 过酸或者过碱都不利于平凡舟形藻的生长, pH为7.5是最适宜平凡舟形藻生存的环境。Sakarika等[45]的研究表明在碱性值7.5和8.0时, 微藻的生长超过对照组20%—25%,表明在这个狭窄的pH范围内生长条件是最优的。微藻培养过程中过酸或者过碱会对平凡舟形藻的细胞产生一定的伤害, 使其破坏或死亡, 影响其生长。
3.3 盐度和pH对平凡舟形藻油脂含量的影响
盐度胁迫影响着与微藻生长发育相关的各种生理生化机制, 它也可以引起微藻脂肪含量的增加,它在引起脂肪酸代谢变化方面也发挥了重要作用[46]。在本研究中, 在0—0.24 mol/L随着盐浓度的逐渐升高, 总脂含量渐渐增加由13.7%上升到34.9%。对于脂肪含量, Hu[47]发现盐度的上升伴随着渗透压的增加, 藻类的总脂含量略有增加。在不同盐浓度胁迫下, 微藻的生长条件会发生改变, 从而对总脂含量产生影响。本研究表明, 高盐度条件(盐度高于0.12 mol/L)会抑制藻类的生长, 但高盐度有利于藻类总脂的积累。当氯化钠浓度从0.06增加到0.4 mol/L时, 普通小球藻的含油量可以提高到49%[48], 比本实验中的总脂含量高。有研究表明梅尼小环藻在盐度为25 g/L脂质含量达到最高, 达21%[49], 比本实验研究中的低。所以, 最适盐度和叶绿素含量趋势的差异可能因藻种而异。因此, 应选择合适的盐度来提高脂肪产量。这说明盐浓度的增加可以促进平凡舟形藻的油脂积累。有可能是因为在高浓度盐调控下微藻的生长需要更多能量, 使藻细胞内储存的油脂含量相应增加[50]。也有研究表明, 细胞脂质的增加可能是对渗透胁迫引起的微藻细胞损伤而形成的一种防御机制[51]。盐胁迫有效地促进了微藻细胞脂质的积累。到目前为止, 已有研究证明盐胁迫可以有效促进微藻细胞中脂质的积累[3,52,53], 但与此同时, 通常也会显著抑制微藻的生长, 导致最终微藻的脂质生产力不能得到很大的提高[54,55]。为了克服生物量产量和脂质生产之间的矛盾, 需要一种允许最佳生长速度和允许脂质增强的培养模式。有报道称在微藻大规模培养中, 首先提供最适合微藻生长的最佳条件, 随后需要具有应激条件的介质来诱导脂质生物合成[56], 以确保微藻的生长和收获更多油脂。
pH的不同也引起了平凡舟形藻总脂含量的变化。pH为7.5时总脂含量最高, 其余pH处理水平下的总脂含量低。对于pH胁迫, 据报道在最适pH条件下培养微藻可以提高生物量和脂肪含量, 在pH为7.5时, 可以显著提高微拟绿球藻的生物量和总脂含量[57]。但也有研究表示微藻在碱性的环境下利于油脂积累[58], 与本研究结果不同。所以不同的藻种对pH适应性不同, 应选用合适的pH。在pH为7.5时是最有利于平凡舟形藻的生长, 也可以提高其总脂含量。Sakarika等[59]报道称微藻细胞在适应了新的pH条件后, 代谢被激活, 除了繁殖外, 微藻还消耗提供的碳源(葡萄糖), 以积累脂质。
淡水藻株和海水藻株在不同环境因素下诱导油脂含量不同, 本实验中所使用的藻种为淡水藻株,不同淡水藻株在盐度和pH胁迫下的生长状况和总脂含量也不相同。根据藻类及其自然栖息地的不同, 诱导脂质积累的最佳氯化钠浓度水平也不同。淡水藻株Desmodesmus abundans在添加氯化钠的情况下, 可获得较高的油脂产率[56]。Chlorella sorokinianaHS1是一株淡水小球藻, 但其在海水盐度下也能生长, 并积累脂质, 在淡水中培养, 随后在咸水中积累脂肪。这可以缓解大规模养殖中的应激诱导等生产限制, 特别是营养限制, 以实现更高的脂肪生产率[60]。丰富栅藻在pH为6和8时是最适合生长和油脂积累[58]。淡水微藻在环境胁迫下特别是在海水盐度或者高于海水盐度的条件下, 会导致微藻油脂的积累。使微藻在淡水或废水中培养, 导致微藻生物量的增加, 然后在盐水中培养以积累脂质。这可以缓解生产限制, 以实现更高的脂质生产力。此外, 也有一些关于盐度和pH对各种海洋微藻的生长和脂肪积累影响的报道。梁英等[61]研究表明盐度对6种海洋硅藻生长与脂肪酸均有不同影响,证明盐浓度对于不同藻种作用不同。有报道研究了在同一盐度下5株海洋硅藻的总脂含量, 从而筛选出3种具有开发潜能的海洋硅藻[62]。江灵芝等[63]以蛋白核小球藻820为实验材料, 研究了3种盐度对其生长和油脂含量的影响, 以了解该小球藻对盐度的适应能力。研究结果表明, 蛋白核小球藻820的生长随盐度增加而变慢, 而油脂含量随盐度增加而升高。在300 mmol/L氯化钠培养处理的原壳小球藻虽然生物量只有对照的73.55%, 但是总脂含量却是对照的2.22倍, 可见一定浓度的氯化钠处理可显著提高原壳小球藻的油脂含量[64]。当培养液盐度上升至2.0%时, 是不添加NaCl培养基中微藻的油脂含量的1.6倍, 表明通过一定的盐度胁迫将微藻的油脂含量提升了60%左右。盐度对于提高微藻的油脂含量具有明显的促进作用[65]。Mus等[66]在研究pH对三角褐指藻的实验中发现在pH从8.0变化为7.5时, 脂肪含量增加。目前, 利用海洋微藻生产油脂的研究较多, 对淡水微藻相关研究相对较少, 本研究可以为淡水硅藻产油方面提供基础资料。
随着水体盐碱化程度的日益严重和淡水资源的日益匮乏, 藻类被视为发展生产可持续能源的最佳原材料, 盐胁迫是最常见的一种环境刺激, 在微藻细胞中, 盐胁迫可以影响脂类的合成, 微藻在一定程度上都可以耐受一定的盐浓度, 但是过量的盐胁迫也会影响光合作用进而影响生物量和脂质合成量。氯化钠是非常廉价的盐类, 同时也是营养的组成部分, 通过调整培养基中氯化钠的浓度达到不影响生物量同时增加脂含量, 可以大大降低生产成本。添加氯化钠和找最适pH是一种经济适用的增脂策略, 可考虑应用于微藻生物柴油的生产。
4 结论
微藻的生长受到光照强度、营养元素、温度和pH等培养条件的影响, 进而影响微藻的油脂产量, 为探究平凡舟形藻在不同盐度和pH培养条件下生长和油脂含量的影响, 进行为期12d的胁迫培养。结果表明, 盐浓度较低时可以促进平凡舟形藻的生产能力, 随盐浓度升高, 平凡舟形藻细胞生长受到抑制, 盐浓度为0.12 mol/L是适合平凡舟形藻生长的盐浓度。但总脂含量则相反, 随着盐浓度的不断增加至0.24 mol/L, 总脂含量达到34.9%。在pH 7.5的培养条件下, 平凡舟形藻表现出较高的细胞密度增长, 油脂含量升高。藻类可以在应急条件下产生大量的脂质, 这些脂质可以很容易地转化为可替代的燃料, 是一种可持续生物燃料来源。本研究结果表明, 适当改变培养基中盐的浓度和pH, 可以调控平凡舟形藻的生长速度和油脂含量, 本研究为微藻生产生物柴油提供参考数据。微藻作为可持续的生物能源原料, 具有巨大的发展潜力。在营养物质耗尽的情况下, 淀粉合成会转向油脂的合成, 因此胁迫条件将是提高微藻油脂含量的重要手段之一。如何将培养条件与胁迫方式有机结合, 从而使微藻细胞既能达到生物量最大, 也能保证油脂的积累量最多, 还需进一步进行深入研究。
猜你喜欢
草业学报(2022年5期)2022-07-08 06:24:40
古今农业(2022年1期)2022-05-05 06:58:42
粮食与食品工业(2021年3期)2021-06-18 07:50:48
草业学报(2021年3期)2021-03-22 02:22:00
水生生物学报(2019年3期)2019-05-29 06:58:48
草业学报(2018年10期)2018-10-19 05:25:44
电镀与环保(2017年5期)2017-12-19 12:06:03
环境保护与循环经济(2017年3期)2017-09-26 11:42:30
中国油脂(2017年7期)2017-09-16 04:28:48
水生生物学报(2014年2期)2014-05-27 08:07:30
